
诺顿人际神经生物学系列
Allan N. Schore, PhD, 系列主编
Daniel J. Siegel, MD, 创始主编
心理健康领域正处于一个极其激动人心的增长和概念重组时期。来自各种科学研究的独立发现正在汇聚成关于心智和心理健康的跨学科视角。人类发展的人际神经生物学使我们能够理解,心智和大脑的结构与功能是由经验塑造的,尤其是涉及情感关系的经验。
诺顿人际神经生物学系列将提供前沿的跨学科视角,进一步增进我们对人类心智复杂神经生物学的理解。通过借鉴传统上相互独立的广泛研究领域——如神经生物学、遗传学、记忆、依恋、复杂系统、人类学和进化心理学——这些著作将为心理健康专业人员提供对临床医生来说往往难以接触的科学发现的回顾和综合。这些书籍旨在通过将众多研究领域的发现转化为共同的语言和概念框架,找到知识的统一性或融贯性(consilience),来推进我们对人类经验的理解。该系列将把现代科学的精华与心理治疗的治愈艺术融为一体。
诺顿专业书籍
Jaak Panksepp Lucy Biven
前言 Daniel J. Siegel

献给 Tiina Alexandra Panksepp (1975–1991)
前言与致谢
前言 Daniel J. Siegel
第一章 祖先的激情
第二章 情感意识的进化:研究其他动物的情绪感受
第三章 寻求系统(SEEKING System):热切期待、渴望、欣快感的大脑来源,以及对一切事物的追求
第四章 愤怒的祖先来源(RAGE)
第五章 恐惧的祖先根源(FEAR)
第六章 超越本能:学习与记忆的情感基础
第七章 心智的欲望激情(LUST):从生殖冲动到浪漫爱情
第八章 养育之爱:关怀系统(CARE System)
第九章 生而哭泣:恐慌/悲伤系统(PANIC/GRIEF System)与维持生命的社会纽带的起源
第十章 大脑中如梦般的玩耍回路(PLAY):社交快乐与笑声的祖先来源
第十一章 迈向灵魂的神经生物学:核心自我(SELF)与初级过程感受的起源
第十二章 大脑情绪系统与心理生活的情感品质:从动物情感到人类心理治疗
第十三章 哲学反思与质疑:我们能否从小鼠到人类再回到小鼠?
参考文献
索引
我们所有人有时都会生气,尤其是当我们的利益被忽视或受挫时。传统脑科学告诉我们这种情绪是如何产生的吗?还没有。我们所有人有时都会感到孤独和悲伤。现代神经科学是否试图阐明我们本性的这些方面?我们才刚刚开始谈论这些事情,尽管在某些领域已经取得了巨大进展。我们大多数人从与他人的玩耍性互动中获得极大的快乐;有些人则不会,尤其是当他们抑郁时。神经科学对快乐的本质基本保持沉默,而心理学在研究和讨论其认知衍生物——幸福方面经历了一场革命,但对快乐的神经本质却鲜有洞见。
就像我们心智的许多其他情感力量一样,所有这些都来自大脑的功能,传统神经科学对于我们称之为情感(affects)的强烈情绪感受如何能从大脑活动中产生,却相对没有太多可说的。这是因为感受是主观体验的,有人说科学的传统第三人称测量方法(即对现象的外部观察)无法有效处理第一人称体验。我们不同意这种观点,因为其他哺乳动物拥有进化相关的大脑系统。现代神经科学完全有能力最终阐明哺乳动物大脑以非言语感受状态的形式对世界事件产生情感评价的方式——或者如一些文艺复兴时期学者所描述的,心智的激情。
这本书描述了一门名为情感神经科学(affective neuroscience)的新科学学科,它致力于阐明我们最强烈的情绪感受——原始情感(primal emotional affects)——如何从位于新皮层”思考帽”下方大脑区域的古老神经网络中产生。新皮层是一个产生复杂认知能力和文化的器官,对于复杂的知觉、学习和认知具有决定性的重要性。新皮层负责人类能够实现的几乎所有文化里程碑。神经科学也提供了一个重要信息——皮层内几乎所有的心理专门化功能都是习得的。目前还没有任何功能被实证证明是内在的、进化决定的”模块”。然而,如果没有大脑深处进化而来的基础心智,皮层将一事无成。新皮层下方那些古老的神经区域构成了我们的祖先心智——情感心智,它是进化专门化的,我们与许多其他动物共享这一心智。它是”考古宝藏”,因为它包含了我们一些最强烈感受的源头。那些古老的皮层下大脑系统是珍贵的、多彩的”宝石”,对于任何想要理解我们一生中曾经知道和将要体验的所有基本价值观根源的人来说都是如此。情感是构建生活美丑的基础。情感也会随经验而改变,但更多是量的改变而非质的改变。
这本书是对早期教科书《情感神经科学:人类和动物情绪的基础》(Panksepp, 1998a)的更新和通俗化尝试。这本教科书作为情绪心智科学的一种重要新方法获得了广泛关注,并已成为希望理解来访者基本情绪的临床医生的资料书。尽管对同类动物的研究对情感神经科学的发展至关重要,Jaak Panksepp开始他的工作时主要关注人类情绪,尤其是临床疾病中的情绪障碍。他很快意识到,如果没有适当的动物模型,就无法获得深入的神经科学理解。随着现代脑成像技术的出现,这一立场有所改变,但如果想真正理解大脑进化的功能网络,改变并不大。当人们静静地躺在无法容忍移动的脑扫描仪中时,很难产生强烈的情绪。尽管如此,通过那些壮观的人类脑成像技术获得的新证据已经阐明了许多关于情绪认知方面的内容,但关于大脑中这些感受的源头却知之甚少。初级过程情绪(primary-process emotions)都与运动相关,现在的证据表明,原始情绪感受来自控制我们本能情绪生活的同一古老大脑网络。尽管该领域有许多理论,事实表明这些原始情绪感受来自大脑的情绪行动网络。
总的来说,情绪这个话题引起了几乎所有人的极大兴趣——从必须处理已变得极端的人类感受的精神科医生,到任何对支配我们所做之事和我们在世界中身份的那些强大状态感到好奇的人。我们希望在这些章节中发现的内容将对许多人追求理解自己和他人(包括动物同伴)大有裨益,并认识到所有哺乳动物在情绪响应世界的方式上有多么相似。我们怀疑许多不同群体的人会发现这些观点特别有用。
我们发现哺乳动物大脑的古老皮层下区域至少包含七种基本情感系统:在这里,我们将这些系统称为寻求(SEEKING,期待)、恐惧(FEAR,焦虑)、愤怒(RAGE,怒火)、欲望(LUST,性兴奋)、照料(CARE,养育)、恐慌/悲伤(PANIC/GRIEF,悲痛)和玩耍(PLAY,社交快乐)。(我们稍后会解释为什么使用大写字母来标记这些系统;现在只需说明它们指定了我们大脑中进化上非常古老区域的特定功能网络。)
这本书应该引起精神科医生和其他心理健康专业人士以及情感、行为和认知神经科学学生的特别关注(它们各自对情绪的研究和讨论采取了相当不同的方法)。我们这里的重点将放在这些系统的初级过程(primary-process)性质上,但我们不会忽视大多数其他研究者正在研究的层次——次级过程(secondary process,内置的情绪学习机制)和三级过程(tertiary process,在人类体验中如此明显的情绪思考和审议)。
神经科学家未能从经验上处理情绪组织的初级加工(primary-process)层面(进化层面),这阻碍了情绪研究中不同方法之间尽可能连贯的综合。当人们沿着大脑和心智的进化层级上升时,设想情绪生活的方式会越来越多样化。相比之下,有大量证据表明,哺乳动物大脑的基本情感系统是哺乳动物心智的古老普遍价值结构,它们以个体情感体验类别的形式提供对世界的评价。在脑心智(BrainMind)复杂性中越往上——从初级到三级层面——整体方程就变得越可变和复杂。多种情绪流可能在思维心智中交叉,创造出大量心理学家经常关注的高级情绪——骄傲、羞耻、自信、内疚、嫉妒、信任、厌恶、支配等等,有数百种可能的变体。然而,如果没有对初级过程的清晰认识,对高级过程的重要研究仍然严重不完整。如果不能可靠地理解我们作为生存进化工具所继承的基本情感感受,我们就无法拥有可靠的心智理论。高级(社会建构的)感受很可能都需要我们进化能力的某些排列组合来感受某些方式。心理生活的所有方面都可能受到我们初级加工感受的影响,而低级脑心智(MindBrain)的整体情感谱系是高级心理健康问题的基础。心智的低级力量最终在多大程度上被新兴的高级功能所塑造,这将是未来工作中非常有趣的内容。我们已经知道,高级大脑过程既可以唤起情绪,也可以显著地减少情绪。所有这些在很长一段时间内仍将是情感神经科学最有趣的方面。
医生,特别是精神科医生,必须了解这些情感系统,因为它们为身心互动提供了新的见解。其中一些互动已经广为人知。例如,考虑持续焦虑的痛苦,这是恐惧(FEAR)系统的表现。恐惧系统的唤起最终会导致皮质醇的过度产生。在最佳条件下,当动物感到害怕时,皮质醇的分泌会动员葡萄糖作为骨骼肌的能量供应,以防动物决定逃跑。通过这种方式,皮质醇分泌是有益的。然而,如果升高持续时间过长,过度分泌可能会开始损害身体。通常当皮质醇通过血液循环回到大脑时,下丘脑的室旁核(PVN)会施加抑制作用,停止皮质醇的进一步释放。然而,如果一个人或动物受到过量的压力——当他们长期感到恐惧或焦虑时——室旁核可能无法停止皮质醇的产生。
尽管皮质醇的情绪效应的强度和时间模式在不同人之间可能有很大差异,但所有内脏器官和大脑的许多区域,以及免疫系统,都可能受到皮质醇长期过量的不利影响。大脑和身体中许多由此产生的压力诱导级联反应也可能导致这些不利影响。长期高皮质醇水平在许多精神综合征中很常见,尤其是在抑郁症中。目前尚不清楚皮质醇的过度分泌如何促进临床抑郁症。然而,各种生长因子(如脑源性神经营养因子BDNF)正常产生的破坏已被牵涉其中。玩耍往往通过这类化学物质促进积极情感(见第10章),为常识原则提供了证据,即积极和消极感受在心智的情感经济中相互抵消。
此外,当人们严重抑郁时,他们经常遭受海马体损伤,因为过量的皮质醇会导致海马细胞萎缩,有时甚至死亡。也许令一些人惊讶的是,简单地挠老鼠痒痒并激发老鼠”笑”可以促进海马体中新神经元的萌发(见第10章)。海马体是一个对创建陈述性(declarative)和情景性(episodic)记忆——知识和经验的意识记忆——至关重要的大脑结构(见第6章)。没有这个大脑区域,人将生活在永恒的当下,对已经过去的事件没有记忆。因此,过度的皮质醇释放可以参与许多严重的精神障碍,包括记忆缺陷。
同样,小剂量的阿片类物质会提升情绪并促进社会团结。大剂量则会引起中毒。事实上,适量的内源性阿片类物质可以产生医学上的有益效果。例如,安慰剂效应(placebo effect)——患者对假药物产生良好反应——可以用这种情绪化学来解释。如果患者感到自己的需求得到考虑和照顾,那么被照顾的积极感受会伴随着大脑中镇静性内源性阿片类物质的释放,从而减轻与悲伤/恐慌(GRIEF/PANIC)系统相关的感受。
除了产生良好的情绪感受外,阿片类物质还能减少应激性唤醒,减轻身体和心理疼痛的感觉,并产生各种免疫益处。因此,这些患者会感到安慰,在医疗上的状况也会比他们认为没有人关心时好得多。我们现在知道,安慰剂效应是真实的医学现象,主要通过激活大脑阿片类系统发挥作用。因此,这些治愈倾向可以被纳洛酮(naloxone)和纳曲酮(naltrexone)等阻断阿片类物质作用的药物减弱,甚至消除。
过去,当一个表面上健康的患者表现出情绪激动并抱怨身体症状时,医生往往认为这些症状是心身性的,“完全是心理作用”,因此不是身体上的或”真实的”。这不再是对心身疾病的公认观点。一旦我们认识到情感来自于由大脑化学物质驱动的情绪系统,这些化学物质也可以对大脑和身体的功能产生最终影响,那么情绪障碍和身体障碍之间的界限就会缩小到消失的程度。虽然心智和大脑看起来是不同的实体,心智是无形的,而大脑是物质的,但它们实际上是同一事物。心智大脑(MindBrain)或大脑心智(BrainMind)是一个统一的实体,与身体之间没有任何界限——它是整个物理系统不可分割的一部分。
理解大脑情绪系统以及它们可能产生的心理和身体症状,不仅对一般医学很重要,还为当代精神病学提供了一个全新的视角。情感神经科学为治疗情绪失衡的真实和特定症状指明了方向,即大脑心智的自然内表型(endophenotypes),而不是诸如自闭症、抑郁症和精神分裂症等模糊的疾病学抽象概念,这些概念是从神经科学之前的精神障碍分类传承下来的。这些诊断概念是从平均临床表现中推断出来的。但我们现在知道,所有这些都是高度模糊的——每个诊断类别都是一个概念性的总括,涵盖了大量重叠的心智大脑问题。
例如,大鼠天生害怕捕食者的气味。它们也天生害怕光线充足的开放空间,因此更喜欢待在黑暗和隐蔽的区域。当它们被放置在一个不熟悉的笼子里时,也经常表现出恐惧症状(通常通过僵住行为、血压升高和排便频率增加来测量)。常见的抗焦虑药物如苯二氮䓬类(benzodiazepines)可以平息对开放空间和新笼子的恐惧。然而,大鼠仍然害怕捕食者的气味,这表明这是一种有些不同的恐惧。令人惊讶的是,吗啡在减少分离痛苦方面非常有效,也能够减少大鼠对捕食者气味的恐惧反应。通常我们会将不同类型的恐惧归入同一类别,但情感大脑研究表明,存在针对不同类型恐惧和焦虑的神经模型。如果是这样,那么我们应该能够开发出特定的药物来治疗每种类型。正如我们将在后面章节中详细探讨的,与身体危险相关的恐惧(FEAR系统)和与分离焦虑相关的恐慌型恐惧(GRIEF/PANIC系统)之间存在令人信服的区别。
在相当长的一段时间里,精神病学医学的发展一直受到人为概念的阻碍,这些概念是从复杂的症状学而非大脑研究中收集的。如果精神病学研究更多地与心智大脑的实际情绪症状联系起来,并更有效地与功能神经科学联系起来,我们可能会取得更快的进步。例如,我们可能很容易开发出针对易怒和愤怒的特定药物。这目前很难实现,因为没有为过度愤怒设计官方诊断类别(也许除了间歇性暴发障碍)。然而,整个社会,特别是儿童,经常是过度愤怒(RAGE)的受害者。我们已经有了P物质受体拮抗剂和阿瑞匹坦(aprepitant,目前用于治疗恶心的药物)等药物,如果可以从动物数据推广的话,应该能够减少愤怒易怒(见第4章)。目前,人们对更好地理解这些情绪内表型非常感兴趣,以便我们的诊断工具可以得到根本性修订,并可以开发出更好的药物。
对七种基本情绪系统的了解已经开始彻底改变心理治疗的实践,因为它提供了目前可用的最全面、基于数据的初级过程情绪大脑分类法。对这些系统的了解也意味着对人类情绪如何运作有更全面的看法。我们帮助提供了一个基于数据的分类法来讨论情绪生活的基础,并提供了许多特定大脑功能在情感生活中重要性的例子——例如,内源性阿片类物质和催产素(oxytocin)在支持性社会关系的积极情感中的强大作用。这为健康情绪发展严重依赖于维持支持性人际互动的观点提供了神经生物学支持。在严峻的情况下,开具支持这种大脑化学物质的安全药物可以促进和巩固心理治疗实践。
为了突出我们在心理治疗中处理关键概念问题的方法,让我们来看看当前观点与经典精神分析思想的一些原则之间的对比。我们怀着对该领域理论精妙性的学术敬意来进行这一对比,但这里我们主要关注我们如何以不同于精神分析理论家的方式看待基本情感过程,后者的观点基于临床洞察而非神经科学研究。
尽管心理治疗在过去半个世纪中朝着许多不同方向发展,但许多治疗师仍然继续依赖精神分析理论来了解基本情感。此外,当前流行的情绪观点,即设想在某种程度上简单的正负情感效价的两极图式,通过高低唤醒来调节,实际上与精神分析的树并未相距太远。弗洛伊德认为人类驱力植根于我们的生理需求,他将这些驱力归为两类:力比多(libido)和攻击性(aggression)。驱力在心理上表现为愿望性思想——即充满情感色彩的思想。根据弗洛伊德理论,两种主要情感涉及对性欲望和攻击冲动的愿望。
弗洛伊德论证了几种驱力表达类型,每种都植根于力比多发展的不同阶段:口欲期、肛欲期、性器期和俄狄浦斯期。攻击驱力也类似地沿着这些发展阶段进行划分。这为两种相互作用的驱力及其结果性情感愿望提供了更广阔的范围。然而,离散情感的范围远比神经科学研究所揭示的七种情感系统所产生的要有限得多。我们很高兴地注意到,SEEKING系统与弗洛伊德的力比多驱力提供了一个有趣的平行(只要他将力比多视为一种通用的欲求力量,而不是狭义的性术语)。然而,很难将弗洛伊德关于焦虑的观点,以及他关于与依恋和情感纽带相关的性欲的观点,以及许多其他方面的观点,与我们从严格的神经科学调查中获得的知识相协调。
大多数现代精神分析和认知行为治疗方法未能明确识别SEEKING作为一种基本情感冲动。一些研究者还倾向于混淆FEAR和PANIC/GRIEF,将焦虑视为单一表现。在许多精神分析理论中,社会互动的重要性也没有得到充分强调。弗洛伊德学派将社会互动视为满足性和攻击冲动的衍生手段。社会需求不被视为基本冲动,而这种冲动有时可能在重要性上超越性或攻击,即使在基本本能冲动层面也是如此。尽管客体关系理论家强调人际需求的重要性,但他们倾向于关注家庭内的早期关系,特别是母子纽带。今天我们有了更多关于PLAY的重要性,例如,以及相关的社会支配的基本心理学的信息。
同时,我们在这里提供的内容对每个心理治疗师必须应对的人类心理生活的独特、个体化方面几乎没有涉及。存在着更高级的三级过程认知功能,情绪在现实生活中会与之相互作用。但通过澄清我们在试图帮助情绪困扰者时需要考虑的原始心理能量,它可能会简化心理治疗师的三级过程任务。如何做到?那需要另一本书。但也许现在一个洞察就足够了:下层大脑似乎以这样的方式组织,即在任何一个时刻只有一种原始情感状态占主导。这种”单一狂热”(monomania),由于缺乏更好的词,也会诱导认知装置”跟随”强迫性的自私沉思。治疗的目标是促进高级心理装置中更复杂的视角转换——亚里士多德所称的实践智慧(phronesis),通过理解”低级思维”方式成为自己激情的主人。
也许经典精神分析临床实践中的这个核心问题可以通过情感神经科学来解决。正如我们所看到的,经典精神分析可能不如其本可能达到的效果的一个关键原因在于解释(interpretation)——谈话治疗的核心——长期以来被认为是主要的心理治疗工具。精神分析师倾向于专注于情感状态与其相应认知表现(愿望)之间的关系。他们长期以来一直假设,通过解释相关的思想和观念,通过揭示它们在童年的起源并解释它们原始的情感意义,患者将被治愈。但我们如何知道这能解开大多数人生活中的情感”结”?
假设在童年时期,一个男孩在父亲手中遭受了身体和情感虐待。成年后,这个男人自己倾向于欺凌弱者。心理治疗师会帮助患者识别其成年人格中的问题区域,即他欺凌甚至虐待他人的倾向,然后将这些特征追溯到童年。治疗师可能会解释说,这个男人欺凌弱者和虐待脆弱者是为了以一种不会导致报复的方式发泄他对父亲的愤怒。其他解释可能会强调他欺凌他人是为了恢复他的男性自尊的可能性。由于这些以及其他解释,患者可能会被治愈或至少继续过上更幸福的生活。在这种观点中,认知问题被视为通往情感问题的门户。
精神分析传统之后,在行为主义时代,出现了高度聚焦的”行为矫正疗法”,其中认知和情感问题都被搁置一旁,治疗师试图通过调整强化条件来塑造不适应的行为模式。随着认知革命的到来,焦点转移到”认知行为疗法”(CBT),这种疗法对某些障碍如特定恐惧症非常有效(Beck, 1976)。现在,随着人们认识到情感潮流是精神障碍的核心,风向再次转变。
情感在脑心进化中的首要地位表明,疗法必须对人类情感生活有清晰的认识,以便治疗师能够为精神问题提供最佳的理解和帮助。事实上,这种自下而上的观点可能会颠覆心理学和哲学中认知”解释性”的情绪理论。显然,尽管认知问题在三级过程情绪中占据重要地位,但初级过程情绪必须按其自身的方式来处理。当传统治疗模式(精神分析或CBT)无法平息情绪风暴时,那么可能需要药物治疗。目前,大多数这类药物还不存在,因为精神科医生对情绪大脑的解剖结构和化学机制了解不够。我们希望本书能激发更多研究,从而促成此类药物的开发。从某种意义上说,需要的是对所有治疗传统的更全面整合,从动力-精神分析到新一代情感平衡疗法,这将是本书的主要焦点(见[第12章])。
例如,考虑上述讨论的案例,假设童年遭受的虐待已经致命地敏化了恐惧和愤怒系统,使得相应的情感难以或不可能平息。即使治疗师成功地让患者相信其问题的根源,即使患者清楚地意识到他对他人不公平、不公正,这也可能不足以产生任何治愈效果,因为他仍然会遭受压倒性的易怒,这可能表现为明显的欺凌欲望。
神经科学支持这一假设。两千年前,普鲁塔克指出:“愤怒的持续和频繁发作会在灵魂中产生易怒的倾向:这往往以暴躁、刻薄和乖戾告终,当心灵变得溃烂、易怒和好争吵,并被最轻微的事件所伤害时。”普鲁塔克似乎是对的。我们现在知道大脑的愤怒回路可以被敏化并变得过度反应。因此,即使患者完全理解其愤怒的根源,并做出极大的意志努力来抑制愤怒,他可能仍然无法停止感到长期的烦躁,并且会继续情绪不适。也许其他人可能免受其愤怒的有害影响,但患者本人可能会继续像治疗前一样痛苦,甚至可能更糟,因为他至少曾经有一个无法控制的情感宣泄口。
重点是思想并不总是比情感更强大,这就是为什么认知解释通常对严重精神病理学效果不佳。实际上,来访者可能会被治疗师”清楚”看到的复杂性所困惑。当情感占上风时,谈话治疗很可能会失败,因为解释方法这一基本心理治疗工具,在面对我们的原始激情时常常无效。也许这就是为什么连弗洛伊德本人都期待有一天能够直接对驱力施加化学影响,正如他所看到的那样。但这并不意味着心理治疗应该简单地被药物治疗所取代。情感神经科学研究强调,临床医生不应该把人类当作神经化学物质的袋子或”缸中之脑”来对待。情感感受是完整方程的一部分,当精神科医生寻求问题的新治疗方法时,不应忽视它们。此外,哺乳动物的大脑从根本上说是一个社会性大脑,必须这样对待。基本情绪系统不是在社会真空中运作的,即使在初级过程层面也是如此。因此,几乎所有心理-医学干预都需要适当的心理社会帮助来补充,不仅要追溯和解开(可能是终生的)基本情绪失衡的次级和三级过程衍生物,还要引导、促进和激活期望的初级过程情感。积极情感可以促进韧性(resilience),这对许多情绪问题都能产生持久的有益影响。情感神经科学强调,在所有未来的治疗学派思想中,社会情绪的作用必须保持关注焦点,以便最大化持久的改善。
所有希望充分了解人类情绪的人——从父母到教育工作者——都会想要理解感受是如何从大脑内部产生的。这些情感系统对大多数处理人类的学术学科都有重要意义,从哲学到经济学,从艺术到社会科学。
父母会想了解这些系统,以便评估孩子的正常发展。如果看到所有系统都达到了恰当的平衡,这表明孩子正在以情感健康的方式发展。但如果某个特定系统过度活跃或活跃不足,这可能表明存在问题。例如,一个过度用功或严肃的孩子可能PLAY系统活跃不足。PLAY系统让孩子学习社交行为规则——例如,何时合作、何时竞争,有时还要以幽默的方式退让并让别人获胜。当动物进行粗野打闹游戏时,如果一只动物获胜超过70%的次数,失败的动物就不再享受游戏,可能会完全退出这类互动。所以当孩子玩耍时,他们学习宝贵的社交技能,比如互惠的必要性和偶尔让步。孩子们会学习这些技能,因为如果不学,他们的玩伴可能会开始拒绝他们。
父母应该理解在孩子身上保持积极情感最佳平衡的重要性,尤其是在他们年幼时。皮层下情感系统可以通过经验变得敏感。神经科学家开始了解情感脑系统如何通过生活经历被塑造,通常是永久性的,就像承载我们身体动态进入世界的肌肉和骨骼随着时间发展和强化一样。这些变化可以延伸到基因被激活的水平,有时会导致终生的情感优势和弱点模式。理解这些表观遗传(环境诱导的)基因表达的长期变化,以及由此产生的大脑心智终生的优势和弱点,将是情感研究中最令人兴奋的未来篇章。
因此,如果孩子得到了大量的养育性关怀(CARE),形成了安全的社会纽带,并且通过PANIC/GRIEF系统的低活跃度促进了积极的依恋,那么孩子是幸运的。如果孩子有机会进行大量快乐的玩耍,如果孩子的好奇心得到了激发,那么支持这些能力的神经回路在一生中都会更加强健。另一方面,如果孩子遭受了引发其RAGE系统的不当挫折,或者如果孩子经历了高水平的FEAR或PANIC/GRIEF,那么她对这些负面感受的能力就会增强。然而,这并不意味着父母需要保护孩子免受负面情绪的影响。所有孩子都必须学会应对它们,因为它们是生活的自然组成部分。有理由相信,所有负面情绪,以小剂量可管理的形式,有助于长期心理韧性(resilience),这可能有助于抵御未来更持久的失望,从而避免抑郁。
教师肯定会从了解七个基本情感系统中受益。所有优秀的教师都会激发SEEKING系统,他们让学习成为一种令人兴奋的体验,而不仅仅是死记硬背。然而,鉴于许多学习涉及一定程度的枯燥乏味,教师也需要施加社会约束。尽责的孩子会得到表扬的奖励,从CARE和GRIEF/PANIC系统的积极社会纽带分支中产生满足感。然而,顽固的孩子往往必须忍受不认可的威胁,伴随着上述社会情感系统负面分支的激活,更不用说RAGE和FEAR的痛苦了。如果是这样,那个孩子的生活将被负面情感和担忧所支配,而不是可以激励孩子取得更大成就的积极情感。优雅地给予有过多负面情感的孩子第二次机会,可以是一种美妙的维持生命的体验。无论如何,良好管理的社会约束可以增强孩子容忍挫折的能力,并为他们应对成年生活中不可避免的挫折做好准备。
我们甚至会强调充足的身体游戏如何减少冲动和注意力缺陷多动障碍(ADHD)等问题的发生率。当孩子满足了他们身体游戏的自然冲动后,他们就能更好地准备在教室里安静坐着并集中注意力。重新引入游戏可能最有效的方式是把课间休息作为每天的第一堂课。实际上,这种需求过去在孩子步行上学并早到足以在课前遇到玩伴并一起游戏时就得到了满足。
某些情感类型似乎在特定角色和环境中工作得最好。每个管理者都需要赢得员工的信任和尊重。员工应该感到管理者会帮助他们解决工作中的问题,管理者应该相信员工会履行职责。这种隐含的社会契约建立在CARE系统的相互性之上。他们必须给予彼此所需要的东西,以感到安全并取得卓越成就。管理者也知道团队凝聚力的重要性。团队日可以通过培养PLAY精神来支持这一过程,让大型工作组的成员有机会在更亲密和放松的环境中互动。这种游戏式互动巩固了对劳动力团结重要的社会纽带。
与动物打交道的人会在这里发现许多关于控制动物行为的情绪的重要信息。事实上,美国最敏锐、最杰出的动物行为学家之一Temple Grandin——一位患有自闭症的杰出人士——在她引人入胜的著作《动物让我们成为人类》(2009)中提出了这类信息。这部作品也有助于证实长期以来人们所持有的信念:动物确实拥有情感感受。事实上,在学术领域之外,有一场迅速发展的运动正在认可和重视其他动物的情绪,但其中大部分是基于合理的信念和有趣的轶事,而非精心收集的科学事实。
我们书中总结的证据旨在提供一个基于实证而非观点的视角,来说明哺乳动物物种的情绪思维究竟是什么样的。目前基于证据的观点是,所有其他哺乳动物都充满情感激情——它们相当充满情感。正如我们将看到的,这一结论现在得到了大量实验证据的支持(Panksepp,1998a中有大量详细说明,在这里则较为简略)。那些仍然持否定态度的人坚守着由来已久的怀疑论。在这样做的过程中,他们通常未能将现代情感神经科学研究整合到他们的思维中。也许其他哺乳动物无法像我们那样思考它们的情感生活(它们的三级过程可能非常不同),但有力的证据表明,它们确实体验到全方位的一级过程情感。
我们可以继续列举那些能从理解情感神经科学中受益的人:哲学家、政治家、艺术家,以及其他想要创造更美好世界的文化领袖。但最重要的是,我们认为每个人在某种程度上都希望熟悉这些大自然母亲赋予我们大脑的基本生活工具。
我们都有很多要感谢的。
Jaak特别感谢他从妻子Anesa Miller那里获得的所有支持和建议,她阅读并编辑了整部手稿。她在接受淋巴瘤治疗期间完成了这项艰苦的工作。与此同时,Jaak也在与另一种淋巴瘤抗争(值得庆幸的是,他们目前都已完全缓解)。Jaak是华盛顿州立大学兽医学院VCAPP(兽医比较解剖学、药理学和生理学)系动物福利研究中心的成员。他感谢所有优秀的同事,是他们让科学追求再次成为一种乐趣。Jaak的实验室主管Sheri Six在这段时间里为维持他的实验室运转的诸多细节提供了宝贵的关注,在现代科学时代,这可能是一项艰巨的任务。她还用她对细节的敏锐眼光和致力于在研究中敏感使用动物的理念阅读了手稿。在过去的一年里,Mark Solms这位备受尊敬和爱戴的同事,也为每一章提供了有用而热情的意见。在这段漫长的出版之旅的最后阶段,Tim Lyons——一位前学生,他已经远不止是学生——在2010年夏末回来了几周,协助完成最后的润色,他消除了文本中许多残留的瑕疵。他的精力和奉献精神,特别是基于他在第二职业(在作为律师度过大部分职业生涯之后)临床/咨询方面的培训,大大改进了这本书。感谢所有在这一路上提供帮助的人。
Jaak感谢华盛顿州立大学相关科学系和爱达荷大学人文系的所有优秀同事,感谢他们在他第三段学术生涯的六年中提供的诚挚支持和友谊。1969年在马萨诸塞大学获得博士学位后,Jaak在苏塞克斯大学和伍斯特实验生物学基金会进行博士后工作。Jaak关于哺乳动物大脑一级过程情绪性的愿景,是在鲍灵格林州立大学(BGSU)从助理教授晋升为杰出心理生物学教授的30年工作中成熟起来的,这在其他地方可能无法实现。在因医疗问题和女儿Tiina的过早去世而提前退休后,Jaak加入了西北大学的Falk分子治疗中心,与Joe Moskal、Roger Kroes和Jeff Burgdorf的友谊以及智力和研究支持下,追求情感思维的遗传学研究。他继续与许多前同事合作,特别是在情感大脑的遗传学研究方面,希望确定控制哺乳动物情绪性的新神经化学途径。他感谢BGSU的许多同事,特别是Vern Bingman和Casey Cromwell,他们在2010年5月组织了一场庆祝他工作的学术研讨会,其中大部分内容作为《神经科学与生物行为评论》的特刊出现。
Jaak还感谢Audrey Gruss以及抑郁症研究希望基金会(HDRF)的朋友和同事,感谢他们对抑郁症问题的智力投入以及过去几年中卓有成效的互动。Jaak目前是HDRF的研究联合主任,他正在进行的研究主要致力于开发用于理解和治疗抑郁症的新动物模型。他被认为是该领域的革命者(有些人认为是激进分子),获得了许多奖项和认可。他的工作总结在400多篇科学出版物中,其中一半列在生物档案中,另一半则服务于社会科学。
Lucy Biven 曾担任英格兰莱斯特郡国家医疗服务体系(National Health Service)儿童及青少年心理健康服务部门的心理治疗主任。大约20年前,当她被密歇根州最高法院任命设计并实施一项监护权转移协议时,她开始对神经科学产生兴趣。该协议涉及将一名2岁半女孩的监护权从她视为父母的夫妇家中转移到她的亲生父母家中。和大多数同事一样,Lucy担心这个小女孩的心理发展,然而这个孩子进展良好,如今已是一位情感健康的年轻女性。这一切是如何成功的?只有神经科学提供了答案。
由此开始了对神经科学的持久兴趣。然而,即使在广泛阅读了多年之后,她仍感到不满意,因为大多数研究集中在感知、学习和记忆上,而不是情绪。当神经科学确实涉及情绪时,通常是恐惧及其在条件学习中的作用。神经科学并未聚焦于完整的情绪范围或情绪本身。
然后在2000年,她参加了由Mark Solms主持的国际神经精神分析学会(The International Neuropsychoanalysis Society)在伦敦举办的研讨会。Jaak Panksepp是主题演讲者。Jaak是第一位也是唯一一位直接聚焦于情绪脑的神经科学家。随后,Jaak和Lucy之间进行了一系列漫长而富有启发性的电子邮件交流,最终促成了本书的出版。
Jaak富有深度的研究提升了她的临床工作,但她还感激其他给予她指导和建议的人。首先是她的父亲Charles Brenner,一位精神分析学家,他清晰的思维和通俗易懂的书面表达始终提供了一个典范目标。Anna Freud在指导Lucy接受培训的伦敦汉普斯特德诊所(Hampstead Clinical)时仍然充满智慧活力,直到今天,她仍未遇到过更有天赋的临床医生。还是学生时,Lucy遇到了Vann Spruiell,他的临床和情感诚实让她看到精神分析既可以也应该是一项充满活力的追求以及智力努力。一路走来,还有其他优秀且有影响力的同事,其中包括Josephine Klein、Anne Alvarez和Thelma Hillaby。
Lucy曾在密歇根大学担任高级研究助理,在另一位杰出临床医生Humberto Nagera博士的卓越指导下工作。她是密歇根精神分析研究所(Michigan Psychoanalytic Institute)的教员,1985年,她因一篇临床论文获得了Ira Miller纪念奖。她曾担任《国际精神分析杂志》(International Journal of Psychoanalysis)和《精神分析季刊》(Psychoanalytic Quarterly)的编辑审稿人。
她撰写了几篇关于神经科学及其与心理治疗和精神病学相关性的论文,并在美国、英国、南非和墨西哥广泛演讲。最后,她职业生涯和个人生活中最重要的人是她的丈夫Barrie,她全心全意地感谢他。
我们都感谢W. W. Norton出版社的优秀员工将这项工作带到完成,特别是我们的责任编辑Deborah Malmud,她在本书写作过程中提供了指导和鼓励。
DANIEL J. SIEGEL, 医学博士
对我们内在主观生活和与他人相互联系的理解,在《心灵的考古学》(The Archaeology of Mind)的深入探索中得到了深刻而有益的阐明。通过探索我们的神经架构、社会关系以及我们的精神世界及其如何交织,神经科学家Jaak Panksepp和心理治疗师Lucy Biven创造了对人类生活古老起源的详细视角。这一重要综合的核心是这样一个观念:我们的皮层下回路(subcortical circuits)是”初级”体验的基础基质——情绪和动机塑造我们的主观生活,影响我们的行为,并塑造我们的关系。Panksepp和Biven提出,较高的新皮层区域在我们学习产生情绪反应方面发挥重要但明显”次要”的作用,而更深层的皮层下区域——仍然存在于我们较古老的哺乳动物和爬行动物回路中——塑造了我们日常精神体验的固有质地。
Jaak Panksepp的学术生涯致力于探索这些回路的本质,他的观点是这项工作的核心要素。在倡导非人类动物拥有需要被尊重和理解的内在情感世界的职业生涯之后,这位情感神经科学(affective neuroscience)领域的重要领导者现在将焦点转向利用这些对古老回路的新见解来帮助人类。Panksepp是一位直言不讳的倡导者,呼吁对动物王国的所有成员进行富有同情心的理解。通过他的工作,我们开始看到尊重主观生活内核的重要性,并将这些知识应用于帮助所有生命。
无论您是临床医生、教育工作者、研究人员,还是感兴趣的普通读者,您都会在这些页面中发现有用且详细的信息,内容涵盖了构成我们情感并塑造我们动机的七大主要原始回路的精彩讨论:寻求(SEEKING)、愤怒(RAGE)、恐惧(FEAR)、性欲(LUST)、关爱(CARE)、恐慌/悲伤(PANIC/GRIEF)和玩耍(PLAY)。虽然这些皮层下系统与更高级的新皮层的相互作用在我们作为人类的体验中自然是至关重要的,但在本书中,我们有机会深入探讨这些更古老的情感核心来源。我们知道,心理治疗和心理训练的许多方面都是新皮层随时间学习并能够改变我们情绪大脑各个方面的重要方式(参见Davidson & Begley, 2012年的有益讨论)。例如,正念冥想(mindfulness meditation)已被证明可以改变调节情绪、注意力、共情和自我理解的重要区域的皮层连接。依恋关系(attachment relationships)(参见Schore, 2012; Cozolino, 2010)也可能塑造前额叶皮层区域,这些区域连接我们广泛分离的高级和低级神经区域(参见Siegel, 2012a, 2012b)。因此,新皮层从经验中学习。
自然地,治疗师、教师、父母或其他对学习如何塑造我们的思维和大脑感兴趣的人会将这种神经可塑性(neuroplasticity)视为我们如何在整个生命周期中变化的重要维度(参见Doidge, 2007年关于皮层神经可塑性的概述)。那么,我们为什么要花时间去了解那些在我们出生之前——在子宫外学习开始之前——可能就已经形成良好的更”基本”或”原始”的神经区域呢?答案非常简单:这些皮层下的区域既是皮层以分化方式生长的基础(参见Trevarthen, 1990; McGilchrist, 2009),也是我们体验心理生活——我们核心的、内在的、时时刻刻生活的主观质感——的基础。此外,对这些深层结构的科学观点只会扩展我们的自我理解,并可以提供可能改善我们生活的赋能洞察。
在本书中,您会发现对抑郁、焦虑、悲伤和恐惧的深入讨论,这可能会阐明您个人生活的某些方面。还有一些有益的探讨,关于经验如何塑造记忆和情绪的回路,形成我们内心生活的神经基础,并改变我们调节情感反应的能力。这些讨论为临床医生提供了重要的视角,帮助他们了解客户/患者体验的本质,以及如何利用这些新知识来提高他们的共情理解和临床干预能力。人们在社交困难(如自闭症)、学习问题(如注意力缺陷状况)和情绪调节问题(如情绪障碍)方面所经历的挑战,都会因这部作品所揭示的视角而呈现出新的面貌。本书还为教师提供了一个独特的机会,让他们了解教育体验核心的动机、情绪和学习的深层回路。当我们意识到师生关系基于信任时,我们就会发现这些皮层下回路为有效的学习关系奠定了基础。如果您是学术研究人员,本书提供了关于情感神经科学(affective neuroscience)皮层下方面的广泛而详细的综述,以一个流畅的叙述呈现,这可能会激发一些理解该领域的新想法,并可能直接为您自己的项目提供信息。
作为一名受过研究人员和临床医生双重训练的人,我发现这本书对科学中一个经常被忽视的领域及其在治疗理解中的应用进行了引人入胜的探索。作为一名教育工作者和诺顿人际神经生物学系列丛书(Norton Series on Interpersonal Neurobiology)的创始编辑,我认为了解这些内容可以帮助我们将更有效的治疗方法和教育洞察带入我们的工作和世界。
如果可以的话,让我在这里提供一个建议,这可能有助于你吸收接下来的内容。如果你是科学家,你可能会对文中提供的大量细节和丰富的学术参考文献非常感兴趣。然而,如果你是临床医生、教育工作者或普通读者,你可能会发现采用不同的阅读方式会让这本书更有趣。这里有很多材料——以通俗易懂且引人入胜的方式呈现——关于神经回路、神经递质以及揭示我们对它们认知的研究,详细讨论并不少见。我给你的建议是:像读一本引人入胜的非虚构故事一样阅读这本书。就像你不会去记忆一本小说一样,不要担心记住所有关于研究的细节。你不会被测试记忆了多少内容!当你以这种更轻松的方式阅读时,你可能会发现你的大脑会自然地识别出随时间自然浮现的信息模式。最初不熟悉的术语可能会开始变得熟悉,不寻常的名称看起来和说起来会更舒适,这样你在继续阅读时会对这些不太常见的术语感到更自在。那些在流行媒体中常见的皮层下区域——如杏仁核(amygdala)和海马体(hippocampus)——都在这里出现。但你也会遇到不太知名的皮层下神经区域,如导水管周围灰质(PAG)和伏隔核(nucleus accumbens),它们在我们情感生活的这一考古叙事中也扮演着重要角色。你可能对多巴胺(dopamine)和血清素(serotonin)非常熟悉,但你也会在这里找到关于催乳素(prolactin)和催产素(oxytocin)的详细讨论。放松,只需倾听这个引人入胜的故事展开。放下那些古老的恐惧(FEAR)和恐慌(PANIC)反应(来自童年和学校),如果你试图记住所有阅读的内容可能会产生这些反应。相反,保持玩耍(PLAYFUL)的心态,寻找(SEEK)那些在阅读过程中对你感觉相关的内容。你即将体验贾克·潘克塞普(Jaak Panksepp)充满激情的思想以及他思考我们神经起源的方式。享受与贾克一起的旅程,让自己领略一生的热爱和学习成果!
心智考古学
……某些我们认为能表达某些精神状态的行为,是神经系统构造的直接结果,从一开始就独立于意志,并在很大程度上独立于习惯……我们目前的主题非常模糊,但由于其重要性,必须详细讨论;清楚地认识到我们的无知总是明智的。
——查尔斯·达尔文(Charles Darwin)(1872)
本书带我们进行一次考古挖掘,深入哺乳动物大脑的深处,探寻我们情感心智的祖先来源。据我们所知,所有哺乳动物大脑的基本生物学价值都建立在相同的基本计划之上,这一计划体现在创造意识的情感回路中,这些回路集中在皮层下区域,远低于人类高度发达的新皮层”思考帽”。没有这个基础,精神生活是不可能的。在那里,在我们与其他哺乳动物共享的祖先大脑网络中,几盎司的脑组织构成了我们情感生活的基石,产生了我们内心感受情感上的好或坏的许多原始方式。随着我们成熟并了解自己以及我们生活的世界,这些系统为进一步的精神发展提供了坚实的基础。这些皮层下大脑网络在所有哺乳动物中非常相似,但并非在所有细节上都完全相同。这种相似性甚至延伸到某些鸟类物种,例如,它们也有分离痛苦恐慌(PANIC)网络——一个悲伤(GRIEF)系统,我们在这里经常这样称呼它——这是它们大脑和我们大脑中心理痛苦的主要来源之一(见第9章)。
我们哺乳动物和鸟类共享许多其他基本情感系统,有些甚至似乎存在于冷血爬行动物中,但对它们了解较少。因此,在许多温血脊椎动物物种中,各种基本情感网络在解剖学上位于相似的大脑区域,这些网络具有非常相似的功能。我们将讨论通过对其他动物(以下简称”动物”)的研究所揭示的这些大脑系统的性质。这些知识开始让我们了解人性的更深层面。它提供了一个基于科学的关于心智起源的愿景。
正如序言中简要提到的,哺乳动物大脑的古老皮层下区域至少包含七个情感或情绪系统:寻求(SEEKING)(期待)、恐惧(FEAR)(焦虑)、愤怒(RAGE)(生气)、欲望(LUST)(性兴奋)、关怀(CARE)(养育)、恐慌/悲伤(PANIC/GRIEF)(悲伤)和玩耍(PLAY)(社交快乐)。这些系统中的每一个都控制着不同但特定类型的行为,并伴随着许多重叠的生理变化。据我们所知,这些系统也产生不同类型的情感意识,其中一些最有说服力的数据来自人类(Panksepp, 1985)。正如我们将看到的,当这些系统在人类中受到刺激时,人们总是会体验到强烈的情感感受,大概当这些系统被生活事件正常激活时,它们会为人们产生关于发生在他们身上的事情的丰富记忆和思考。
情感神经科学的三角验证方法(本章后面将讨论)为这些系统的效应提供了收集所需证据的机会。但为了有效推进,我们需要一种新的语言来描述大脑的情感系统,以匹配我们对这些初级加工心理力量不断深入的理解。这就是为什么我们要将情感系统的名称大写。民间心理学流传下来的日常用语可能会造成对这些初级加工精神动力的误解。大写字母表明,哺乳动物大脑中确实存在各种情感的真实物理且独特的网络。
如右侧大脑半球内侧视图所示([图1.1]),这些情感生成脑区集中在最古老的内侧(中线)和腹侧(腹部侧)脑区,范围包括:(i) 中脑,特别是被称为导水管周围灰质(PAG)的区域,或以前称为”中央灰质”;(ii) 下丘脑和内侧丘脑,与(iii) 高级脑区大量连接,传统上称为”边缘系统(limbic system)“,包括杏仁核、基底神经节、扣带皮层、岛叶皮层、海马和隔区(见[图1.2],该图描绘了隐藏在[图1.1]左半球内的回路);以及(iv) 各种内侧额叶皮层和腹侧前脑区域(例如眶额皮层),为情感反应提供更高级的控制。尽管皮层下”边缘系统”的概念受到了一段时间的质疑,但所有人都必须承认,与一些早期观点(例如詹姆斯-兰格理论(James-Lange theory))相比,它是一个巨大的进步,后者将情感定位在高级脑区。
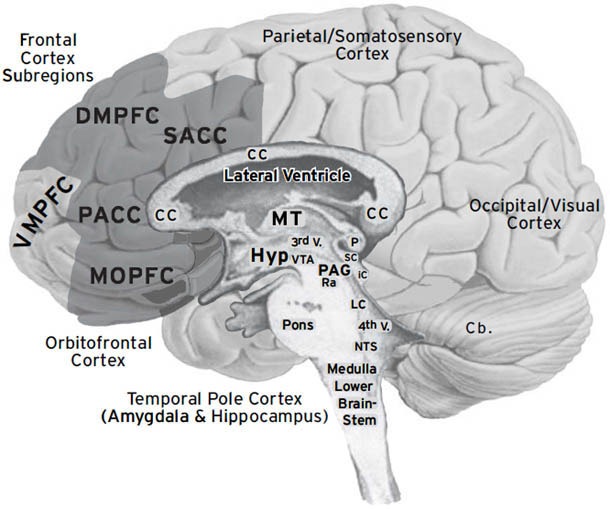
图1.1. 人脑内侧视图(右半球),突出显示了大脑的一些主要区域。从前到后的缩写如下:DMPFC:背内侧前额叶皮层;SACC:上前扣带皮层;VMPFC:腹内侧前额叶皮层;PACC:膝周前扣带皮层;MOPFC:内侧眶额皮层;CC:胼胝体;MT:内侧丘脑;Hyp:下丘脑;VTA:腹侧被盖区(中脑边缘多巴胺系统的来源,支配基底神经节和内侧前额叶区域;见[第3章]);P:松果体;sc:上丘;ic:下丘;PAG:导水管周围灰质;Ra:中缝背核(支配边缘系统的主要血清素系统来源);LC:蓝斑(上行背侧去甲肾上腺素通路的主要来源,供应整个前脑);NTS:孤束核(来自内脏经迷走神经的主要内部受体系统的位置);Cb:小脑。(我们感谢Georg Northoff提供这张大脑视图。)
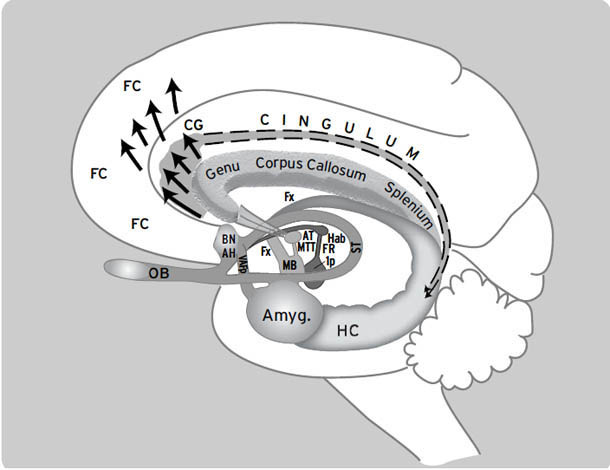
图1.2. 边缘系统示意图,Papez环路用点画突出显示。FC:额叶皮层;CG:扣带回;OB:嗅球;BN:终纹床核;AH:前下丘脑;VAFp:腹侧杏仁逃逸通路;Amyg.:杏仁核;HC:海马;Fx:穹窿;AT:前丘脑;MB:乳头体;MTT:乳头丘脑束;Hab:缰核;FR:后屈束;ip:脚间核;ST:终纹(来自Panksepp, 1998a;经牛津大学出版社许可重新发布)。
据我们目前所知,原始情感系统由神经解剖学和神经化学组成,在所有哺乳动物物种中都非常相似。这表明这些系统在很久以前就已进化,并且在基本情感和动机层面,所有哺乳动物的相似之处多于不同之处。在我们大脑古老的情感深处,我们在进化上仍是亲缘关系。这在我们的身体结构和生物化学中早已显而易见。在各种哺乳动物中都发现了激发这七个情感中介系统的相同类型的神经通路和脑化学物质。根据目前的证据,当这些系统被激活时,人类和其他哺乳动物都会体验到相似的感受。当然,这些感受不可能完全相同,我们也不应该期望它们相同。进化总是为共享的一般原则增加多样性,尽管进化多样化,这些原则仍然为将关键问题从一个物种转化到许多其他物种提供了桥梁。现代医学的许多发现都是基于使用相同推理的动物模型。
正如我们在序言中提到的,这些情感基质是”考古宝藏”——体现我们情感体验能力的多面”珍宝”,这种能力我们仍然与我们的动物近亲共享。然而,作为人类,我们拥有更高级的大脑扩展,使我们能够深入思考我们的本性以及更理性、更具文化性和创造性地生活的选择。我们可以在愤怒时咬住舌头,不说让事情变得更糟的话。但许多人”选择”不这样做。我们在上一句中使用了引号,因为对许多人来说,他们的情感并不受高级思维的意志控制。事实上,有理由相信我们的新皮层功能在很大程度上是由我们的低级思维编程的,结合我们早期的养育,导致幸福的生活(Narvaez等,2012;Szalavitz & Perry,2010)或充满痛苦的生活。
由于我们大脑的高级扩展,我们在认知层面体验生活的方式是其他动物无法想象的。我们可以用微妙的方式反思我们的选择,从而产生越来越微妙的感受,这些感受主要是通过学习建构的。我们独特的心智,在这个世界和宇宙中,源于我们高级新皮层扩展的认知丰富性。但与此同时,我们的高级心智始终植根于我们祖先的过去。可以理解的是,许多人希望将我们的情感生活设想为与我们的认知能力完全交织在一起,但从神经进化的角度来看,这是不正确的。尽管许多认知科学家和哲学家更愿意只思考我们独特的大脑能力,但这根本无助于我们理解心智的起源。但思考我们心智的这些三级加工层面是很有趣的。在那个层面上,我们拥有所有层面相互作用的完整复杂性,这使我们甚至可以以存在性恐惧来思考我们的死亡,或者拥有崇高的感受(Hoffman, 2011)。其他动物不太可能以如此深刻的神经情感焦虑和欣赏深度来体验它们的心智。但它们肯定会体验到它们的原始情绪,也肯定会体验到一些更难理解的其他层面。我们在这里关注的是通过欣赏其他生物的心智,深入到人类心智的最深根源。
尽管神经科学家长期以来对我们大脑的古老情绪回路了解很多,但这些回路直到最近才被明确地与我们的情绪感受联系起来。这使得神经科学家能够深入研究情感的神经基质——我们基本的内部产生的感受的集合。哪些大脑系统给我们带来快乐?我们为什么有时会悲伤?为什么有时有些人总是悲伤?我们如何体验热情?是什么让我们充满欲望、愤怒、恐惧和温柔?传统的行为科学和认知科学无法为这些深刻的问题提供令人满意的答案(这不仅仅是因为研究人员没有提出这样的问题)。
情感神经科学通过自下而上的方式重新开始,既不贬低我们独特的人类能力,也为这些断言提供了关于心智起源的新视野和新数据支持。情感神经科学寻求将情感心智与动物大脑联系起来——在(i)主观心理状态(最容易在人类中研究)、(ii)大脑功能(更容易在动物中研究)和(iii)所有幼年哺乳动物为了生存必须在早期展现的自然(本能)情绪行为之间进行三角定位。这种三角定位使我们能够设想人类心理生活的古老基本结构以及我们价值观的深层神经来源——我们原始的情绪感受。
这些知识为我们指明了必须研究的大脑功能,以便理解情绪障碍——在人类和动物生活中引起心理混乱的各种精神综合征。但成熟的经验很快就会用丰富的思想和学习来补充那些进化而来的工具,使整体图景变得非常复杂。然而,我们计划尽可能保持在初级加工层面的分析。这不仅是因为这个层面被那些研究心理学、哲学和人文学科的人所忽视。无意识次级加工的分析已经是行为神经科学的一个强大而成熟的分支(想想恐惧条件反射,我们将在[第5章]和特别是[第6章]中详细讨论)。我们将忽略人类心智的许多高阶(三级加工)方面,但我们会论证所有这些心理奢侈品都必须建立在对基础问题最透彻的理解之上。我们之所以没有达到这种理解,是因为这些问题只能通过动物大脑研究得到很好的阐明。而且现在已经一个世纪了,关于心智如何在动物大脑中产生的讨论和研究非常少。许多研究人员仍然声称动物是没有心智的僵尸,没有可比的脑心智组织,而这种组织在人类中明显导致自我意识([第2章])。
肯定有许多学者可能不同意上述策略。我们将尽量避免在这里进行复杂的学术辩论(那将是无休止的),但我们确实需要让读者了解一下在这个领域有既得利益的许多科学家可能如何回应我们的立场。我们将以通用的方式来做这件事,通常不会指向任何仍然在世的特定人物。有兴趣追求这个领域多样化视野细节的读者可以参考Jaak Panksepp的其他出版物,他已经多次参与这些问题的讨论。一个强调该领域众多观点的优秀补充读物包含在情绪的本质(Ekman & Davidson编辑,1994)中。
目前在心理学领域,存在一场关于我们是否拥有”基本”情绪的争论,一方相信基本情绪论,另一方则倾向于情绪生活的”维度”观点。要清晰了解这场辩论,Zachar & Ellis (2012) 编辑的即将出版的文集可能特别有用:在该文集中,完整阐述了Panksepp的观点以及波士顿学院James Russell教授的观点,后者一直倡导情绪生活的维度观。维度观认为,一个统一的双价(从积极到消极的效价,以及高低唤醒维度)源自一个称为核心情感(Core Affect)的大脑过程,这是我们情绪本质的基础。这场辩论还得到了不同评论者的补充观点。这种维度观催生了大量优秀的研究,包括最近对动物如何做出复杂的情感相关认知选择的精细动物情绪研究(Mendl et al. 2010)。这种方法现在可以通过情感神经科学策略来补充,将研究发现与脑心(BrainMind)内的神经进化控制层次联系起来(参见Panksepp, 2010a对Mendl及其同事的评论)。这种混合方法对于理解心脑(MindBrain)的全部复杂性至关重要。
我们使用这两个术语——mind和brain,双重大写并以两种顺序排列,以强调情感神经科学是彻底一元论的,没有任何残留的二元论观点。当我们采取自下而上的视角时,更常使用”脑心(BrainMind)“这个术语;当我们采取自上而下的视角时,则使用”心脑(MindBrain)“,两者对于理解大脑进化层次内的”循环因果关系”都是必不可少的。双重大写且不带空格,也强调了将大脑——有些人喜欢称之为”心智肉体”——视为一个统一器官的必要性,没有将心智和大脑视为独立实体的二元论观点的残余,这种知识传统只会阻碍我们的理解(参见第二章)。同时,这个术语的两个版本强调了(i)大脑的某些方面对我们拥有的心智内容类型是内在的(脑心),而(ii)另一个版本则强调在这个器官的上层区域,大量的学习和思维,通常受到社会和文化影响的引导,产生的复杂性可能无法通过动物研究来阐明。
因此,我们有更高级的大脑功能——如今通常被设想为计算认知心智——需要与更普遍的情感心智区分开来。情感和认知方面的心智之间的这种区分虽然不受欢迎,但可以通过多种方式得到支持(图1.3)。将心理治疗建立在对情感过程的知识基础上,从而理解如何最有效地调动有益的认知视角,这一点很重要(Panksepp, 2010b)。
大脑和心智是独立实体的观点是勒内·笛卡尔的最大错误,借用Antonio Damasio (1994)的著名说法。笛卡尔的另一个重大错误是认为动物没有意识,没有体验,因为它们缺乏构成人类心智的那种微妙的非物质材料。这种观念在今天仍然存在,表现为相信动物不会思考甚至不会感受它们的情绪反应。大多数研究动物大脑的人还没有学会如何像他们研究习得行为那样系统而出色地讨论和研究动物的心智,特别是它们的情感感受。动物的原始感受最好通过行为学来研究——通过监测它们自然的情绪倾向。我们的观点是,现在是时候开始这段艰难的旅程了,因为它可能比任何其他尝试过的方法更能告诉我们关于我们自己心智的古老基础。

图1.3. 大脑中介导情感和认知过程的脑系统之间主要差异的总结。总体而言,情感系统控制大脑的整体状态,而认知则处理来自外部感官的输入信息。
因此,主要从动物研究中获得的现代神经科学的详细知识表明,区分心智和大脑不再有用,尽管我们确实必须区分心智的类型和大脑的类型:情感感受,心理学家和哲学家试图主要从观念角度来理解,实际上是大脑的功能。但是能够获得神经”机制”(即神经系统实际如何运作的细节)的大脑研究,在人类身上从伦理角度来说是完全不可能的。在动物身上能否合乎伦理地进行这样的研究仍然是一个争论的问题。无论如何,我们相信有确凿的证据表明其他动物确实有情感体验,理解这些系统对于生物精神病学和心理治疗实践都非常重要。因此,我们将自由地使用心脑(MindBrain)或脑心(BrainMind)这两个术语,具体取决于我们希望强调大脑的哪个方面,无论是在人类还是动物中。但我们在这里主要关注的是通过动物大脑研究阐明的心脑的初级过程情绪。
在继续之前,请考虑以下额外的术语澄清:在本书中,我们最关注的是,首先,产生原始情感感受的本能情绪反应,这些是大自然母亲植入我们大脑的;我们称之为初级过程心理体验(它们是脑心(BrainMind)的进化”既定物”之一)。其次,在这个”本能”基础之上,我们拥有各种学习和记忆机制,我们在此将其视为大脑的次级过程;这些已被研究恐惧条件反射的学者特别深入地研究过(见第5章和第6章);我们相信这些中间层大脑过程是深度无意识的。第三,在大脑的顶层,我们发现了多样化的高级心理过程——各种认知和思维,使我们能够反思从经验中学到的东西——我们称之为三级过程。认识到这些控制层级对于理解脑心的完整复杂性有极大帮助(图1.4)。
一旦我们开始认真考虑已存在的证据,我们相信关于心灵地下室中存在许多基本情感感受这一点几乎毫无疑问(Panksepp, 1998a)。这种对情感生活的”基本”观点也长期以来被研究人类面部表情的学者所倡导(Darwin, 1872; Ekman & Davidson, 1994; Izard, 2007)。事实上,最近对人类脑成像的”元分析”,综合了大多数相关研究的证据,也得出了同样的结论(即Vytal & Hamann, 2010)。
许多争论已经出现(例如,Ekman, 1994; Russell, 1994),因为人类研究实际上无法清楚地描绘人类心智的初级情感过程,因为几乎所有关于人类的研究都在三级和次级分析层面进行。但由于初级过程情感的心理力量,那些研究我们面部表情的学者已经足够清晰地看到了基本情感的微光,足以说服大多数人相信我们的情感本质存在某些基本的东西。但他们没有工具告诉我们那是什么。然而,由于动物研究,我们可以确信所有哺乳动物都拥有许多初级过程情感系统,以及其他情感系统(感觉和稳态系统——图1.4)。而且这些系统并不集中在新皮层,尽管它们与我们的高级脑功能有相互关系(图1.5)。
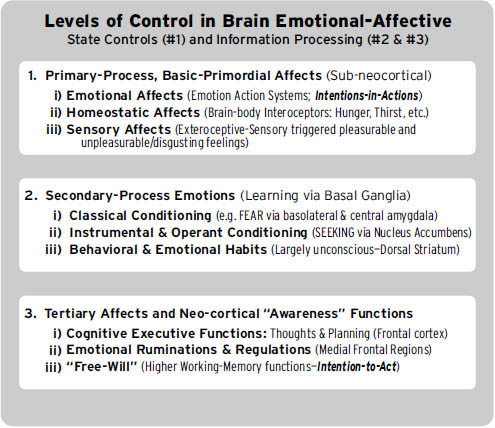
图1.4. 大脑内控制的总体层级概要:(1) 三种一般类型的情感,(2) 三种基本学习机制类型,(3) 新皮层的三种代表性意识功能(完全依赖于通过基底神经节向下循环到丘脑,再循环回新皮层,才能充分阐述思维和行为)。
很少有神经科学家,更少有心理学家在研究所有哺乳动物共享的初级过程情感机制是如何在大脑中构成的。几乎没有人在研究感受(情感)方面。这有助于解释关于情感实际上如何在大脑中产生这一问题长达一个世纪的沉默。相比之下,许多许多科学家正在研究感知功能,如听觉和视觉(关于低级脑感知能力的精彩总结,见Merker, 2007)。对大脑初级过程情感网络的几乎普遍忽视,导致许多人类心理学学者,更不用说社会科学家和哲学家,忽视了他们最亲密的跨学科同事不谈论的问题。
在认识脑心内的进化层级时,关于大脑专业化的一个问题至关重要:出生时,我们心脑的新皮层”思维帽”在很大程度上是一块白板,经验在那里”自然地”印记了许多能力和技能。这些印记包括看似”硬连线”的大脑功能,如我们复杂的听觉和视觉能力。在新皮层层面,这些能力是通过在世界中生活的过程构建的,而不是由任何严格的遗传指令构建的。在众多关键证据中,最令人信服的如下:如果我们在出生前消除”注定”成为视觉处理区域的皮层区域,完美的视觉功能会在皮层的相邻区域出现(Sur & Rubinstein, 2005)。皮层下(例如,丘脑)的影响,可能直接来自外侧膝状核(LGN)的视觉投射,或者可能来自皮层本身的化学梯度,足以使大脑表面发展出视觉能力。顺便说一句,我们可以确信复杂的听觉在脑心进化中是比视觉更古老的过程。这是因为在中脑层面,听觉处理的中央车站——下丘,投射到丘脑的内侧膝状核(MGN)——比中脑视觉处理中心(上丘,投射到LGN)更靠下(更尾侧,意味着更古老)。这也可能有助于解释为什么从触觉进化而来的听觉,比视觉是一个更情绪化的感官。

图1.5 一个关于如何思考动物可能具有的情感特质的真相图表。二十世纪的大部分时间都认为右下角在哲学上是正确的位置,这样可以避免第一类错误(Type I errors),即将不真实的结论误认为是科学正确的。这导致了讨论动物的”类焦虑”行为,而非动物的真实恐惧。本书基于数据得出的结论是,科学家应当明智地将自己置于左上象限,因为这样我们可以避免第二类错误(Type II errors),即由于错误信念或评估现象存在的方法不足,而错过对真实现象的检测。
目前,我们用来粗略”判定”大脑系统年代的这一原则只是一个经验法则,并且存在例外。例如,来自新皮层的更现代的向下影响确实穿透了大脑的许多古老层次。也许最显著的例子是大脑中最长的通路——皮质脊髓束(cortico-thalamic tract)。这条通路从大脑额叶区域的运动皮层一直延伸到脊髓深处,使我们能够随意控制手指和脚趾,这对于演奏钢琴和所有其他乐器、表演舞蹈以及写书都是必需的。
许多情绪研究者以及神经科学同行在情感(affect)和情绪(emotion)之间做出了明确区分,将情绪视为纯粹的行为和生理反应,而不包含情感体验。他们认为情绪唤起仅仅是一组生理反应,包括与情绪相关的行为和各种内脏(激素/自主神经)反应。在他们的科学观点中,动物可能表现出强烈的行为性情绪反应,却实际上没有体验到任何东西——许多研究者认为其他动物可能感受不到它们的情绪唤起。我们不同意这种观点。有些人声称我们将要讨论的系统是深层无意识的——在我们称之为初级过程脑心智(primary-process BrainMind)的祖先体验剧场中什么都没有发生。我们认为证据表明并非如此。
大多数神经科学家愿意承认,许多生理和情绪行为反应是由位于大脑深处的皮层下结构启动的,但他们通常否认或忽视这些相同的结构可以产生原始的情感感受。根据他们的观点,如果动物暴露于危险中,深层大脑结构会产生自动行为(如僵住或逃跑)以及内脏反应(如心率加快和向血液中分泌皮质醇这种通用压力激素)。他们认为这种反应纯粹是生理性的——纯粹的情绪行为而没有任何伴随的情感。这些学者非常乐于声称拟人化(anthropomorphism)——将人类类型的心理过程归因于其他动物——从根本上是不正确的(关于此类问题的精彩讨论,见Daston & Mitman, 2005)。许多其他人选择对这些问题保持沉默,更倾向于谨慎的不可知论立场。我们对所有以情感神经科学方式研究过的哺乳动物的证据解读是,人类和动物的心智建立在基因同源的——进化相关的——情感系统之上,为更高的心智活动提供了许多相似的生物”价值结构”(参见图1.5中需要指导每个人思考这个问题的真相图表)。显然,一些系统会非常相似,而其他系统,特别是社会性情绪(social emotions),会因进化分化的选择压力而差异更大。
原始情绪对成熟的人类来说并非日常现象,但大多数人都能记得在愤怒中握紧拳头、脸红,感到极度恐惧,以及感受到深深的悲伤和喜悦。我们在这里的任务是分享关于这些初级过程心智机制的证据,其中大部分来自对动物的研究。这些感受创造了一种充满能量的意识形式——充满情感强度的意识——我们将其称为情感意识(affective consciousness)。原始感受本质上并不明亮和聪慧,但它们被构建到我们的大脑中,因为它们对于立即应对世界和了解其潜力非常有用。原始情感是帮助我们生存的祖先记忆。这些古老的大脑网络可以让我们以许多方式感受——我们有时称之为核心情绪情感(core emotional affects)和原始情绪感受(raw emotional feelings)的体验。无论我们使用哪个术语,我们讨论的都是同一件事。
研究人类的认知科学家倾向于声称情感感受源自人脑的某些最高级区域。许多和我们一样对人类心理学感兴趣的科学家认为,情感(affects)是当人或动物能够从认知上理解情绪的外周生理变化时产生的。换句话说,情感是由对身体反应的认知反思所定义和衍生的,而不是大脑本身固有的。按照这种观点,如果一个人感到胃部翻腾或拳头紧握,高级认知大脑(新皮层,neocortex)会解释这些原始的生理反应,当它们通过感觉神经进入大脑时,并将这些感受标记为情绪。据称只有在那时,人才会有焦虑或愤怒的主观体验。这就是著名的詹姆斯-兰格情绪理论(James-Lange theory),该理论在一个多世纪前就被提出了(见第2章)。现在我们知道,大脑本身通常会引发伴随情绪的身体唤醒。但尽管如此,一些同事进一步断言,情感只有在我们能够实际用语言表达它们时才会产生——感受源于我们将心智的无意识力量概念化的能力。由于新皮层(大脑的外层)是认知和语言的所在地,这些认知/语言理论认为,情感是在新皮层”读出”位于大脑内部的情绪生理控制时产生的。对他们来说,我们将关注的大脑深层部分无法产生任何体验。我们认为证据表明并非如此。
读出理论(read-out theories)隐含地将意识等同于认知——我们对自己感受和伴随思想的自我意识觉察。如果相信意识总是认知性的,那么情感也必然在某种程度上是认知性的。根据读出理论,情感意识不能从产生情绪的生理变化和本能行为的深层大脑功能中产生,因为这些深层基质是非认知性的,因此必然是深度无意识的。情感只能从严重依赖大脑最顶层(我们的新皮层)的有意识思维中产生,而新皮层对我们所有的高级认知活动都是必不可少的。然而,大量的动物研究和许多临床观察反对将意识与认知等同起来。如果接受情感感受作为意识的一种基本形式,有许多方法可以将这些心智状态与构成认知意识、人类理性基础的信息处理区分开来(图1.3)。
这里有一个极端的例子:基本上没有大脑半球(他们是无脑畸形,anencephalic)因此基本上没有新皮层的人类婴儿,在智力上会保持不发育状态,但如果他们在养育性和社交参与的环境中长大,他们可以成长为情感充沛的儿童(Shewmon et al., 1999;有关这类儿童的照片,见图13.2)。正如我们将看到的,许多去皮层实验(decortication experiments)已经在实验室动物上完成。对于外行来说,这些动物与正常动物无法区分。事实上,它们比正常动物更情绪化。由于这些儿童和动物几乎没有新皮层,他们的情感能力必定源于下方的其他大脑部分。这是科学中最接近证明的例子,而在科学中,结论通常受制于多种可能的解释。革命性的神经学家和神经心理学家现在指出,如果没有允许它们运作的低级皮层下系统(subcortical systems),即使是我们的高级认知心智也无法工作(例如,Damasio, 2010; Koziol & Budding, 2009)。我们的观点也是,心智的古老情感基础对许多高级心理活动至关重要。简而言之,要理解整体心智,我们必须尊重在大脑进化中首先出现的祖先心智形式。
不用说,失去说话能力甚至用语言思考能力的失语症中风患者(通常是由于左侧新皮层损伤)也会保留他们的情感能力,这表明情感意识独立于语言。因此,临床观察表明,认知能力和用语言思考的能力都不是情感意识的必要条件。感受体验可以是无知性的(anoetic)——一种非反思性的、不经思考的初级过程(primary-process)意识,它先于我们对世界的认知理解,或我们所谓的知性(noetic)(基于学习、知识的)次级过程(secondary-process)意识。继续用受人尊敬的神经心理学家恩德尔·图尔文(Endel Tulving, 2002, 2005)的话来说,这让我们拥有自知性(autonoetic)三级过程(tertiary-process)思考意识——在我们的心智中进行时间旅行以及向前和向后看的能力。
这一观点包含了一个激进的主张:初级加工过程的核心情感是非认知的(anoetic,缺乏外部知识),但却以一种情感形式强烈地有意识(被体验到)(这反映了内在的、非反思性的大脑”知识”)。当我们感受到自己的情感状态时,我们不需要知道我们在感受什么。换句话说,初级加工过程的情感感受是原始情感,它们会自动为我们做出重要决定,有时是不明智的决定,至少从我们高级认知心智的角度来看是这样。在有行为规则的文明社会中,情感的外显行为往往是不受欢迎的。尽管如此,产生这种情感感受的能力是大脑进化中的一个关键事件,它使更高形式的意识得以出现。完整的有意识觉知(awareness)肯定要等到我们有了足够的大脑皮层,尤其是额叶区域,才能让我们以自知的(autonoetic)、执行的、决策的能力进行思考。但所有这些精细的心智机制仍然受到我们情绪的强烈影响。情感感受向高级大脑传达的内在评估使人类和动物能够确定他们在生存方面做得有多好或多坏。但有时,它们只会给我们带来麻烦。如果这种情况持续发生,心理治疗通常会非常有用。
另一个有助于设想这些心智进化层次的方法总结在图1.6中。在左侧,我们设想这些层次在早期发展中的”量级”——婴儿起初几乎完全是初级加工过程意识,但随着婴儿成熟并成长为成人,那些祖先价值”似乎”变小了,因为我们的高级大脑充满了知识和观点(在图的右侧)。大多数心理学家试图处理心智的上层,也通过研究基本的学习和记忆过程来处理中层(第6章)。神经科学家是唯一能够阐明心智机制的科学家群体——了解我们如何体验自己和世界。遗憾的是,迄今为止很少有人试图阐明意识的情感感受方面,而这对于理解人类情绪问题和精神障碍可能特别重要。
我们这里的主要目标是处理那些初级情绪过程的本质,它们是大脑心智装置的基础支柱。在生命早期,初级过程指导婴儿做什么和感受什么;在成熟期,获得的高级大脑功能似乎完全处于控制之下——正如每个心理治疗师所知,情况很少如此。我们只会简略地涉及高级情绪和认知过程,但很明显,如果没有它们所建立的坚实的情感/进化基础,那些高级大脑功能就会崩溃。这种层次结构方案使我们能够轻松处理该领域的一些传统悖论。例如,人们经常问为什么人类喜欢看恐怖电影。答案很简单:在心智活动的最高三级加工过程层次——例如,自知意识——我们可以在实际上安全的情况下通过操纵我们的初级加工过程系统而获得极大的娱乐。我们也可以享受雷暴;然而,大多数动物会颤抖。如果没有这种更高的反思过程,我们人类不太可能”自愿”将自己暴露于能够触发负面情感(如恐惧)的感知中。我们也可以确信,我们的思想常常跟随我们的感受。最早的证明之一很简单:当人们被诱导感到快乐或悲伤时,他们的思想倾向于跟随他们的感受(Teasdale et al., 1980)。这是一个普遍的观察。但这并不意味着表征快乐和悲伤的感受来自我们的高级大脑。没有证据表明这种原始感受是由新皮层”读出”的。但这种信念仍然存在。
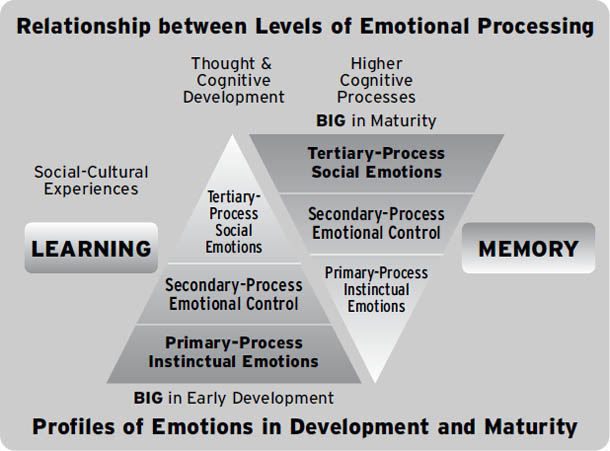
图1.6. 总结婴儿脑心(BrainMind)内控制层次的图表,其中本能的初级加工过程情绪反应非常突出,而高级心智过程尚未发育。这可以与成人的心脑(MindBrain)组织进行对比,在成人中,高级心智过程(三级过程)发育良好,但初级过程受到抑制,这可能表明初级过程对心智生活只有适度的影响,或者它们仍然相当有影响力,但在教养良好的个体中,受到高级心智的调节。
读出理论意味着情感只能发生在足够聪明以解释情绪生理学的动物身上,或者发生在有语言的动物身上。这意味着只有人类,也许还有一些其他灵长类动物是情感生物。据推测,智力较低的哺乳动物在没有欲望的情况下交配,在没有愤怒的情况下攻击,在没有恐惧的情况下畏缩,在没有感情的情况下养育。它们无法感受到社会失落的刺痛——心理痛苦。这可能是对主流观点的极端描述,但在那些实际进行动物大脑研究的人中,这与事实相差不远,因此(据推测)他们应该深切关注这些问题。
尽管有,或者也许正是因为,近期时代思潮的变化——从动物权利运动到关于动物情感的畅销书——大多数神经科学家在他们研究的动物情感话题上仍然坚定地持不可知论态度。许多人说,如果你不能直接测量情感,那么你就不应该讨论它。但我们可以测量核心情感。我们只需要采用间接方法,例如确定某些古老脑系统的人工诱导唤醒(可以通过局部脑刺激来实现)是否能在各种学习任务中充当”奖励”或”惩罚”。事实上,本书始终强调的一个一般原则是,作为动物存在独特情感体验的最有力证据,每当我们通过直接的脑操作唤起本能的情绪行为模式时,动物都会将这些人工诱发的内部状态视为可以导致接近和逃避学习的奖励和惩罚。这样的证据为其他动物的情感心理状态提供了严格的支持。它还告诉我们,在理解这些感受如何由神经网络构成之前,我们需要更详细地理解哪些脑区。
脑科学家必须学会如何有效地使用这些证据,就像那些已经如此详细地研究自然本质的人所做的那样。如果物理学家忽视了自然界这些相对隐藏的方面——可以说采取了鸵鸟政策——我们可能就不会经历导致悲剧性核弹头的量子革命。虽然理解动物的原始情感可能不会那么具有爆炸性,但它将改变我们科学家讨论人性及其各种精神疾病的方式。它可能会改变我们对心智进化的看法,将其明确视为一个自下而上的过程,最终允许自上而下的控制([图1.6])。但整整一代行为神经科学家必须学会如何明确地谈论他们所研究动物的内部情感状态。对于参与这些传统上被回避的话题的全面讨论,仍然存在重大阻力。
在后面的章节中,我们将论证,现在最可信的观点是,(i)原始情感感受、(ii)本能情绪行为和(iii)伴随的内脏反应的各种形式,都是由至少七个”相对”独立的皮层下系统协调的——这些系统分别负责寻求(SEEKING)、恐惧(FEAR)、愤怒(RAGE)、性欲(LUST)、关怀(CARE)、恐慌/悲伤(PANIC/GRIEF)和玩耍(PLAY)。我们说”相对”是因为这些系统中的许多都有重叠的控制:例如,由著名的神经递质如乙酰胆碱、去甲肾上腺素和血清素介导的通用唤醒/注意力促进系统——它们的细胞体大量集中在脑干深处(见[图1.1],其中提供了人脑中几个关键群体的大致位置)。
我们还必须强调”相对”,因为最大的系统,如寻求系统(SEEKING),对其他情绪系统的运作至关重要。我们寻求许多事物并以许多方式进行寻求,因为这个系统指导着多种类型的预期学习。据我们所知,寻求系统(SEEKING)以及所有其他情绪系统在所有被研究的哺乳动物中都非常相似。其他动物的感受肯定与人们在使用各种日常术语(愤怒、焦虑等)时所谈论的感受不完全相同,这些术语通常与特定的生活事件相关,但它们必定非常相似,因为这些感受是由相同的脑区产生的,并涉及相同的神经递质和其他脑化学物质。因此,我们将讨论的存在于其他动物中的核心情感必定与人类体验的情感有很强的对应关系。
但还有其他类型的情感,我们不称之为”情绪”(包括各种身体状态的脑表征,如原始的饥饿(HUNGER)和口渴(THIRST),即稳态情感,在日常用语中包括尿意和便意)。此外,还有外部刺激引发的感觉的愉悦和痛苦(例如,甜味和苦味,以及其他感觉情感如厌恶(DISGUST)和许多其他,包括不同类型的疼痛)。我们在这里不会详细讨论这些稳态和感觉情感。这些话题的行为方面已经受到行为神经科学家的大量研究关注,尽管几乎没有提到它们可能也伴随着情感状态。那么,除了它们如此有趣这一简单事实之外,我们为什么要关注情绪感受呢?这是因为对这些类型的情感的研究对于理解人类精神疾病最为重要,它还将使我们能够拥有有效的人类情绪问题的动物(“临床前”)模型。如果我们只谈论动物的行为变化,而不谈论它们的感受以及这些感受在大脑中是如何被控制的,我们就无法取得那么大的进展。
总之,我们的观点是,谨慎地接受情感是所有哺乳动物情绪表达的重要组成部分,而不仅仅是少数物种的认知后续思考,这是明智的。我们是说动物的感受与我们完全相同吗?当然不是!多样性是进化的规则。当然,每个物种的大脑和身体过程的所有细节都有很大差异。事实上,即使是同卵双胞胎,在其神经系统的精细结构上也不相同。当原始情感与我们更高的心智能力混合时,必然会产生许多进一步的变化和排列组合——这些将创造出复杂的社会情绪,如嫉妒、内疚、妒忌和羞耻,以及敬畏、希望、幽默……甚至体验崇敬和崇高的能力(Hoffman, 2011)。我们可能永远无法科学地知道动物是否有这样的高级情感,因为这需要我们了解它们的思想,而我们还做不到这一点,不像我们能够如此自信地解读它们的情绪感受。当然,一些高级情感对于不同的复杂生物来说是独特的,特别是那些像我们一样拥有脑力深入思考和谈论自己存在的生物。
在生活的正常过程中,特别是在童年时期,情感与更高认知能力的发展交织在一起。这是由于原始情感基质(我们将重点关注的)与成熟的新皮层之间的相互作用。新皮层在不同哺乳动物物种之间的大小和复杂性差异很大,导致认知能力和智力的水平和类型相当不同。如前所述,高阶情绪在不同哺乳动物物种之间必然存在巨大差异。大多数复杂情绪(认知上精心构建的、社会建构的”混合情绪”,在人类中非常常见——想想羞耻和蔑视)尚未经过任何详细的神经科学分析。嫉妒和内疚的现实实验室模型尚不存在,尽管在妒忌等感觉方面正在取得一些进展(Panksepp, 2010c)。由于技术的进步,如功能性磁共振成像(fMRI)脑扫描,我们现在甚至可以成像人类心智大脑(MindBrain)中这种微妙的高级心理过程。妒忌在男性和女性大脑中呈现出不同的图像(Takahashi et al., 2006),男性的妒忌更多地产生于较低的情绪脑区,而女性的妒忌则出现在较高的皮层区域。也许这表明女性的妒忌更多是一种认知反应,基于对她们在经济上可能失去多少的评估。男性更关心性方面的问题。值得注意的是,当对”低等”灵长类动物(恒河猴)进行妒忌的脑成像研究时,让一只占优势的雄性观看顺从的动物与他的配偶交配,大脑的激活类似于前述人类研究中观察到的(Rilling et al., 2004)。很容易想象男性的妒忌是寻求(SEEKING)、欲望(LUST)、恐惧(FEAR)和即将到来的悲伤(GRIEF)等感觉的混合(Panksepp, 1982, 2010c),但这目前只是一个理论推测。
在我们自己的智慧物种中,复杂的想法与情感交织在一起。其他动物不同的认知能力无疑会创造出不同的高级心理景观。然而,同源的情感基质位于皮层下脑的深处,在解剖学和神经化学上与新皮层是可区分的,并且在所有哺乳动物中非常相似。这些事实表明存在着能够在哺乳动物物种中产生各种相似的初级过程情感体验的系统。大多数复杂的社会情绪可能是通过学习,由更原始的情感动力学与认知态度结合而产生的。也就是说,初级过程的情感肯定控制着次级过程的学习机制,然后这两者与更高的认知结合成为大多数心理学家关注的第三级过程心理景观。目前,人们对其他动物的复杂学习甚至更高的心智能力非常感兴趣,但这些有趣的工作很少与大脑研究联系起来。
由于情感与复杂想法和个人经历在我们前瞻性和回顾性的自我意识(autonoetic consciousness)中的交织,我们人类常常难以想象情感可以独立于它们发生的更高心理背景而存在。我们常常发现很难以最纯粹的形式来概念化感受。在我们生活的详细认知背景中看待它们要容易得多。我们认为某个特定的人让我们感到愤怒,或者一次可怕的经历导致我们体验恐惧。(用哲学术语来说,这意味着情感是有意向性的——它们总是”关于”某事。它们是由”情绪评估”产生的”命题态度”——我们在这里只会简单考虑这些问题。)由于大脑高度互联的方式,我们将想法和情感体验为完全交织的体验,而且因为我们是高度认知的生物,我们倾向于将认知视为主要的,假设情感是由思想或感知创造的。仍然有一些心理学家断言,生活经历教会我们拥有情感,如果没有这些经历,我们就不会有情感能力。他们声称,以前从未遇到过危险或痛苦情况的人将无法感到害怕。对于这些理论家来说,情绪在很大程度上是习得性反应。
但在初级过程层面,情绪并不是个体学习的结果。它们是通过进化构建到大脑中的:它们是祖先的”记忆”。据我们所知,我们天生就具有完整的七种基本情绪的先天神经能力,这些能力硬连接在所有哺乳动物大脑的皮层下网络中。我们在使用局部刺激特定大脑区域等技术的动物研究中清楚地看到了这一点。例如,如果以电刺激或化学刺激的形式向产生恐惧的系统提供人工唤醒(从杏仁核到中脑中心——导水管周围灰质[PAG]的长通路,在第5章中有进一步描述),即使是年幼、缺乏经验的动物也会畏缩,如果刺激足够强烈,它们会试图惊恐地逃跑。它们还会迅速学会关闭这种大脑唤醒,并避开曾有过这种经历的地方。然而,人工刺激并不提供任何关于环境的信息。因此,体验恐惧的能力,以及其他基本情感,都独立于任何环境经验。从某种意义上说,感受情感的能力在很大程度上是”无对象的”——最初只有少数刺激能够开启这种大脑-心智状态,但这一刺激阵列会通过学习迅速扩展(见第6章)。
恐惧是哺乳动物大脑的先天能力。然而,恐惧就像所有其他基本情绪一样,随着它受到学习的调节并在我们的意识心智中编码,迅速与世界事件交织在一起。因此,至少在人类中,我们的基本情绪与关于世界的意图和思想(哲学家们如我们所指出的,称之为”命题态度”)交织在一起,结果我们对世界的评估就能产生感受。
大多数基本情绪不需要在出生后立即表达。有些情绪,包括关怀、性欲和玩耍(在不同物种间更具变异性),会在其他情绪(如寻求、愤怒和恐惧)之后很久才上线。但所有这些情绪都有基因硬连接的神经基质。在一些哺乳动物中,恐慌/悲伤反应在生命早期就变得活跃(如那些出生时非常成熟或早熟的食草动物);在另一些中,它会在后期变得活跃(如大多数出生时非常不成熟或晚熟的食肉动物)。在另一些动物中,比如在实验室中繁殖了数百代的实验室大鼠,某些情绪原型(实际上,也许只是它们的行为表达)由于自然(进化)选择压力的大幅放松而变得退化。例如,大鼠和小鼠不像大多数其他哺乳动物那样有强烈的分离叫声,这可能是因为无意中选择了可以单独饲养而不会太痛苦的动物。它们适度的叫声可能只是由身体压力源(如感到寒冷)引起的痛苦叫声。因为我们的基因控制着初级过程情绪,不同物种的情绪气质可能存在很大的变异性,以及为研究而繁殖的不同实验室品系(如小鼠,其中有数千个变体,许多具有独特的个性,其中一些是人工创造的)也是如此(Crawley, 2007)。
虽然体验情感的能力是内置于大脑中的,但在出生时,人类和动物只对少数特定刺激有无条件或本能的情感反应。几乎所有动物都害怕巨大的噪音和疼痛。如果人类婴儿没有被安全地抱住或被允许跌落,他们会哭。几乎所有年幼的哺乳动物如果没有母亲而独自留下都会很快哭泣,但这种反应在许多物种(包括狗和人类)中需要一些时间才能成熟。还有一些本能的情感倾向是特定物种所特有的,因为它们具有感官专长。例如,大鼠天生害怕捕食者(如猫或雪貂)的气味。即使一只大鼠是在圈养中长大的,以前从未接触过捕食者,如果在它的笼子里放一点捕食者的毛皮,它也会变得警惕和害怕。在这种情况下,气味是特定的本能触发因素,或者用行为术语来说,它是引发恐惧的无条件反应(UCR)的无条件刺激(或刺激物,UCS)(如果与任何中性线索,即条件刺激[CS]配对,可以导致经典条件作用——产生条件反应[CRs],正如伊万·巴甫洛夫所发现的,他创造了著名的实验,狗对预测食物的节拍器的声音流口水)。虽然行为主义者认识到厌恶的UCS(如捕食者气味或电击)可以在许多学习任务中充当”惩罚”,但他们可以忽略UCRs(如恐惧)也有内在感觉这一事实,认为它无关紧要。其他UCS可以充当”奖励”,促进接近行为而不是回避行为的学习。然而,传统上很少讨论动物行为学习逻辑背后的任何相应感受。当然,奖励和惩罚之所以能如此有效地控制学习,很可能是因为它们在大脑中产生了情感感受。强化这一神秘过程可能反映了感受在大脑中的作用方式。
随着人和动物在日常生活中经历条件反射和其他学习体验,这些条件性唤起刺激的简短列表很快呈指数级增长。例如,条件反射体验让动物能够对先前对它们来说是中性的刺激产生情绪反应。比如,如果一只猫脖子上戴着铃铛,而一只老鼠与那只猫发生了冲突,老鼠很快就会学会在听到铃铛声时感到害怕并逃跑。更聪明的动物对因果关系(通常存在严重缺陷,我们将在第3章中看到)和时间流逝有认知理解。人类可以灵活运用过去的学习来制定行为,以增强舒适度和生存能力,同时降低不适和死亡的可能性。例如,当人们进行山地徒步时,他们通常会学会携带各种安全设备——充足的水、额外的外套、防晒霜、防水火柴等等——因为他们足够聪明,能够预见和理解各种可能变化的条件可能带来的危险后果。
情感反应(affective responses)以及我们能看到的外显情绪行为,是整个神经科学领域中研究最少的大脑方面之一。情感(affects)以各种特定方式让人感觉良好或糟糕。性满足源于我们的欲望(LUST)能力,其良好感觉与打闹玩耍(PLAY)的快乐或抚摸、养育和照顾(CARE)婴儿的温柔幸福感有很大不同。恐惧(FEAR)是一种与受挫的愤怒(RAGE)完全不同的情绪”痛苦”;两者都不同于社交隔离带来的恐慌(PANIC)痛苦。而在世界中寻求(SEEKING)事物——无论是安全、坚果还是知识——都有一种非常特别、充满活力、有时甚至欣快的感觉,但它也可能制造许多负面事件。
这些多样的愉快和不愉快情感为生活提供指导,因为它们在进化过程中各自赋予了增强生存的优势。情感是我们在生存和繁殖游戏中表现如何的祖先记忆;这些记忆通过我们遗传密码中积累的无意识”智慧”传递下来。引发各种愉快情感的互动——与食物、水、配偶、后代或玩伴的相遇——帮助动物生存和繁殖。引发痛苦情感的生活经历——捕食者、竞争对手、混乱的天气等——会危及生命和繁殖能力。
因此,原始情感为我们最基本的本能行为模式——接近和回避——提供了必要的基础设施,没有它们我们无法生存。人类和其他动物会接近引发愉快情感的事物,并远离让它们感觉不好的事物。因此,情感变化可以强化(reinforce)新的行为模式,尽管行为主义者从未深入了解强化的大脑过程(如前所述,这个术语可能仅仅意味着”情感”——而不仅仅是基本的初级过程情感——在学习背景下如何运作)。动物不一定”知道”或沉溺于这些感觉——这些感觉在大多数物种中可能只是原始的非认知性(anoetic)体验。然而,人类确实对他们的个人经历有许多想法和沉思,这些可以进一步阐述情感,允许认知性(noetic)(事实性知晓)和自我认知性(autonoetic)(自传式时间旅行)形式的情绪体验(总结参见Vandekerckhove & Panksepp, 2009)。其他哺乳动物,甚至像大猩猩和大多数食肉动物这样高度智能的动物,在多大程度上具有这种更高层次的认知(思考性、反思性)意识,肯定是一个比我们正在讨论的问题更困难的问题,我们讨论的是所有哺乳动物中原始情感-情绪体验的存在。
为什么动物情感对理解人类幸福如此重要?因为理解它们为我们提供了关于我们自己基本价值系统的知识——生活中本质上感觉良好和糟糕的方面。我们无法在人类大脑的精细神经层面研究这些过程。为了理解跨哺乳动物物种的情感,使用三角验证研究方法非常有帮助,该方法同等关注我们对(i)哺乳动物大脑、(ii)其他动物的本能情绪行为,以及(iii)人类心智的主观状态的理解。这种三角验证是我们能够研究我们自己物种以及其他动物情感生活神经基础的主要方法(Panksepp, 1998a)。这种方法可以对情感研究基础理解的进步以及生物精神病学和心理治疗的实践产生重大影响。它还首次提供了一种科学理解其他动物体验的方法。
这种三角验证方法的第一个组成部分涉及大脑系统和功能。物理大脑必须始终是严格神经科学研究的主要组成部分。只有当我们了解大脑如何工作时,我们才能深入理解动物和人类的行为和心理过程。然而,在本书的总体涵盖范围内,我们不会像在专业科学论坛上那样深入探讨潜在的神经学、神经化学和神经遗传学问题(有关这些细节,请参见Panksepp, 1998a)。
第二个组成部分是对动物行为的仔细研究,特别是它们的自然(本能)行为倾向——它们的无条件反应(unconditioned responses, UCRs)。大量证据现在表明,产生无条件情绪行为的大脑网络,实际上伴随着情感体验(有意识的、无条件的大脑内部过程,可以在学习任务中充当”奖励”和”惩罚”)。因此,我们可以进一步得出结论,引发动物自然情绪行为的大脑操作也会诱发伴随的情感状态。当然,大脑本可以以其他方式构建。但现在已充分确立的原始情绪情感与本能行为表达之间的对应关系充分证明,情感体验是所有哺乳动物,可能还有大多数脊椎动物情绪唤起的组成部分。
第三个组成部分是心理分析,其中主要包括人类关于情感体验的言语自我报告。人类可以详细讲述他们的感受。因此,如果某个大脑操作在动物身上产生情绪行为,并且如果人类在相似的大脑区域受到刺激时描述相关的情感体验,那么这就补充了动物观察。此外,由于有大量方法可以通过动物回避或追求其神经系统某些状态的倾向来确定它们是否有感觉,我们至少可以确信它们确实有令人向往和不向往的心理体验。例如,我们可以通过实验”询问”动物它们是否会为某些大脑操作工作或回避某些大脑操作,比如对特定大脑区域施加电刺激,或者它们是否会返回或回避它们曾经接受过此类大脑操作的地方。它们的反应提供了我们寻求的答案,特别是当与类似情况下人类可以提供的言语自我报告一起考虑时。
总之,目前关于情绪感受和其他情感如何在哺乳动物大脑中组织的最令人信服的知识来自对特定大脑系统的直接操作。尽管我们无法询问实验动物其体验的确切性质,但如果它们实验诱发的情绪行为是明确的,并且人类在类似唤起时报告明确的情绪体验,我们就有初步证据表明大脑中的情感基础结构比许多心理学家所支持的简单的全局”积极”和”消极”情感更为清晰。我们还可以在动物身上设计辨别测试来确定它们是否区分某些感受(例如,Stutz等人,1974),但该研究领域才刚刚开始。
除了局部的大脑电刺激,特定化学物质可以被应用于动物大脑的特定区域以产生特定的情绪行为。例如,促肾上腺皮质激素释放因子(corticotropin-releasing factor, CRF)——启动大脑-身体应激反应的执行系统——产生恐惧(FEAR)的形式(冻结和逃跑),并且我们有充分理由相信,它在哺乳动物和鸟类中产生惊恐/悲伤(PANIC/GRIEF),因为CRF可以显著增加因社会分离而产生的哭泣。如果我们有理由得出动物情绪行为的变化表明动物情感状态的结论,那么我们可以假设对人类大脑的类似操作会产生类似的情感变化。尽管用局部化学刺激人类大脑的工作很少,但在动物身上进行的大量此类工作对初级过程情感如何在人类大脑中产生具有丰富的启示。事实上,阻断分离痛苦系统的药物处于生物精神病学中新型抗抑郁药开发的前沿(完整概述见Watt & Panksepp,2009)。用以特定方式影响大脑化学的外周给药进行了大量工作。动物和人类数据非常吻合。例如,所有哺乳动物通常会对同类型的药物上瘾。这一知识具有重大实用价值,因为它允许在人类和动物情感体验之间进行直接的神经药理学转化。
我们不会深入探讨大脑的多样化神经解剖学和神经化学,但至少会分享当前思考的简要概述。例如,在所有哺乳动物大脑中,被称为”阿片类物质”的内部类鸦片递质化学物质(这些在功能上类似于吗啡或海洛因等成瘾性药物)在神经细胞之间传递”信息”——有时更好地理解为”存在状态”。例如,β-内啡肽与特定皮层下区域中所谓的μ受体(集中在神经细胞突触表面的大型”监听”分子)结合,以产生各种理想的内部状态——社交陪伴的快乐,或令人愉悦的味觉和触觉。这种内部阿片类物质感应的μ受体可以消除疼痛感,并向大脑发送令人愉悦的满足感信息。正如第8章和第9章中将要总结的那样,第一个被发现由阿片类物质控制的微妙情感满足是我们在所关心的人面前、在情感安全和社交满足时所体验到的成瘾性爱的感觉(Panksepp, 1981a)。最近,这种化学物质被发现能够介导我们对甜食的成瘾(Avena et al., 2008)。我们将在本书中强调许多其他情感例子。事实上,许多自然的快乐可以抵消药物成瘾。最引人注目的发现之一是,母性”点亮”了许多与可卡因效果相同的皮层下大脑区域,其吸引力不亚于这些滥用药物(Ferris et al., 2005)。
递质与其特定受体的结合以”钥匙”和”钥匙孔”的方式发生,其中相对较小的递质分子充当钥匙,而更大的受体分子充当”锁”的钥匙孔,控制神经放电。在大脑的情感区域,这些分子可以解锁我们的感受。用不那么诗意的术语来说,特定的钥匙状分子与特定的受体分子结合,跨越许多突触(神经元之间的信息传递间隙),并可以启动复杂的化学级联反应,导致几种不同类型的情绪唤醒。重要的是要注意,这些情绪化学物质中的许多在大脑中以全局方式起作用——它们在许多大脑区域释放,使各种网络功能受到一个情绪指挥者的协调。目前看来,一些由许多氨基酸链构成的较大递质——神经肽——为我们可以体验到的不同情绪倾向和感受提供了相当的特异性。
神经科学家尚未绘制出神经化学系统活动与情绪表达之间的所有神经学步骤。这将需要很长时间。但现在非常清楚的是,某些大脑化学物质,特别是神经肽,可以产生高度可预测的情绪感受反应。例如,参见第9章中的图9.3,了解促肾上腺皮质激素释放因子(由31个氨基酸组成的递质分子)在激活幼鸟大脑中反映分离痛苦的哭泣类型方面的力量。
正如第9章将广泛讨论的那样,当少量阿片类物质与μ受体结合时,会出现完全相反的感觉。这启动了一个化学级联反应,产生情绪上满足的反应。动物看起来快乐和放松,它们似乎相当自我满足。即使被置于隔离状态,它们也没有表现出任何哭泣的动机,而且一点也不显得困倦;事实上,在平息它们情绪痛苦所需的极低剂量下,这些动物通常更加活跃。它们玩得更多。如果剂量更大,动物确实会变得困倦。在高”药理学”而非”生理学”剂量下,它们表现出紧张性,几乎昏迷的状态。然而,仅仅减少哭泣的微小剂量不会产生任何此类效应,除了在某些新生”早产儿”中,如胎儿状的新生大鼠,在其未发育完全的神经系统中,这种小剂量会产生更大的大脑效应。如果我们假设微小剂量阿片类物质后的满足行为反映了动物的满足感受,那么考虑到哺乳动物物种间皮层下神经网络和功能的相似性,我们可以假设人类会有类似的反应。事实上他们确实如此。这对所有成瘾性阿片类物质都是众所周知的。当人们处于阿片类物质的影响下时,他们说他们感到舒缓和安慰。这是因为他们的恐慌/悲伤系统不那么活跃,这有助于解释为什么孤独、被剥夺权利的人更容易沉迷于这些药物。事实上,如果阿片类物质不那么容易上瘾,它们几乎将是完美的抗抑郁药。现在有更安全、成瘾性低得多的阿片类物质(例如丁丙诺啡),可以用于治疗对其他疗法有抵抗力的抑郁症(Bodkin et al., 1995)。由于大量的动物研究,我们现在可以为控制各种情绪状态的大量神经肽甚至更小的递质分子产生类似的想法。
其中一种分子是多巴胺(dopamine),它由单个氨基酸——酪氨酸合成而来。这种小型递质分子促使动物对其环境进行热情的探索([第3章])。这种情感和行为唤醒可以通过服用称为”精神兴奋剂(psychostimulants)“的药物来实现,这些药物会增加大脑中多巴胺的释放。然后,多巴胺充当与多巴胺受体锁孔结合的钥匙(多巴胺受体有五种主要类型,每种功能略有不同)。许多在突触处增加多巴胺活性的兴奋剂分子——例如苯丙胺和可卡因——在所有哺乳动物中也具有高度成瘾性,尽管它们引发的感觉与阿片类药物不同。
每当大脑中多巴胺释放增加时,动物就会以一种独特的方式变得更加兴奋。它们变得更加渴望和好奇。如[第3章]所述,当这种情况发生时,动物会表现出兴奋的寻求行为(SEEKING behaviors),可以预期环境中所有类型的吸引人的事件。下丘脑外侧区(LH)是动物处于这种兴奋状态时被激活的一个大脑结构。其他的还有大脑上方的伏隔核(nucleus accumbens)以及更上方的内侧额叶皮层(medial frontal cortex)。所有这些大脑区域都通过一条非常大的通路连接,该通路连接大脑的下部和上部区域,称为内侧前脑束(medial forebrain bundle, MFB),它包含许多不同的神经化学网络,其中一些使用多巴胺运作。直接电刺激这些大脑区域中的每一个,沿着MFB的所有位置,也会产生这种兴奋反应。动物喜欢自我激活这些电极位置——它们很容易开始以强迫性、成瘾性的方式自我刺激自己的大脑。
大脑多巴胺系统是几乎所有形式的药物成瘾以及哺乳动物所有自然欲望的基本中介,这不再令人惊讶。当人体内的MFB受到多巴胺或电流刺激时,他们会报告兴奋、兴趣和期待的愉悦感。他们可能变得狂躁。动物很容易返回它们获得这种体验的地点。人类的主观报告使我们可以推测动物也经历类似的情感。当这个大脑系统的活动被抑制时,动物相应地表现出抑郁,而人类报告感到心理上迟缓,对任何事情都没有热情。
从体外引入的化学钥匙(那些从身体外部引入的)能够与受体锁孔结合,但不会引发接收神经元放电速率的变化,实际上会使它们暂时失效,这些被称为受体阻断剂(blockers)或拮抗剂(antagonists)。例如,纳洛酮(naloxone)和纳曲酮(naltrexone)等化学物质可以阻断μ受体。纳洛酮和纳曲酮也会抑制外部阿片类药物(如吗啡和海洛因)以及一些内源性阿片类物质(endogenous opioids)——在大脑内产生的类似阿片的化学物质——的作用。当内源性阿片类物质被阻止与μ受体结合时,动物表现得更加紧张,而且它们似乎不喜欢这种心理影响。人类报告类似的不良情感,但这些变化往往很微妙,需要长期服用大剂量。同样,阻断多巴胺作用的关键分子可以在人和动物中引起嗜睡和抑郁。已经开发出的大量突触受体拮抗剂在研究各种内源性大脑突触神经化学的心理影响方面特别有用。
另一方面,当外源性药剂与受体结合并产生与内源性大脑化学物质相同的结果时,该外源性药剂被称为激动剂(agonist)。在某些罂粟植物(罂粟)中发现的阿片类物质产生与内源性阿片类物质相似的情感感受。两者在情感上都令人舒适。因此,阿片类物质充当内源性阿片类物质的激动剂。还有大量其他受体激动剂药物可以增强大脑许多特定内源性化学物质的作用。例如,可卡因和甲基苯丙胺都通过增加突触处多巴胺的可用性来促进多巴胺活性。
还有许多其他药物在所有哺乳动物中起作用,可以改变神经递质合成或降解的速度,为神经科学家提供了一套令人难以置信的工具,用于在情绪状态的神经、心理和行为分析之间进行三角测量。所有这些药物都可以在动物研究中局部用于大脑。人们还可以直接测量动物在情绪行为时大量神经化学物质的释放。从这些工作中我们知道,几乎在任何使动物行为上兴奋的条件下都会释放多巴胺。其他药物通过作用于其他神经化学系统产生明显不同的行为影响和感觉。
在继续之前,让我们先处理一个大多数读者会好奇的问题。即使是”低等”动物,比如无脊椎动物,也有情感体验吗?它们也会追求对哺乳动物成瘾的药物吗?许多会的。我们现在知道,小龙虾会对它们被给予精神兴奋剂或阿片类药物的地方产生偏好(Panksepp & Huber, 2004; Nathaniel et al., 2009)。这表明情感体验在脑心智(BrainMind)进化中的深度远超哺乳动物发育水平。但也可能有其他解释,无脊椎动物截然不同的神经系统不允许我们像对待其他哺乳动物那样,在它们的行为、大脑机制和心理感受之间进行轻易的三角验证。因此,我们在此不会详述这些有趣的问题,但我们必须始终对很少有人实验考虑过的合理可能性保持开放态度。
尽管神经解剖学和神经化学分析对于跨物种三角验证方法至关重要,但对动物行为的详细研究,特别是动物自发表现出的自然情绪行为,目前是情感神经科学的关键要素。也许在未来,我们对大脑功能的了解将足以使用现代人类脑成像设备(例如,正电子发射断层扫描[PET]和功能性磁共振成像[fMRI]),从我们看到的”图像”中常规预测情感体验。但这在人类或动物身上目前都还不可能。不过,正在取得一些进展。例如,通过对比接收大量多巴胺(dopamine)信息的伏隔核(nucleus accumbens)等大脑区域与介导厌恶感的脑岛(insula)等其他区域,研究人员已经证明,在购物时,人们会决定购买”点亮”其伏隔核的东西,但如果激活了脑岛,就几乎没有购买欲望(Knutson & Greer, 2008)。
不幸的是,fMRI等一些技术要求人类和动物完全静止,这在行为上与强烈的情绪唤醒不相容。PET可以更容易使用;研究人员甚至可以在将动物放入脑扫描仪之前注射正电子发射成像分子。PET已被用于监测猴子”嫉妒”期间的大脑变化(Rilling et al., 2004),但这项技术对于常规动物研究来说过于昂贵。虽然fMRI在越来越多的动物研究中得到有效使用,但动物必须完全固定才能获得任何有用的图像。
必须认识到,大多数人类神经成像研究提供的是大脑较高级、新皮层(neocortical)部分的更好视图,主要是因为这些大脑区域比古老的皮层下结构大得多,而且它们在代谢上更活跃。通常很难可视化细胞放电较慢或只是改变放电模式的皮层下区域(例如,第3章中讨论的多巴胺神经元)。此外,许多可能产生冲突信息的邻近系统重叠更广泛。而且,即使可以可视化皮层下区域,它也不总能提供正在发生的神经细节的清晰图像,因为神经成像技术监测的是整体区域脑活动(例如,血流或糖消耗)。
基本假设是大脑功能需要氧介导(有氧)代谢形式的能量;因此,局部血流、氧合或葡萄糖水平的变化反映了区域脑活动。然而,能量消耗和血流可以是神经元抑制信号以及兴奋的反映——在下游突触产生抑制的神经元放电的产生也需要消耗能量。因此,甚至不可能确切知道人脑中似乎亮起的许多”灯”是反映大脑兴奋(放电增加)还是抑制(下游放电减少)。此外,还有许多统计陷阱,过于复杂无法在此考虑,可能导致对所见效果强度的错误印象(例如,Vul et al., 2009)。对于外行来说,最糟糕的是,极其微小但一致的大脑信号变化被转换成显示器上任意强烈的颜色,这很容易欺骗不知情者相信大脑变化比实际更大。从情感神经科学家的角度来看,也许最麻烦的方面是这些技术并非设计用于观察大脑最古老的区域,在那里神经化学的力量往往比神经放电的绝对变化更有影响力。尽管如此,用人类脑成像获得的数据确实相当壮观。
因此,尽管与最先进的神经影像技术相比,动物行为观察可能显得过于简单,但动物行为提供了非常好且有用的科学数据,因为动物的初级过程(本能)情绪行为可能准确反映了其初级过程情感体验。人类大脑成像在阐明人类的初级过程情绪方面相当薄弱。人类能够思考自己的情感并抑制情绪行为,正是因为他们如此聪明。一般来说,大脑中更深层的情绪部分会唤醒控制我们认知的表层皮质区域,而更高的皮质层通常会抑制和调节从下方产生的情感唤醒。拥有异常巨大新皮质的人类,通常能够抑制伴随情绪唤醒而来的行为。例如,恐惧的人往往可以假装镇定。事实上,人类的大部分社交生活都涉及某种程度的情感抑制和混淆。我们不会去抓取我们想要的东西,我们倾向于淡化胜利和失败的感觉,即使在恼怒时我们也试图表现得友好。动物通常没有这种自我产生的能力来抑制和掩饰它们的情绪反应。当一只老鼠或猴子体验到情感时,它的行为通常反映了它的感受。因此,在我们探索情绪行为和情感感受如何在大脑中产生的过程中,没有任何现代大脑成像技术能够取代对动物行为的仔细研究。
如果情感不是身体生理变化的认知解读,如果它们源自大脑深层的非认知部分,那么情感感觉起来像什么?我们认为情感不像任何其他东西。它们是原初的现象体验,不能仅仅用身体的伴随变化来充分解释,尽管在情绪唤醒期间必然会有许多独特的身体感觉。情绪感受和生理唤醒的交织,很大程度上可能是因为初级过程情绪系统位于大脑中调节我们内脏活动、激素分泌以及注意力和行动能力的相同区域。
可以肯定的是,身体反应也可以影响情绪唤醒。例如,愤怒总是伴随着血压升高。血压也会对情感施加影响,因为任何升高血压的化学物质都会使愤怒的人或动物感到更加狂怒。这是因为动脉中的压力感受器可以直接促进古老内脏大脑区域(即代表我们内部身体器官的大脑部分)中的愤怒(RAGE)回路。然而,人工升高血压并不会在尚未被激怒的人或动物身上产生愤怒。因此,情感似乎不只是反映外周情绪生理。正如我们已经说过的,情感是用于编码价值的古老大脑过程——大脑用于快速判断什么会增强或损害生存的启发式方法。
那些认为语言是情感标志的人离真相更远。词语最适合解释我们周围世界的运作方式。词语可以解释乔治华盛顿大桥连接纽约和新泽西。词语可以告诉你如何烤蛋糕。但词语无法解释原初体验。词语甚至无法解释看到红色这一原初感知体验。像”猩红色”、“深红色”或”宝石红”这样的词语并不描述任何东西。它们只是看到红色变化这一共同体验的标签或符号,这严格来说是一种主观的大脑功能。人们可以使用任何符号,包括非语言符号,作为看到红色体验的标签。“红色”没有内在意义,但红色的体验有意义——它象征着生活中一些最令人兴奋的事物,从水果的成熟到性的激情和流血。词语无法向盲人描述看到红色的体验。
词语也无法描述情感。人们无法解释愤怒、恐惧、欲望、温柔、孤独、嬉戏或兴奋的感觉是什么样的,除非间接地使用隐喻。词语只是我们都曾有过的情感体验的标签——我们普遍认可的原初情感体验。但因为它们隐藏在我们的脑海中,源于我们大脑古老的前语言能力,我们还没有找到连贯地谈论它们的方法。
关于这些系统如何连接到人类更高级的意识能力的科学研究,在很大程度上仍然是未来的任务。然而,由于这些系统对临床精神病学现象的重要性,我们将在每个致力于”七大”情绪情感的章节中简要讨论这些更高级的认知方面。
到目前为止,三角测量法已经揭示了七个基本系统的存在,这些系统在所有哺乳动物物种中都是同源的。我们不知道动物何时开始有情感体验,但目前的研究表明,一些情感已经存在于非哺乳类脊椎动物中。例如,被隔离的幼鸟会经历分离痛苦,就像被隔离的幼年哺乳动物一样(第9章)。此外,正如在早先的小龙虾例子中简要提到的,有提示性证据表明一些无脊椎动物具有情感体验。
我们有理由相信,在哺乳动物出现之前,七种基本情绪系统的完整配置就已经以雏形形式进化出来了。这是因为鸟类具有明确而独特的情绪特性。在所有哺乳动物和鸟类物种中,相似的化学物质会激活和抑制这些系统;根据我们目前的知识,每个系统都会产生独特的情感体验。但这些系统之间存在许多重叠的方面;例如,寻求系统(SEEKING)参与了大多数其他系统。所有系统都受到通用大脑唤醒调节器的调控,如血清素、去甲肾上腺素和乙酰胆碱。这些为我们带来了无法避免的复杂性,就像我们的语言会创造复杂和重叠的含义一样。
语言,尤其是晦涩的专业术语,无法充分描述情感。因此,我们将使用常见的日常用语——简单的词汇——作为七种情绪系统的标签。然而,为了避免混淆,我们将(如前所述)使用全大写字母,以强调我们所说的是独特的大脑系统,特定的情感和情绪从中产生;我们不是仅仅在谈论这些词通常表示的普通感受。
同样重要的是要明确,我们通过情感神经科学的三角测量所获得的知识,并不能解释现实生活中发生的完整情绪体验的复杂性。虽然部分与整体的混淆在认知神经科学中很常见(Bennett & Hacker, 2003),但我们希望在这里避免这种情况。我们所说的是特定的神经系统,它们是我们生活心理整体的重要组成部分。我们并不声称在谈论作为整体实体的情绪。科学仅限于研究现象的各个部分。
只有理论叙述才能将各个部分统一成可理解的整体。例如,达尔文收集化石并观察不同岛屿上的生命。他观察到的各种数据项是科学部分,最终使他能够设计出关于进化的整体理论(适者生存)。我们还没有一个完全统一的情感理论——在这个理论中,高级和低级大脑功能的整合可以在神经和心理细节上得到理解。为了使这种进步成为可能,还需要收集更多的数据。但什么样的数据最有价值?也许最重要的是独特情绪行为的神经机制,以及它们的奖励和惩罚效应。与此同时,现代神经科学的巨大成功产生了越来越多精巧的分析工具,观察神经活动越来越精细的方面,这些工具往往无法以清晰的洞察力应用于全局性的心理问题。因此,我们拥有大量关于神经机制的知识,这些机制正在寻找功能;这是一种相当奇特但在智力上很有启发性的状况。例如,“沉默突触”的功能是什么?是随时待命,等待学习的正确神经条件(见[第6章])吗?这种技术资源的丰富也有其缺点。它促进了一种”无情的还原论”,即对神经机制的研究很重要,但它们产生的体验却不重要。我们在这里不支持这种对心智的忽视。
在本书中,我们将关注通过识别七种情绪大脑基质所取得的实质性实证和理论进展,这些基质能够可靠地唤起独特的情绪行为,并在所有被研究的哺乳动物中产生情感体验。我们并不声称这七种构成详尽的列表。可能会发现更多。此外,关于调节这些系统或系统部分的不同化学物质,还有很多需要学习的地方。我们也还不完全了解情感和其他心理过程是如何从大脑的精细复杂性中产生的。然而,我们的方法确实鼓励以新的方式来考虑这些困难的神经科学和现象学问题。这是可以进行的,因为我们现在确实了解了很多关于基本大脑区域和过程的知识,特别是一些关键的神经化学。
在本书的结尾,我们将提出一个新颖的、也许是革命性的关于情感生成的假设——这个假设依赖于我们设想”核心自我(core-SELF)“的能力,鼓励我们思考”灵魂”的古老神经生物学([第11章])——它在情绪-运动/行动网络中提供了一个重心,使原始情绪感受能够从大脑活动中涌现。在这里,理论(得到一些启发性数据的支持)正在把握心智中相对无形的方面,这些方面仍有待用神经科学工具充分探索。然而,目前我们可以确信,一个或多个七种情绪系统的唤醒是哺乳动物产生情感的必要条件。未来的研究者将不得不解决更多、更多的细节,关于情感实际上是如何在大脑中产生的,以及为了正常工作,这些大脑功能如何与身体的其他部分协同作用。
尽管语言无法完全描述这七种基本情感,但我们会尽力而为,有时会诉诸生理相关性来具体充实它们的含义。在这里,我们提供”七大”的概要。关于这些原始情绪系统以及一些更高级情绪复杂性的有趣描述,请参见[图1.7]。
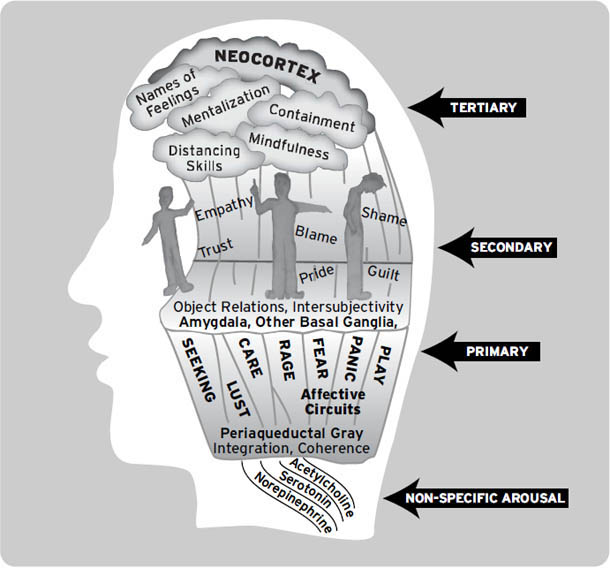
图1.7. 初级过程情绪系统及其各种次级和三级过程后果的示意图。本图改编自Sandra Paulsen为本书绘制的艺术作品,经其许可使用。
RAGE系统(见第4章)与SEEKING系统形成对比,使动物将身体推向冒犯性物体,它们撕咬、抓挠并用四肢猛击。愤怒本质上是消极情感,但当它与认知模式相互作用时可以变成积极情感,例如战胜对手的体验或将自己的意志强加于能够控制或征服的他人。纯粹的RAGE本身不包含这种认知成分,但在成熟的多层哺乳动物大脑中(图1.4),它确实包含。
FEAR系统(见第5章)产生所有人和动物都希望逃离的消极情感状态。它在身体中产生紧张感,在较温和的唤醒水平上产生颤抖的静止不动,这可能会加剧并爆发成动态的逃跑模式,以混乱的抛射运动脱离危险。如果如我们上面推测的,当FEAR系统唤起SEEKING系统时触发逃跑,那么初级过程FEAR的厌恶特质可能最好通过静止”冻结”反应和其他形式的行为抑制,以及积极情感的减少来研究,而不是逃跑。
当动物处于LUST系统的支配下时(见第7章),它们表现出丰富的”求偶”活动,最终移动到与接受的配偶紧急结合身体(图7.1),通常在高潮的愉悦中达到顶峰——这是生命所能提供的最戏剧性和积极的情感体验之一。在没有配偶的情况下,处于性唤起状态的有机体体验到一种渴望的紧张感,当满足即将来临时可能变得积极(可能是因为SEEKING系统的同时唤起)。当满足难以捉摸时,这种渴望的紧张感可能成为情感上的消极压力源。LUST是爱的来源之一。
当人和动物被CARE系统唤起时(见第8章),他们有用温柔的爱抚和体贴的照料包围所爱之人的冲动。没有这个系统,照顾幼崽将是一种负担。相反,养育可以是深刻的奖励——一种被珍视的积极、放松的情感状态。CARE是爱的另一个来源。
当被PANIC/GRIEF(也常称为”分离痛苦”)系统压倒时(见第9章),人会体验到深深的精神创伤——一种没有明显身体原因的内在心理疼痛体验。在行为上,这个系统,特别是在年幼哺乳动物中,其特征是坚持不懈的哭泣和迫切试图与照顾者(通常是母亲)重聚。如果未能重聚,婴儿或幼儿逐渐开始表现出悲伤和绝望的身体姿势,反映了大脑从恐慌级联到持续抑郁的过程。PANIC/GRIEF系统有助于促进积极的社会联结(这个系统的次级表现),因为社会纽带缓解这种精神痛苦并用舒适和归属感(充满CARE的感受)取而代之。因此,儿童珍视并爱护照顾他们的成年人。当人和动物享有安全的情感纽带时,他们表现出放松的满足感。这些感受的波动是爱的又一个来源。
7. PLAY系统(参见第10章)通过轻快跳跃的动作来表达,参与者经常以快速交替的模式互相戳或推搡。有时,PLAY看起来像是攻击行为,尤其是当PLAY以摔跤的形式出现时。但仔细观察这种行为就会发现,粗暴游戏的动作与任何形式的成年攻击都不同。此外,参与者享受这项活动。当儿童或动物玩耍时,他们通常会轮流扮演主导和顺从的角色。在对照实验中,我们发现一只动物会逐渐开始战胜另一只(可以说成为头狗),但只要失败者仍有一定百分比的机会最终获胜,游戏就会继续。当头狗和弱势者都接受这种让步方式时,参与者就会继续享受这项社交活动。如果头狗想一直获胜,这种行为就接近欺凌。正如我们将在第10章中看到的,即使是老鼠也会通过情绪化的发声清楚地表明它们在游戏活动中的立场:当它们被剥夺获胜的机会时,它们快乐的笑声般的声音就会停止,情绪化的抱怨就会开始。PLAY系统是友谊的主要来源之一。
再次重申,这七个系统被认为是情绪系统,因为每个系统的唤醒都会产生强烈的内脏、行为和情感反应。例如,催产素(oxytocin)以及其他一些化学物质在CARE系统中产生母性行为方面起着关键作用,同时也减少了PANIC/GRIEF系统的分离痛苦。在正常情况下,在怀孕结束时会内源性地产生大量催产素混合物。它会在分娩时引起子宫收缩,并在分娩后促进乳汁分泌。这两种反应都是CARE系统被唤醒时发生的内脏组成部分。然而,大脑中还有一个心理奖励。当动物的大脑充满催产素时,它们会变得不那么具有攻击性,更加自信和更有养育性。
如果给处女大鼠注射催产素,并发生其他几种生理变化,她就会表现出CARE行为和感觉的唤醒。她会寻找幼崽来养育;她会开始为它们筑巢;她会盘旋在它们上方提供温暖;当它们走失时,她会把它们聚集起来。这些都是在产后母鼠身上看到的典型CARE行为。我们从口头报告中得知,产后人类母亲的大脑分泌类似的催产素混合物,对婴儿感到温柔和强烈的保护冲动。这些是CARE系统被唤醒时发生的情感反应。但催产素这种在婴儿哺乳时释放但也可能因各种压力源而升高的激素,是主要原因吗?人类研究可以解决这个问题,但只能在心智的三级过程层面上。动物对初级过程的研究是否有助于为原始情感原则提供关键的清晰度?让我们详细考虑这种可能性。
在动物中直接进行脑内注射催产素的工作已经进行了三十年,范围从更好的母性照顾和母亲与婴儿的结合(Kendrick, 2000)到婴儿与母亲的结合(Nelson & Panksepp, 1996)。这些研究路线直接导致了大量的人类研究工作。
目前,关于鼻内催产素在人类中的作用的有趣发现继续以不断增长的速度出现,我们于2010年8月完成的文本将不会反映所有最新的活动。由于所有这些关注,在大众想象中,催产素几乎等同于”爱的分子”:当我们在网上搜索”催产素 爱”时,有205,000个搜索结果,其中大多数是轻量级的炒作或营销,尽管支持这些推测的科学研究一直在增长。但直到今天,几乎没有令人信服的证据表明催产素能像许多许多成瘾性分子那样强有力地提升积极情绪。如果它是爱的中介者,难道不应该吗?没有确凿的证据表明它对动物具有显著的奖励作用。事实上,如果发现它在某些条件下能持续促进积极情绪,那么人们甚至可以推测这种效应可能是由于催产素促进了大脑中的阿片类物质活性(Kovács et al., 1998),这将符合一个更有支持的理论,即社会依恋以及由此延伸的陪伴之爱是一个由大脑阿片类物质介导的过程(Panksepp, 1981a, 1998a)。
尽管如此,在许多实验中,催产素确实促进了动物和人类的各种亲社会行为和态度。在人类中,它增加了在经济交换中信任他人的意愿(参见 Meyer-Lindenberg, 2008)。当伴侣讨论话题并存在意见分歧时,积极互动(眼神接触、兴趣、情感自我表露、认可、关心、非语言积极行为)与消极互动(批评、蔑视、防御、支配行为、好斗、冷战、非语言消极行为、打断)的比率显著上升(Ditzen et al., 2009),等等(Heinrichs & Domes, 2008)。换句话说,在适当的条件下(与你已经爱的人在一起),催产素使我们更加亲社会——更宽容、更友好。然而,我们应该记住,催产素系统都是深度皮层下的,在大脑中相当低级和古老,所以显然它的原始功能不是控制浪漫爱情和计算他人可信度这样的高级认知活动。因此,这些影响首先必须通过某种类型的初级加工大脑机制的变化来解释。所以一个关键问题是,催产素给药后亲社会友好感受是否上升,还是只是减少了有时在人际交往中出现的压力性焦虑感受?在我们的研究中,所见过的最强和最可重复的临床前效应之一是减少分离痛苦(Panksepp, 1992)。参见图1.8的样本(这构成了我们在这个指标上收集的数据的约5%)。从这个角度来看,我们可以预期,如果没有正常的催产素分泌,母亲容易患产后抑郁症,当我们校对本书时,最近的一篇论文恰好提出了这样的关系(Skrundz, et al., 2011)。
最近,出现了一系列研究,甚至质疑更为理性的”亲社会”结论。例如,在经济游戏中,可以对虚拟(基于计算机的)对手输赢,如果竞争对手碰巧输了,催产素会增加幸灾乐祸。如果虚拟对手得到的分数比你多,它会增加嫉妒(Shamay-Tsoory et al., 2009)。这不是很亲社会。因此,这种促进母性行为的肽有其棘手的一面。当其他人在可以表现利他主义的环境中测试人们时,感受是复杂的。它确实倾向于促进对你的内群体——你的朋友——的合作情感,但它对外群体陌生人的作用相反,它增加了防御性攻击(De Dreu et al., 2010)。这些不是人们从亲社会爱分子那里期望得到的效果。
那么问题出在哪里?大脑中实际的情感变化是什么,能导致如此多样的影响?催产素的递质通路相当有限,没有迹象表明它如何能在三级加工水平上直接产生这种变化。是否仍然存在一种单一类型的初级加工情感转变,可以解释这些以及其他令人困惑的人类结果?也许可以,但没有人提出令人信服的建议。再举一个悖论:催产素给予那些患有边缘型人格障碍(BPD)的人时,会降低信任和合作反应的可能性(Bartz et al., 2010)。我们想提出一个解决方案,可以将这些不同的结果结合在一起,基于我们在鸟类中观察到的催产素效应。
在鹌鹑物种中,陌生的雄性彼此特别不能容忍。它们啄对方的头,直到一方放弃并简单地服从另一方的啄击。通过这种方式,鹌鹑永远知道它们在啄食顺序中的位置,只要占主导地位的动物仍然茁壮成长。所以,我们想知道,如果严格按照初级加工操作的非常年幼的鸟类,直接在它们的大脑中(进入它们的脑脊液,这在动物研究中很常见)接受催产素,会发生什么。我们测试了刚孵化的家禽雏鸡,当它们与其鸟群的安全感分离时,明显的效果是它们几乎不哭(图1.8)。此外,它们还表现出更多的打哈欠。它们摇头更多,表现出更多的拍翅(Panksepp, 1992)。如果在群体中测试,注射催产素的动物比单独测试时表现出更多的拍翅,这似乎表明它们”感觉精力充沛”——它们通常更自信(打哈欠和摇头没有受到社会促进)。
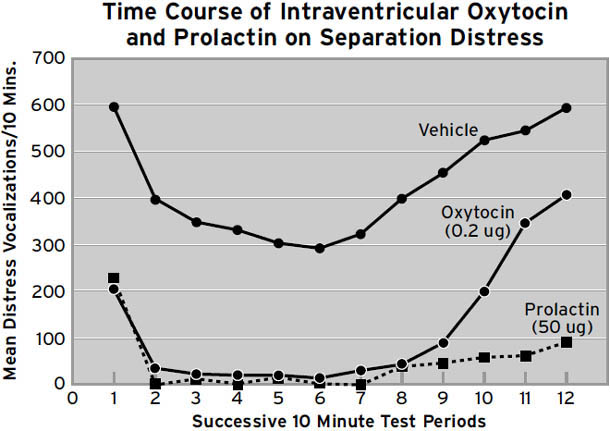
图1.8. 脑室内催产素和催乳素对5至6日龄雏鸡分离痛苦叫声的影响,这些雏鸡与其鸟群社会隔离2小时。这些对哭泣的显著影响是在没有任何明显镇静作用的情况下产生的,就像刺激μ受体的低剂量阿片类药物一样。
我们想知道,当一只鹌鹑获得类似催产素的提升时,社会支配地位会发生什么变化。令我们惊讶的是,获得这种提升的鹌鹑真的被其他鸟类啄了头。也许它们变得更顺从了,但另一种可能是,它们只是对其他鸟类的”不良行为”变得更加宽容。如果正常鹌鹑的头被啄这么多次,它随后会变得极度顺从。但当我们第二天再次测试相同的配对时,那只看起来如此顺从的鹌鹑现在像强劲反击一样回来了,并成为了胜利者(Riters and Panksepp, 1997)。这种”逆转”在正常鹌鹑中几乎从不发生。一旦你成为失败者,你就会保持那个位置。那么为什么在最初那天,当一只鹌鹑正在失败时,类似催产素的提升会让这只”顺从”的鸟在第二天成为胜利者呢?这只鸟只是忘记了吗?如果是这样的话,最多也应该是势均力敌。事实上,那只输了的鹌鹑回来后更强了,这表明它前一天只是平和而已,但仍然感觉相当强壮。我们能说”自信”吗?这当然似乎是一个合理的假设。
那么如何在动物身上测试这样一个微妙的情感构念(construct)呢?把一群幼鸟(在我们做的未发表研究中是家养小鸡)放在一个大房间里的桶下,看看它们会移动多远来探索新房间如何?事实上,正常的幼鸡往往会聚在一起,并作为一个紧密团结的群体移动。但当我们将催产素注入它们的大脑时,它们分散得更松散,好像它们不那么亲社会,或者说,它们更”自信”,焦虑减少了。我们从1980年代中期就已经知道,这种分子在减少鸟类的分离焦虑方面非常强大([图1.8])。
也许催产素可以增加动物的自信。这能解释人类研究吗?似乎合理的是,当你自信时,你在经济交易中会更有安全感。当与配偶讨论冲突观点时,你难道不会更友好和宽容——更少防御性吗?如果一个陌生人在赌注中赢的钱比你少,你感觉更自信时难道不会更愿意幸灾乐祸吗?如果你赢得更少,你难道不会更愿意在你心智的更高层面承认你有点嫉妒,并更愿意表达出来吗?如果你有边缘型人格障碍(BPD)的慢性不安全感,提升自信的催产素剂量难道不会合理地将你转向独立感,你可能更愿意主张自己的观点,而不是保持在慢性依赖需求的控制中吗?因此,我们需要的是一个良好的自信心理测试。与此同时,已经有一些适度的数据表明催产素可以减少社交焦虑障碍(Guastella et al., 2009)。
因此,通过对情感焦点的小小转移,转向社会生活的一个非常基本的方面,突然之间,人类研究中令人困惑的发现多样性开始变得有意义了。当然,自信对于称职的母性来说是一个非常重要的特质——当个人责任急剧增加时,能做到的态度对母亲很有帮助。如果这样的解释是可能的,这里有一个预测,基于一个众所周知的事实,即很多人害怕公开演讲。如果催产素增加社会自信,那么表演焦虑应该会减少。通过研究动物大脑中的初级加工系统和催产素回路,我们可以对人类可能预期的心理变化做出一些显著的预测。例如,催产素应该增加我们探索另一个人眼睛的倾向,试图读懂他们的想法,因为你感觉更安全。事实上,它确实如此,特别是如果你抑郁的话,尽管单次鼻内剂量的催产素并不能显著改善抑郁症状(Pincus et al., 2010)。催产素甚至减轻了一些精神分裂症的症状(Rubin et al., 2010)。这些分子的进化跨度是巨大的。即使是鱼类,当这些分子被注入它们的大脑时,也会表现出各种社会效应,包括促进一夫一妻制交配行为(Oldfield & Hofmann, 2011)。
那么催产素是爱的分子还是减少焦虑和促进自信的分子?聪明的选择应该是后者。也许如果你在爱情中充满自信,你会有更多的性生活和更多的孩子。
当我们思考进化时,我们通常指的是动物物种在漫长岁月中身体变化和发展的方式。然而,当我们谈论情感时,我们通常指的是心智,它通常被认为是一个非物质的实体,这是无稽之谈。心智只是意味着某些大脑状态有一种主观感受,这服务于某种适应性功能,例如为更高级的意志行为提供”行动中的意图”基础,即”行动的意图”([图1.4])。因此,如果我们理解情感是物理大脑的功能,那么将初级加工情感作为进化现象来讨论就是有意义的。它们在如此多物种中的相似性表明,情感能力是大脑的古老功能。像许多适应性进化发展一样,支持生物学上成功的情感能力的大脑系统在动物进化过程中被保留了下来。其他被保留的进化发展包括DNA复制、代谢功能如消化和呼吸,以及细胞能量的产生。如果你理解克雷布斯循环(Krebs cycle)在一种动物中如何工作,你就对它在所有动物中如何工作有了很好的理解。
进化逻辑表明,情感能力在各个物种通过自然选择出现时得以保留,是因为这些大脑功能提供了有效的生存和繁殖方式。这些大脑功能提供了选择优势,因为它们有效地预测了普遍的、未来的生存需求。拥有这些能力的动物以更大的成功率存活和繁殖。从这个角度来看,情感是大脑内置的预期性神经心理机制。想象一下疼痛对你的生存有多么有用。
情感为生存提供了灵活的指导。在情绪进化之前,动物的行为方式必定更加刻板。例如,原始海洋生物别无选择,只能以统一的动作起伏前行。相对僵化的行为也可能相当复杂——蜜蜂执行着多种本能功能,其中一些我们认为可能具有情感维度。例如,当实验者将蜜蜂从高浓度的甜糖溶液转换到浓度低得多的糖溶液时,蜜蜂确实会表现出类似挫折的反应,这种低浓度糖溶液可能吸引力小得多(Wiegmann et al., 2003)。然而,哺乳动物完全成熟的情感能力使动物能够以高度灵活的方式应对当下的生活挑战。
例如,如果一只老鼠习惯于在田野的某个角落觅食,而一只雪貂在附近居住,即使雪貂不在场,老鼠也会闻到雪貂的气味。这种捕食者的气味无条件地激活了老鼠的恐惧系统。这种激活触发恐惧情感,感觉很糟糕。老鼠为了避免感到恐惧而躲避雪貂的气味。为了避免感到恐惧,老鼠会找到另一个觅食地。通过这种方式,情感使动物能够预测事件。但请注意,这种预期不是一种认知功能。它是一种自发的情感反应,导致无意识的学习机制被激活,使动物能够避免恐惧的感觉。
虽然老鼠的行为可能向我们暗示老鼠在某种程度上意识到可能在哪里找到雪貂,但事实并非一定如此。单独的恐惧就是预测未来事件的可靠方式,即使老鼠有限的认知能力无法将关于未来的想法概念化(这显然是脑心的三级方面)。由恐惧系统产生的防御性情感保护个体的生存,而关怀(CARE)的养育情感保护他人的生存(特别是那些携带部分关怀个体基因的其他个体)。性欲(LUST)同样保护物种的生存。关键是,先天的情感能力以增强当下和跨代生存的方式指导动物行为。
虽然情感系统位于皮层下大脑的深处,但另一方面,认知从新皮层中产生,新皮层是大脑的最外层,也是进化上最新的部分。这表明情感体验的能力在允许动物应对复杂环境情况的复杂认知能力之前很久就已进化。同样值得注意的是,情感系统在大脑内更深的进化位置使它们更不容易受到损伤,这也可能突出了一个事实,即它们是比认知系统更古老的生存功能。
我们说情感是初级过程体验,因为它们是纯粹的心理元素,不同于其他任何东西。但我们也可能有理由将情感视为意识的原始形式——情感可能是大脑中进化的第一批感受体验的来源。但它们有几个变种——情绪性的、稳态性的和感觉性的([图1.4])。原始情感可能是无知觉意识(anoetic consciousness)的原始来源——没有理解的初级过程体验。
总而言之,我们设想的脑心进化中的分层方式促使人们首先关注最古老的层次,并利用这些知识来阐明二级过程,在那里原始情绪功能与感知相整合,允许条件学习。例如,一只开始害怕猫铃铛声音的老鼠正在使用二级情绪过程,初级认知策略也是如此,例如老鼠学会在听到猫的铃铛时跑到它隐蔽的家中。这为动物提供了关于世界的事实性知识——一种原始的知识性(noetic)或知晓形式的意识。但老鼠也会思考这种意识吗?它们是否”意识到”自己正在经历某事。我们根本不知道。也没有人提出解决这一困境的方法。
当前两个层次的心智开始产生更复杂的认知能力时,我们将高级情绪功能称为三级处理层次,例如为周末远足做准备或规划未来职业目标的计划。三级处理允许对世界和自我进行智能反思,同时考虑过去和未来的框架——在自我意识(autonoetic)意识中。这种水平的心理活动在动物研究中非常困难。三级层次与额叶皮层和顶叶皮层的功能密切相关——这是新皮层中最近进化的区域,在人类和其他一些大脑发达的生物中大量存在。
我们所有人都想理解自己内心深处发生了什么,也想理解我们认识的人内心的想法,包括野生动物的思想,以及为我们的生活带来如此丰富色彩的各种驯养和伴侣动物的思想。情感神经科学(Affective neuroscience)为情感性心智-脑功能的本质提供了一个新的、独特的、基于证据的视角,为我们打开了一扇了解我们最深层情感价值观的祖先来源的窗口。
在下一章中,我们将探讨一些科学和历史原因,解释为什么情感作为神经科学研究的主题被边缘化。我们还将简要概述支持其他动物存在情感的研究。当我们讨论SEEKING系统时,我们将更全面地研究同样的研究,该系统为这个问题提供了决定性的证据。我们将为七个初级过程情感系统分别撰写章节。由于学习领域的大量研究都集中在FEAR系统上,我们将在该章节后暂停,总结大脑的一些学习(次级过程)机制。特别是,我们将展示条件反射(conditioning)——有些人认为它是一种认知功能——实际上根本不是这样。它是一种自动的大脑反应,不需要任何新皮层的参与就能成功。与心智的初级过程不同,这种脑心智整合水平似乎是深度无意识的,但为我们提供了认知意识(noetic consciousness)的基础。我们还将强调情感本能——无条件情感网络——在”打开学习之门”方面可能发挥的关键作用(这个话题在很大程度上被那些研究动物学习的大脑机制,特别是恐惧条件反射的人所忽视)。
我们将始终回到关注复杂三级过程和带有情感色彩的思想的人类临床问题,以及情绪调节和失调。正是在这个领域,人类研究是必不可少的,目前各种现代心理治疗学派正在推进许多研究和发展方向,这些学派越来越强调情感问题(见第12章,Panksepp从情感神经科学的角度阐述了他对心理治疗未来的一些看法)。在此过程中,我们还将反思”自我”的本质,以及这些大脑系统鼓励我们考虑的对生命的新敬畏的可能性。
总的来说,我们的观点是,理解情感对于理解人性至关重要。我们的人格结构不仅植根于情感(Davis et al., 2003; Davis & Panksepp, 2011),而且大量与社会相关的人类问题需要从情感和认知的角度来处理。有洞察力的现代心理治疗师长期以来就知道,心理治疗的目标是情感调节。尽管心理治疗可能看起来专注于思想,因为患者主要通过语言交流,但治疗的目的是积极改变患者的情感体验。这不可避免地需要改变他或她的思维方式,但心理治疗的目的不仅仅是改变认知风格或内容。相比之下,许多精神科药物直接改变情感,无需认知干预,但往往在情感得到更好调节后,会出现显著的认知变化。事实上,越来越明显的是,对精神问题的环境、人际和药物治疗方法一起使用比单独使用这些方法中的任何一种都更有效。在本书的最后,Panksepp将讨论替代疗法在更直接地处理情感方面可能采取的一些方向。
最终,情感是我们心理存在的基础。当情感令人满意时,生活就是一种快乐。当情感受到干扰时,生活可能是地狱。正如生活在苏格兰比特岛的诗人约翰·斯特林(John Sterling,1806-1844)所说:“情感回转到自身,而不引向思想或行动,就是疯狂的元素。”在第11章中,我们将为以下结论提供论据:原始情感感受位于心理机制的原始基础——它们是核心自我(core-SELF)的原始生物基质——也许是”灵魂”概念的神经基础。
现在有推论性证据表明,一种对有机体连贯性至关重要的普遍核心自我类型结构,存在于大脑古老区域的深处,那里存在着初级过程情感系统。我们大脑中多样的、进化”赋予”的情感工具可能都依赖于这个广泛的基质来进行原始身体表征,以产生所有哺乳动物体验到的多种类型的原始情感感受,其中有许多细微的进化差异,我们目前对此知之甚少。
相比之下,我们的许多高级情感观点——从指责到羞耻,从嫉妒到同理心和善良——与我们的认知机制紧密交织。我们的高级认知机制为我们提供了大量的情感选择,包括同时让自己远离主导情感,沉浸在接纳或”正念(mindfulness)“中。
认知科学仍然几乎完全依赖于心智的计算理论(computational theory of mind),一旦学者们意识到情感性感受(affective feelings)对人类思维的深刻影响,这一理论可能会被彻底颠覆(Davies, 2011)。关于情绪如何支配我们习得的观点以及我们经验的再加工,最终的图景可能与我们目前的暂时性设想大相径庭(参见[第6章])。随着对情感的更好理解,治疗事业有望转向一种更精细的、基于神经科学的视角,了解一个人如何帮助另一个人走向情绪平衡,并协同使用心理疗法和精神药物。
对原始激情(primal passions)的理解可能使人们更容易追求亚里士多德的实践智慧(phronesis)(见[第4章]题词)——即懂得如何以智慧的方式在认知上处理自己的情绪,而不是成为无助的受害者,生活在永恒的冲突中,被我们心智的祖先力量无情地掌控。我们应当认识到,这些力量与指引许多其他动物生活的力量是相同的。我们最终理解自己更深层次精神本质的方式,就是理解动物更深层次的神经本质。我们还在等什么?让对话开始吧。
我们无法绝对确定其他人类有体验,更不用说非人类动物有体验(“他心问题”)。但基于进化论,似乎可以合理地假设,意识的形式随着体现它们的生物形式一起进化。但蜜蜂看到的是什么?……飞蛾或海豚听到的又是什么?
——Max Velmans (2009, 第192页)
Max Velmans的评论突出了我们的困境。原始体验——哲学家所说的现象意识(phenomenal consciousness)——是如何从大脑活动中产生的?这不仅是意识研究的”难题”(hard problem),也是整个神经科学的难题。事实上,解码其他动物的大脑如何体验感觉输入,可能比解码基本情绪感受的情感性质要困难得多。为什么?(i)因为我们可以通过刺激动物大脑的特定区域来唤起明确的情绪行为模式,(ii)因为每一种如此唤起的初级过程情绪都伴随着消极或积极的情感状态,这些状态可以通过各种学习任务客观地监测,而不需要语言性的自我报告。因此,我们确定神经回路如何在大脑内产生情绪”奖励”和”惩罚”,比确定知觉要容易得多。我们可以确定的是,动物对任何形式的人工诱发情绪唤起都不是中立的。通过我们可用的各种学习和偏好测量方法,我们知道所有被研究的哺乳动物都不喜欢某些类型的大脑唤起(愤怒、恐惧和恐慌/悲伤),而喜欢其他类型(寻求、性欲、照料和玩耍)。
然而,必须强调的是,这些积极情绪在某种程度上都分享寻求冲动(消极情绪的唤起也可能分享这一点,如在恐惧中寻求安全,在悲伤期间寻求母性照料)。这些情感性评估能力与所有其他被研究的哺乳动物共享——这一点我们有科学的信心。许多这样的大脑回路存在于其他脊椎动物中。相关的大脑化学物质甚至可能在一些无脊椎动物中介导情感:某些物种(如小龙虾)对俘获人类的成瘾药物表现出明显的偏好,如吗啡和安非他明(Huber et al., 2011)。
关于其他哺乳动物各种初级过程积极和消极情绪的体验性质,我们能得出更多结论吗?也许可以。这些各种感受状态的内在动力学可能与相应本能的外在情绪表达有着密切的相似之处。每一种感受到的情绪都通过可见的迹象在行为上表达出来,这在”低等”动物中特别明确——从寻求到悲伤的各种表现。成年人可以轻易抑制他们的情绪表现,可以说让他们的感受”潜入地下”(事实上,新皮层在这种原始情绪被调节——被控制住时功能最佳)。然而,在我们的孩子身上,这种身体动力学仍然传达着我们最强烈情绪唤起形式的整体性质。想想愤怒的猛烈坚持、恐惧的颤抖、玩耍的轻快活泼、慈爱照料的温柔抚摸,以及我们将比其他任何情绪都更关注的——寻求的热切搜索和四处探寻。这些也是可以通过刺激大脑特定区域唤起的行为类型。这些自然的情绪表达可能与情绪感受本身有着密切的相似之处。这是一个关键点:情绪感受及其自发的行为表达源于相同的古老神经系统。因此,我们现在知道在哪里寻找情绪感受的构成和神经机制。
但我们如何知道各种负面和正面的感受实际上是不同的,而不是一种原始的好感受和一种坏感受的微小变体呢?在积极情感中,可以确定动物是否能区分由各种神经化学物质(例如,神经肽和精神药物)或各种奖赏和惩罚形式的直接脑刺激所引起的不同情绪状态。事实上,我们确实知道动物能够区分大脑中某些不同”奖赏”部位的积极感受(Stutz et al., 1974),以及成瘾性阿片类药物如吗啡和精神兴奋剂如可卡因所产生的内部状态(Overton, 1991),所有这些对所有哺乳动物都具有高度奖赏性(Tzschentke, 2007)。但在我们了解不同原始情感(primal affects)的实际数量以及产生不同情绪感受的大脑机制之前,还需要进行更多这方面的研究。
处于基本情绪状态的动物也会发出特征性的声音,这些声音通常与我们发出的情绪声音没有太大区别。想想疼痛的尖叫、愤怒的咆哮、笑声的重复鸣叫声。这些声音来自灵长类动物大脑中不同的网络(Jürgens, 2002)。同时,每种类型的声音基本上都来自所有被研究过的哺乳动物物种的相同大脑区域(总结见Brudzynski, 2007; Brudzynski et al., 2010; Newman, 1988)。因此,产生情绪情感的皮层下脑系统在整个哺乳动物王国中都非常相似。也有大量证据表明,人类的基本情绪感受来自这些相同的低级脑系统,而不是来自新皮层的高级区域(Damasio et al., 2000; Northoff et al., 2009; Vytal & Hamann, 2010)。
因此,动物的初级过程情绪感受与我们自己的相似的可能性不仅基于大量数据,还基于我们初级过程情绪本质在跨物种进化上的显著连续性(Darwin, 1872/1998; Panksepp, 1998a)。这种相似性还在大脑的基本情绪学习机制中得到了显著证明(LeDoux, 1996)。我们不能轻易地将这些概念推广到第三级过程的心理复杂性水平。其他动物似乎不太可能体验到敬畏或崇高的感受,缺乏可靠的证据将使我们无法考虑这种可能性。尽管黑猩猩在争吵后确实表现出和解行为(de Waal, 2009),但它们可能不会像我们那样体验到宽恕的优雅。高阶感受用现有程序根本无法研究。因此,没有实验证据表明其他动物会思考幸福的意义,或有足够的自我反思来感受尴尬、内疚和羞耻的刺痛。也许当它们被某人虐待时会怀恨在心(想想大象在反复被人类虐待后暴走的故事)。但我们无法像窥探它们的情绪那样有效地窥探它们的思想。关于对人类事务具有重要意义的微妙的第三级过程情绪—从贪婪到同情—的问题,可能永远无法在其他动物身上进行神经科学细节研究。尽管可以从仔细的行为观察中推断出一些可能性(Bekoff, 2007; Grandin & Johnson, 2009),但没有科学可靠的模型来研究其他动物的这种复杂的第三级过程情绪。然而,原始的情绪感受终于可以进行实验研究了,这些知识可能对理解我们自己更深层的本质以及我们与其他动物的亲缘关系具有深远的意义。
因此,与初级过程情绪相反,初级过程情绪在大脑中有专门的(进化的)神经控制,而动物的大多数高阶情绪(同理心、幽默、嫉妒、羞耻等)的行为指标必然仍然模糊且有争议,即使可以系统地收集人类的意见(Morris et al., 2008)。在科学层面上,它们的存在目前是基于轶事证据。当然,轶事的复数可能就是数据,至少对于那些愿意相信许多其他动物确实有高级情绪的人来说是这样(Bekoff, 2007)。而且有大量行为证据表明,许多高等灵长类动物表现出复杂的社会情绪(de Waal, 2009)。甚至老鼠也表现出可能表明同理心的行为和自主神经变化(例如,恐惧性僵住和心率变化)(Chen, et al., 2009)。
这些微妙的、高阶的情绪过程当然可以通过人类脑成像技术来研究(Decety & Ickes, 2009; Iacoboni, 2009a, 2009b)。当这些研究工具变得足够精细,能够常规化地可视化我们在本书中讨论的皮质下情绪网络中发生的变化[1](例如,通过使用更强大的磁场和更高度复杂的统计技术),我们可能会发现,所有高级人类情绪的情感力量——那些奇妙而微妙的感受——仍然根植于古老的神经区域,哺乳动物的原始过程情感正是从这里产生的。原始过程可能仍然是这种新兴多样性的坚实进化平台。事实上,越来越多的证据支持这一点。在脑扫描仪中,情感变化与皮质下唤醒的正相关性远高于与新皮质唤醒的相关性;皮质唤醒往往反映出感受强度的降低。因此,当我们的大脑充满情绪感受时,皮质唤醒通常处于低点,而当感受强度低时,它则处于高点。这表明高级脑活动倾向于抑制来自低级脑区的感受(Northoff et al., 2009),正如去皮质动物的过度情绪化长期以来所显示的那样。然而,也有研究表明,当人类在脑扫描仪中反复思考他们的情绪时,他们这种自我关注的沉思通常会唤醒大脑的内侧额叶区域(Northoff et al., 2011)。像”正念”这样的冥想策略——学习如何在当下时刻保持平静——在减少这种反复性情绪唤醒方面,可能比传统的心理治疗方法更有效(Siegel, 2007)。
仍有可能只有人类和相关灵长类动物(也许还有大象、鲸鱼和海豚),通过它们丰富而复杂的家庭生活和延长的早期认知发展,才能体验到比大多数其他动物更复杂的社会情绪。但这些仍然是未经研究的问题,也许超出了当前科学审查的范围。在人类中,基本情绪和复杂认知的融合必然会对一个人的情绪生活产生深远影响,有时是好的,但往往更多是坏的。我们似乎比其他动物更容易受到情绪障碍的影响,因为我们有能力通过高级认知过程的力量让情绪持续发酵。当人们纠结和反复思考他们的烦恼时,这可能会维持和激起独特的情绪动荡。然而,我们在这里不会过多讨论这些高级人类情绪。我们的任务是为将其他动物纳入体验原始过程情感的群体中建立有力的论证,我们将努力客观地、神经科学地做到这一点。
我们曾说过,理解人类和其他动物的原始过程情感,对于理解心智-大脑如何运作至关重要。我们相信,如果我们要破解意识的神经密码,并为精神病学上重要的生活问题带来新的更好的治疗方法,这是一个关键的研究领域。神经、行为和心理分析之间的科学三角验证,跨物种科学研究现在允许进行,最终提供了对共享的动物和人类情绪比以往任何时候都更深刻的理解。但要做好这一点,我们还必须审视一些历史原因,即为什么心理科学和神经科学倾向于边缘化对动物心理生活的研究,特别是对动物情感生活的研究,至少直到最近(Panksepp, 1998a)。然后,我们将总结神经科学证据,证明原始情感——我们心智的祖先感受——如何从我们与这个世界上如此多其他生物共享的皮质下系统中产生。在我们继续讨论后续章节中每个原始过程情绪系统之前,我们在这里分享情绪研究的历史,特别是情绪感受的研究,以便从透视角度看待仍然影响该领域的各种交叉潮流。对于那些不想反思这些历史力量的人,请随意移步下一章,讨论SEEKING(寻求)系统。
直到最近,许多哲学家甚至一些科学家倾向于将心理生活视为非物质的和附带现象——作为硬生物科学永远无法解决的话题。神经科学,像其他硬科学一样,必须依赖于生理和行为事实的客观观察,许多同事仍然认为动物体验(原始意识)无法被测量。它无法被称重。它没有长度或宽度;它只由模糊的神经动力学深度组成,即使在人类中也无法以任何方式严格监测,而在人类中,语言反馈可能是特殊的和具有欺骗性的。只需考虑影响右半球的中风患者的虚构,这使得他们的说话半球失去了深层的情感指导。这些人往往用他们自私自利且具有语言能力的左半球生成的幻想故事来否认他们明显的左侧瘫痪——这些虚构有时会在精神分析疗程中消失(Kaplan-Solms & Solms, 2000)。例如,这些人可能会长篇大论地说得好像他们没有任何障碍,但当他们放下社交面具,自由地谈论这些残疾对他们”毁灭”的生活的意义时,突然承认他们的虚弱和恐惧。
许多神经生物学导向的科学家坚持认为,我们无法对心理生活做出任何深刻实质性的陈述,当然也不能对其他动物的心理生活做出陈述,我们甚至不能断言意识是真实的——它不过是我们想象的产物。1992年,著名进化生物学家乔治·克里斯托弗·威廉姆斯写道:“我倾向于将它(心理领域)从生物学解释中删除,因为它是一种完全私人的现象,而生物学必须处理可公开证明的事物。”许多同事都同意这一观点。我们不同意。如果我们不处理痛苦中的人们的真实感受,不尝试科学地理解他们深刻的、往往是消极的感受,我们就永远无法真正理解他们在情感上的困扰。这种理解的很大一部分必须来自对我们的动物同伴的研究。我们可以设想这样一天,像抑郁症这样的心理疾病可以通过运用我们对积极情感的知识来治疗,以重新平衡被消极情感压倒的心灵。当然,这也需要协同的人际互动,特别是当我们发展新的、更有效的心理治疗实践时(例如,参见本书最后几章)。总之,我们的主张是,如果没有对动物大脑和心智进行适当的、理论指导的研究,就无法获得对情感的生物学理解。这可能会让许多人感到惊讶。但如果仔细思考所有相关的科学和伦理问题,情况必然如此。
无论如何,当我们研究脑心(BrainMind)时,我们面对的不仅仅是大脑回路和分子,还有从这些神经物理基质中产生的复杂感受质地如何帮助创造心理生活。要理解各种精神障碍,我们必须科学地面对情感体验的本质。我们不能从”抑郁症”的诊断标签直接到达对这种神经-心理现象的完全基于大脑的理解,除非我们问:“为什么抑郁症会痛苦?”以及更准确地说:“这是什么样的痛苦?”
“从大脑中删除心智”的倾向有着悠久的历史——它有两个主要分支。一个分支是二元论(dualism),这是一种相信存在两个本体论领域的信念:非物质的与物质的并存。二元论是古希腊人思想的组成部分。在过去四个世纪中,它最著名的支持者是哲学家勒内·笛卡尔(1596-1650),他在最近之前有许多追随者。历史的第二个相关分支源于一场科学运动,这场运动兴起于十九世纪后期一群致力于现代化医学课程的革命性德国医生群体,那时科学家对神经系统还知之甚少。让我们逐一审视这些论点。
在笛卡尔的著作之前,二元论已经被许多学者接受了几个世纪。它是古希腊人思想的组成部分,他们通常认为非物质现实比物质世界更重要。柏拉图(公元前424-348年)认为”形式”(forms)——非物质的概念性现实——捕捉了物质现实的真正本质。例如,人们可以在单个物体中看到美,但要理解美的本质,就必须将美理解为一种”形式”,一种存在于所有个别美的实例之上和之外的概念。因此,对柏拉图来说,物理现实只是终极非物质现实的反映:理想形式的现实。
亚里士多德(公元前384-322年),古代伟大的生物学家,提出所有生物都赋有灵魂(soul),他将其视为不是个人的灵魂,而是一种解释物理世界变化的非物质自然力量。例如,幼苗的灵魂将解释其成长为树的潜力。现在我们认识到这些原因来自遗传。圣奥古斯丁(354-430),基督教最有影响力的早期思想家之一,相应地将灵魂描述为一种特殊的物质,赋有理性,有助于统治身体。笛卡尔给奥古斯丁传播的亚里士多德思想赋予了特别宗教化的诠释,可能至少部分是出于政治原因(他不希望像伽利略那样被教会审查,酷刑的威胁使这一点具有深刻意义)。他将非物质力量理解为个人意识,他将其描述为上帝精神在人类心智中的表达。通过这种方式,上帝的非物质精神决定了人的行为。
笛卡尔以不同的眼光看待动物。他不认为它们是有意识的生物,因为他相信上帝不会在如此低等的生命形式中显现他的神圣精神。他将动物视为不过是活的机器,没有神圣火花的生物。这种观点导致了对动物的不人道实验(例如,没有麻醉的活体解剖);它们的抗议声音和逃跑努力被视为不过是缺乏任何意识体验的反射反应。只有人是有意识的存在,人的意识是上帝神圣领域的一部分。因此,人的意识决定了他的行为。为了使这个牵强的想法奏效,亚里士多德的灵魂和笛卡尔的人类神圣心智必须被视为受非物质力量控制,这些力量也决定了物理世界的行为。亚里士多德的理论解释了所有生物的变化,笛卡尔的理论特别解释了人类的行为。
不可物质的存在领域这一概念也出现在古代希波克拉底医学中,该医学信奉活力论(vitalism)。活力论遵循亚里士多德关于不可物质力量导致物质世界变化的信念。根据希波克拉底的观点,生命力量创造了疾病和健康(Smith, 1979)。希波克拉底(约公元前460年-约公元前370年)被称为医学之父,因为他将疾病归因于身体状态,而不是归因于神秘力量。然而,尽管他拒绝完全以神秘主义作为医学基础,但他仍然是一个二元论者;他坚信不可物质的活力论力量的存在。他认为四种基本的身体体液(humors)或液体(黄胆汁、黑胆汁、黏液和血液)是决定健康或疾病的生命力量的物理表现。在他看来,这些体液的稳定平衡会带来健康,而所有疾病都源于失衡状态。在中世纪,这种思维方式被扩展到情绪气质,出现了胆汁质(易怒)、抑郁质(悲伤)、黏液质(冷漠和恐惧)以及多血质(快乐)性格的概念。
前现代欧洲时代的医学干预主要是为了重新平衡体液,这反过来意味着控制身体和心灵的不可物质生命力量被带入平衡状态(Smith, 1979)。例如,人们认为葡萄酒可以通过促进血液和多血质水平来抵消过多的黄胆汁。柑橘类水果被认为可以减少黏液,等等。在最糟糕的情况下,希波克拉底原则促使医生给患者放血或施用藜芦等毒药,引起呕吐和腹泻。除了可能的有益安慰剂效应外,这些干预措施很可能经常伤害患者或根本不起作用。
在希波克拉底时代,医学科学是一种粗糙的工具,特别是因为国家禁止尸检。希波克拉底对身体的内部运作知之甚少。尽管如此,希波克拉底原则统治医学超过两千年。然而,文艺复兴之后,科学进步开始削弱人们对希波克拉底理论的信心。显微镜等现代发明使科学家了解到某些疾病是由微生物引起的,而不是由体液失衡引起的。但变革总是缓慢且不受欢迎的。直到十九世纪中叶,一群致力于实证医学的欧洲大陆医生(由卡尔·路德维希[1816-1895]、埃米尔·杜布瓦-雷蒙德[1818-1896]、赫尔曼·冯·亥姆霍兹[1821-1894]和恩斯特·冯·布吕克[1819-1892]等知名人士领导—他们都在某种程度上对大脑感兴趣)组成了一个志同道合的医生-科学家团体,后来被称为柏林生物物理学俱乐部(Greenspan & Baars, 2005)。他们拒绝了与现代物理学关于疾病的发现不符的希波克拉底四体液理论。
柏林生物物理学俱乐部也普遍拒绝活力论。他们拒绝所有被假定支配身体功能的神秘力量的存在。这些杰出的科学家坚持认为,非物理力量无法接受科学审查,因此人们无法知道关于它们的主张是否真实,甚至无法知道它们是否真的存在。出于这些原因,柏林生物物理学俱乐部的成员在科学中果断地放弃了二元论。对他们来说,科学必须植根于对物理世界的研究。
这些革命性的医生满足于对物理身体进行实验,并根据他们的观察构建机械论理论。但他们并不将自己的理论视为凌驾一切的不可物质真理。科学理论只是被视为对现有证据的最佳解释。事实比理论更重要。原则上,面对矛盾的证据,理论总是可以被推翻的。
这场革命运动迅速成功地在坚实的科学基础上建立了新的、严格的医学课程。俱乐部的胜利导致了基于证据的医学方法,这仍然是医学教育的基础,直到今天。然而,在心理学中,反活力论采取了一种特殊形式。如何从生物物理学角度研究心灵?由强大的行为主义运动提出的临时性前神经科学解决方案—该运动统治学术界直到二十世纪最后二十五年,并且在神经科学中尚未消亡,特别是在行为神经科学中—是意识并不重要。行为主义者选择只研究大脑功能的外部可观察维度(即行为,以及行为作为”反应”所对应的传入”刺激”)。行为主义者最重要的工具是他们称为无条件刺激(UCS)和无条件反应(UCR)的实体—比如电击和由此产生的僵住行为(见第5章和第6章)—这些工具诱导动物快速展示习得的应对策略。通过这种方式,行为主义者能够绕过大脑的”黑箱”(Skinner, 1938),从而绕过心灵。他们似是而非地将从可观察行为和其他科学数据推断心理力量(任何类型的)等同于被否定的活力论力量概念。因此,他们认为无法以任何科学方式研究心灵本身的实际本质。于是心灵不复存在,至少对于二十世纪科学心理学中的大多数研究者而言是如此,尤其是在动物研究的背景下讨论时。
哲学中的实证主义运动将所有概念的严格定义(实证主义)视为建立坚实科学的唯一途径。伟大的语言哲学家路德维希·维特根斯坦(1922/1981)在其《逻辑哲学论》中,通过其著名论断”当答案无法用语言表达时,问题也同样无法表达”(命题6.5),为对心智研究的无情唯物主义挑战提供了”决定性”的支持声明。由于心理特质不可能用清晰的、可操作化的科学语言来表达,人们面临以下困境:“即使所有可能的科学问题都得到了解答,生活的问题仍然完全未被触及。当然,这时就不再有问题了,而这本身就是答案”(维特根斯坦,1981,命题6.52)。
关于维特根斯坦更多的怀疑论指导,请参见[第13章]结尾。令人感慨的是,在第二次世界大战的人类悲剧之后不久,这位一生大部分时间饱受情感折磨的维特根斯坦,在其《哲学研究》中为心理生活的研究提出了一个更为宽容的愿景——在这个愿景中,我们的相对主义语言游戏占主导地位,导致了心理学中强大的社会建构主义运动,这一运动至今仍然兴盛。
当然,活力论和心智是截然不同的。活力论提出存在一种基本的非物理现实。活力论的力量被设想为没有任何生物学前因或物理基础。相反,它们被认为是决定身体健康的无形力量。另一方面,心智具有明确的生物学前因,毫无疑问是物理大脑的一种属性。它不是一种脱离实体的自然力量。它是一种大脑功能,因此可以像任何其他生物学事实一样,用正常的科学方法来研究。我们所需要做的就是继续进行这项艰难的工作,这正是情感神经科学(Panksepp, 1998a)领域的研究者所寻求做的。由于神经科学的进步,这终于成为一个可行的项目。
不幸的是,柏林生物物理学俱乐部的一些边缘成员未能理解这些区别。生理学家雅克·洛布(1859-1924)在美国工作,先是在布林茅尔学院,然后是芝加哥大学,最终到了洛克菲勒大学(当时称为”研究所”)。在芝加哥大学期间,他影响了约翰·B·华生(1878-1958),后者最终成为”行为主义之父”。在哈佛,B·F·斯金纳(1904-1990)也被洛布的思想所说服。华生和斯金纳在洛布的启发下,共同制定了一种新的、方法论上严格的——并最终成为教条式的——激进行为主义。
他们是许多心理学家的英雄,尽管他们将”心智”从课程中剔除。在某种程度上,他们之所以成功,是因为现代科学的笛卡尔基础,即深刻的怀疑论。让我们回忆一下,笛卡尔通过怀疑一切来开始他的哲学。他很容易想象他周围的世界不过是一场梦或幻觉。他认为怀疑逻辑和数学的现实性没有问题,因为他相信邪恶的恶魔可能在控制他的推理。他唯一不能怀疑的是他在怀疑,这导致了他从无限怀疑中的救赎:唯一无可辩驳的证据——我思故我在。因此,怀疑论成为科学领域的通行货币。“向我证明”成为口号,尽管在二十世纪科学中已经清楚,没有科学证明,只有数学和逻辑证明。由于科学的本质,它必须基于证据的分量。从这个角度来看,本书的主要论断对于那些遵守科学规则的人来说应该是一个好消息:大量事实表明,其他哺乳动物确实具有情感体验,我们都拥有非常相似的初级过程情绪的神经基础。但就这一重大问题而言,对许多神经科学家来说,他们对怀疑论的热爱仍然超过了对证据分量的理性权衡……以至于几乎没有关于这个话题的讨论,至少在行为神经科学家中是如此,而他们拥有将这些问题推向经验解决方案的最佳工具。
因此,行为主义的奠基者们——这些对心理科学中任何心理构念(mental construct)必要性持极端怀疑态度的人——为行为分析带来了新的复杂程度,这为心理学领域提供了此前缺失的严谨性。他们给了我们第一个有前景的方法来分析习得性行为变化的原因——即学习。他们为科学家提供了能在实验室中可靠地产生行为变化的工具。但为了实现这一目标,他们觉得必须拒绝所有对内在情绪和动机过程的引用。华生(Watson, 1929)最初对情绪感兴趣,但他认为智力能力,独立于任何气质问题,是在没有多少先天功能影响的情况下习得的。他著名的声明是:“给我一打健康的婴儿,体格健全,以及我自己指定的世界来抚养他们,我保证随机选一个,把他训练成我可能选择的任何类型的专家——医生、律师、艺术家、商业领袖,是的,甚至是乞丐和小偷,不管他的才能、倾向、趋势、能力、职业和祖先的种族如何。”斯金纳(Skinner)走得更远。他从一开始就鄙视新行为科学中的情绪概念,并著名地宣称:“’情绪’是我们通常归因行为的虚构原因的绝佳例子”(Skinner, 1953)。奇怪的是,这两位科学家都不认为心理学必须参与大脑研究才能成为一门完整的科学,但那是在神经科学成熟为理解生物体行为的最重要科学学科之前很久的事了。
因此,意识体验——特别是情感体验——对这些激进的行为主义者来说毫无意义。他们忽视了达尔文关于动物行为提供其情感状态指示的建议,也忽视了威廉·詹姆斯(William James)的信念,即情绪感受不是在情绪行为之前唤起的,而是随之而来(或与之相同)的表达。从某种意义上说,这就是本书的信息,但它只是认识到是大脑的情绪-行为系统承载情感信息,而不是外周身体的情绪行为。这不是一个小的区别,因为即使达马西奥(Damasio, 1994)也被类似的皮层情绪感受观点所吸引。
所有这类思考,对行为主义者来说,都”只是空谈”。行为主义者也忽视了爱德华·桑代克(Edward Thorndike, 1874-1949)提出的原始的、著名的”效果律(Law of Effect)“的措辞,他是最早系统研究动物学习的心理学家之一。桑代克的原始版本坚持认为动物体验到”满意”和”不适”的感受,这不仅驱使它们表现出偏好和厌恶,而且还指导它们的学习。原始的”效果律”实际上是一个”情感律(law of affect)“。行为主义者拒绝了这一方面。以下是桑代克确切提出的内容:
对同一情境做出的几个反应中,那些伴随或紧随动物满意[强调添加]的反应,在其他条件相同的情况下,将与该情境更牢固地联系起来,因此当它再次出现时,它们更有可能再次出现;那些伴随或紧随动物不适[强调添加]的反应,在其他条件相同的情况下,将削弱它们与该情境的联系,因此当它再次出现时,它们不太可能发生。满意或不适[强调添加]越大,这种联结的加强或削弱就越大。(Thorndike, 1911, p. 244)
行为主义者没有使用像满意和不适这样的主观词汇——这些词暗示着伴随着感受基调(feeling tone)的有动机的心理状态——而是用更客观的术语替代,指的是外部可观察的事件:奖励和惩罚(或在学习的语境中使用时的强化(reinforcements))。他们认为所有行为都是基于奖励和惩罚的心理上无法定义的方面来习得的。他们明确选择忽视大脑中的情感变化赋予奖励和惩罚事件控制行为的力量这一可能性。行为主义者没有对奖励和惩罚通过在大脑内产生体验而起作用这一可能性保持开放态度,而是纯粹从操作术语(operational terms)来定义”强化”——从世界上的物体在一个方向或另一个方向”强化”行为变化的能力来定义。直到今天,我们仍不知道”强化”是否是一种特定的非情感性大脑功能,还是仅仅是一个用来描述我们如何通过系统地操纵控制动物感受的大脑系统来训练动物的词汇。
有一件事是确定的,动物确实会可靠地努力获得奖励并避免惩罚。人类也是如此。人类和动物都出于情感”原因”做这些事情,这正是行为主义者无法接受为科学上可行的,因此也是可信的,他们的偏见一直传承给行为科学家直到今天。很少有人选择质疑这些假设。由于对情感和动机状态(如饥饿和口渴)的引用不被接受,因此不被允许,这些概念从大多数心理学话语的词汇中消失了。第三人称客观语言成为新行为主义领域的通行货币;第一人称主观语言实际上被禁止出现在科学话语中。这适用于对动物和人类的讨论。但现在,值得庆幸的是,在我们这个开明的时代,这项禁令已经解除。或者说真的解除了吗?事实上,在20世纪70年代初的认知革命之后,行为主义偏见在很大程度上被大多数人更隐晦地保留了下来,而且它仍然是许多研究动物行为的人的主流观点。似乎受过教育的公众并不了解这一事实。我们希望本书能改变这种状况,揭露行为主义原教旨主义的这种残余本质:一种只对在特定传统中受过教育的人才有意义的时代错误,本身并没有任何内在意义!它目前仍在阻碍关于动物大脑和人类心智的心理功能,尤其是情感功能的丰富讨论。
有趣的是,没有迹象表明柏林生物物理学俱乐部的成员会反对研究感觉或意识,仅仅因为它们不是容易研究的身体过程。如果病人抱怨疼痛,19世纪的现代医生肯定会认真对待他们的主张,并试图发现疼痛的生理原因。然而,疼痛的体验不仅仅是一个无意识的物理实体。它是一种物理心理状态,一种现象体验。它是主观的,但它是真实的——大脑的一个生理过程。疼痛有原因,也有影响。它帮助我们生存。因此,即使它是主观的,它仍然值得科学考虑,用于诊断人类和动物的身体损伤和疾病。也许最重要的事实是,疼痛不仅由身体功能障碍引起;它也由大脑中的神经活动引起(实际上是产生或构成的)。尽管疼痛定位于特定的身体部位,但这种体验并不包含在它发起和心理上似乎存在的地方,尽管一些哲学家有不同看法。事实上,据我们所知,大脑将疼痛的感觉投射到身体被表征的神经空间上。有时疼痛(例如,神经性疼痛)主要是由于神经组织的内部刺激性。无论如何,疼痛是大脑的一种属性,它不是在大脑之外的身体中体验到的东西。
古人不确定大脑是否是心理事件的基质。柏拉图和希波克拉底认为是,但亚里士多德认为情绪源于心脏。然而,早在柏林生物物理学俱乐部重振医学科学之前,一些研究人员就已经接受了将对物理大脑的研究作为更好理解心智功能的手段。在具有现代观点的伟大历史先驱中,有托马斯·威利斯(Thomas Willis,1621-1675),一位详细解剖大脑的英国医生(如他1664年的《脑解剖学》所述),随后发表了一篇关于大脑病理学的论文,以及另一篇关于医学心理学的论文:《关于野兽灵魂的两篇论述》(Two Discourses Concerning the Soul of Brutes,1672)。威利斯试图描述心理变化如何与大脑功能相关,同时没有放弃身体的经典体液控制情感气质的想法。到19世纪之交,颅相学家弗朗茨·约瑟夫·加尔(Franz Joseph Gall,1758-1828)和他的弟子约翰·加斯帕·斯普尔茨海姆(Johann Gaspar Spurzheim,1776-1832)进行了更精细的大脑解剖,使人们普遍接受了心智从大脑活动中产生的观念——尽管加尔和斯普尔茨海姆将人格与颅骨形态(颅骨上的”隆起”)联系起来的实用方法是失败的。颅骨形状被错误地认为能准确反映潜在大脑区域或”心理器官”的大小,但这个猜想被认识为一个可耻的过度简化还需要一些时间。无论如何,到19世纪中叶,许多神经系统学者已经准备好摒弃二元论,将大脑设想为心智的器官,正如许多医生准备好抛弃医学迷信并使医学科学现代化一样。
尽管柏林经验主义医学圈子的大多数成员并不关注情感问题,但值得注意的是,伊万·巴甫洛夫(俄国人,开创了系统性条件反射研究方法)师从卡尔·路德维希,而西格蒙德·弗洛伊德(维也纳人,心理治疗之父)师从恩斯特·冯·布吕克。巴甫洛夫在研究狗的自主神经反射时从未忽视情感。他认识到情感的力量,特别是在他的实验室被涅瓦河洪水淹没,差点淹死他的狗之后。他的许多狗随后表现出我们现在所说的创伤后应激障碍(PTSD)。当然,弗洛伊德将情感作为其过早抱负的核心(当时脑科学还不够成熟),试图创建一种名为精神分析的科学深层心理学。弗洛伊德最终放弃了脑科学研究,发展出一种基于情感的精神分析元心理学,但他承认它缺乏”科学的硬性标志”(Freud, 1895/1968)。
柏林生物物理学俱乐部的成员可能会接受一种根植于脑科学的情感心智理论。事实上,在十九世纪确实有学者,如查尔斯·达尔文和威廉·詹姆斯(1842-1910),对情感和意识持有相当现代的观点(Darwin, 1872/1998; James, 1892)。这两位伟大思想家都没有享受到现代脑科学的成果。事实上,直到今天,大多数关于情感的心理学研究似乎很少关注底层的初级加工(primary-process)神经细节,而三级加工(tertiary-process)细节目前几乎无法获得,尽管我们可以通过现代脑成像技术估计感兴趣的区域及其相互作用。与精神分析等经典”纯心理学”理论的追随者不同,目前有几个新运动,包括神经精神分析(neuropsychoanalysis)(见 www.npsa.org),它们提供了心理和神经分析的明智结合。然而,很少有人追随沃尔特·赫斯(1881-1973)等先驱的脚步,他是1949年诺贝尔奖获得者。赫斯首次证明,通过电刺激下丘脑的特定区域,可以在猫身上激发完整的初级加工愤怒行为以及相应的自主神经反应(完整总结见Hess, 1957)。
也许赫斯在心理学界追随者寥寥,是因为他避免谈论他所激发的动物的情感感受。像他那个时代的其他人一样,他选择将这种电刺激诱发的愤怒表现称为假性愤怒(sham rage)。在退休后,他承认对自己过于胆怯感到遗憾,没有忠于自己的信念,没有声称他的动物确实感受到了真正的愤怒。他坦白这样做是因为他担心这样的言论会招致强大的美国行为主义者的攻击,他们可能因此也会边缘化他更具体的科学发现。他在最后一本书《心智的生物学》(The Biology of Mind)(1964)中试图在一定程度上纠正他的”错误”,但这部作品影响甚微。尽管如此,他至少提供了可以为心理学提供神经生理学基础的数据,这是威廉·詹姆斯和查尔斯·达尔文都会非常欣赏的。
行为主义统治学术心理学约50年,直到二十世纪后三分之一时期才逐渐失去影响力,当时认知革命(cognitive revolution)重新确立了心智研究的科学合法性。受计算机发展的启发,认知科学家认为心智就像一台活的计算机,使人类和动物能够计算偶然性并做出指导行为的决策。心智的计算理论(Computational Theory of Mind)由此诞生,据推测,这也可以在没有脑研究的情况下被理解。特别令人感兴趣的是无意识或先天认知能力的概念,例如诺姆·乔姆斯基的假说(1968),即人类儿童对所有语言共有的基本语法结构具有先天知识。然而,在大多数情况下,认知科学关注的是信息处理的机制——感知和学习——而不是活体心智的内生性和生成性特性。认知革命主要关注心理活动中最接近计算机软件的那些方面——心智的”信息处理”部分——因此直到最近才开始涉及情感、动机和情绪问题(Gardner, 1985; Panksepp, 1988)。此外,如前所述,认知革命主要关注人类的认知,因此在动物研究领域,行为主义仍然占据主导地位。只有少数科学家,如哈佛大学的唐纳德·格里芬(2001),推动动物行为学领域在思维上变得更加开放,但他主要关注认知领域,而从神经科学角度来看,这比情感问题更难解决。
公平地说,行为主义者的目标是创建一种高度可重复的科学,研究者可以明确”行为控制”的变量(这个术语用于指定将习得行为引导到可预测方向所需的精确环境条件)。他们中的大多数人从未真正声称要理解控制动物行为的基本机制。在他们有限的领域内,他们只是想在良好控制的环境中明确和预测动物的行为方式,而不是在它们自然状态下的真实世界中(那是动物行为学家的领域)。因此,他们建造了人工隔间(斯金纳箱),在那里动物外部环境的每个方面都可以被控制和系统地操纵。行为主义者根本不关心这些生物体内部发生的未观察到的事件,并且他们不相信这些事件能对行为的科学理解做出贡献。然而,悲剧在于,一旦神经科学成熟,许多此类事件,甚至是情感事件,都可以被科学研究。但行为神经科学家在很大程度上对研究它们不感兴趣,实际上是抵制的。大脑功能中具有深刻心理意义的方面,例如情绪的初级加工本质,在此时已成为可解决的科学问题,却被忽视和有意忽略。因此,神经科学家未能处理情感感受这一主题,直接源于行为主义的寒蝉效应。在动物研究中,这种情况至今基本未变。
现代神经科学革命始于大约40多年前,随着研究大脑工作方式的奇妙新程序的发展,最终形成了今天的神经成像设备,使研究人员能够观察活体(在活生物体中)人类大脑在执行各种活动时内部发生的情况。许多被这项神奇新技术所吸引的人接受的是行为或认知传统的教育。前者不接受情感感受作为其研究计划的一部分。后者倾向于将情感感受视为认知过程的一个子集,这至少在大脑组织的初级加工层面是一个重大错误,而这正是我们在这里主要关注的。认知是由感知、学习和更高的大脑功能创造的。原始情感是生存的祖先工具,具有用于各种”低级”大脑功能的专用回路。尽管人类的认知心智功能现在被普遍接受为事实,但大多数从事动物研究的研究人员仍然坚持行为主义学说,并会忽视、否认或对动物情感生活的存在保持不可知论态度。
如前所述,某些动物行为学家在认知动物行为学的旗帜下,确实开始思考动物的潜在心智能力(Griffin, 2001)。但通常大多数人避免讨论情感问题,很少有人从事情感大脑研究。这就是导致现代神经科学倾向于拒绝其他动物情感存在,因而拒绝系统科学研究的思想脉络。这第一条思想脉络植根于错误的古老信念,即心智是生机论的——它是一种独立的、非物质的力量,无法被科学审视。如前所述,这种将意识等同于生机论的观点是不正确的。初级加工心智——内在进化价值的体验——是大脑的一种功能,可以像任何其他生物功能一样进行科学分析(实际上,就像自然界中任何其他推断的功能或过程一样,例如物理学中的重力或量子粒子的活动)。
另一条思想脉络,说服神经科学家拒绝或忽视其他动物的情感问题,其根源在于19世纪后期,当时威廉·詹姆斯和卡尔·兰格(1834-1900)独立且几乎同时发展了一种外周反馈情感理论。他们将情绪行为(如逃离可怕情境)视为一种自动的、反射性的身体反应,本身没有情感。他们提出,关于这些身体反应的信息随后被反馈到大脑的思考和观察部分,即新皮层,后者在认知上体验情绪。因此,更高的大脑功能被认为产生了情感体验(Damasio, 1994; James, 1884/1968; Lange, 1885; LeDoux, 1996)。所以你不会因为害怕而逃离挥舞刀子的小偷;相反,你之所以感到害怕是因为你在逃跑,这创造了你身体感觉的各种变化,由更高的大脑功能”读出”。公平地说,我们要指出,心理科学中心智的伟大捍卫者威廉·詹姆斯也注意到,所有本能都有一种感觉,并且感觉和情绪反应是同时发生的(这是我们在这里捍卫的立场)。
尽管现在有科学证据表明,情绪行为的表演可以产生微弱的情感感受变化(Clynes, 1977; Schnall & Laird, 2003; Stepper & Strack, 1993),并且这种效果也可以通过人类大脑内的情绪动作意象获得(Panksepp & Gordon, 2003),但几乎没有证据表明,情绪动作期间的强烈情感感受需要从外周身体向大脑的反馈。相反,大多数证据表明,原始的情绪感受是由大脑组织直接产生的,实际上是由那些产生本能情绪动作的回路产生的。这并不意味着来自身体的输入没有影响。它们确实可以增强或削弱大脑内产生的感受。但它们在产生我们情绪感受的特定方式上并不起决定性作用。无论如何,120年前提出的James-Lange理论的经典解释,仍然是那些对大脑皮层下区域知之甚少的人所青睐的情绪产生方式的观点。
直到今天,还没有坚实的实验证据支持James-Lange理论的传统版本。然而,数据支持威廉·詹姆斯关于初级过程情绪的另一种推测——本能动作具有感受成分——而他的传统皮层读出理论可以帮助我们理解大脑如何理解其情绪。因此,根据我们目前的知识,大脑以两种方式产生情感:大脑的较低部分可以产生特定的情感感受,准确地发出身体需要什么(内稳态和感觉情感)以及大脑需要什么(情绪情感)的信号。然后我们的高级大脑以各种各样独特的认知方式处理这些心理力量,这通常为”人间喜剧”增添了趣味。此外,所有感受都有一个唤醒强度维度,这通常是许多不同感受所共有的。
然而,还应该认识到,大脑和身体有许多唤醒系统,包括一个主要的应激轴(垂体-肾上腺系统),如果在没有真正的情绪被唤起的情况下激活这些系统,那么人们会倾向于根据环境所促进的情绪场景来解释这种唤醒(Schacter & Singer, 1962)。一般唤醒本身并不构成情绪。一个人还必须以多种方式感到好或坏,这些方式对应于各种本能的表演冲动。当某人生气时,他可能想打某人。在皮层下初级过程层面,打某人的冲动同时伴随着愤怒的情绪感受。这就是迄今为止数据所显示的。但我们还需要指出,每一个科学事实总是有多种解释。科学的目的是在这些解释中进行筛选。这就是为什么去皮层实验如此重要,它表明情绪感受在大脑上层(新皮层)区域遭受大规模损伤后仍能存活。
如果您对我们上述关于James-Lange”身体反馈”情绪理论的观点概要感到满意,可以随意跳到下一节,该节专门讨论安东尼奥·达马西奥的有影响力的观点。但如果您对更详细的讨论感兴趣,请继续阅读……
尽管我们不赞同James-Lange反馈理论(或其现代”读出”后代),但我们是詹姆斯的崇拜者。如前所述,将外周身体骚动”读出”到高级大脑区域的概念并不是他关于情绪的唯一理论观察。在我们看来,他还更正确地提出,每一个本能情绪反应都伴随着特征性的感受。如果他知道这种本能反应是由独特的大脑回路产生的,他可能会推测没有必要假设一个认知”读出”来产生情绪感受,尽管沉浸在我们的感受中,甚至通过我们的意识觉察能力来修改它们,确实是我们高级认知装置的一部分。这就是为什么情绪调节如今在心理学中是一个如此受欢迎的话题(Gross, 2009),对心理治疗也非常重要。无论如何,正如我们将在本书中论证的那样,原始的情绪感受是皮层下回路的一部分,该回路也产生情绪行动准备。由于思想史的沉重负担(考虑激进行为主义的案例),詹姆斯理解情绪感受的另一种方法直到最近才得到充分发展(Panksepp, 1982, 1998a, 2005a)。
我们现在知道,来自身体的反馈总体上不可能是感受产生的主要来源。脊髓高位损伤导致没有来自损伤水平以下的躯体感觉输入的四肢瘫痪患者,基本上具有正常的情绪感受(Borod, 2000)。当然,他们的脊髓损伤保留了功能性自主神经,如迷走神经,以及可以影响各种大脑区域的血液中循环的内分泌因子。因此,特别重要的是要注意,即使是脊髓高位横断或产生”闭锁综合征”类型的脑干损伤的个体——只能移动(因此用)他们的眼睛或脑电波进行交流的人——仍然有情绪感受(Bauby, 1997; Birbaumer, 2006; Laureys et al., 2005),尽管身体感觉输入相当显著地减少了。
沃尔特·坎农(Walter Cannon, 1871-1945)是哈佛大学研究外周自主神经系统的生理学家,他提出了许多反对詹姆斯-兰格情绪观点的有力论据,并主张情绪性是大脑的内在功能。坎农指出,许多自主神经反应需要时间发展,不能足够快地反馈到大脑以产生即时的情感反应(Cannon, 1927)。他得出结论,情感不是反馈的问题,而是从大脑本身产生的。保罗·麦克莱恩(Paul MacLean, 1913-2007)是一位医生,他首先通过提出人脑中古老哺乳动物层的概念——“边缘系统”(limbic system)——来更详细地从进化角度发展这一想法,该系统负责基本的社会情绪。麦克莱恩在20世纪50年代和60年代开始对癫痫患者的情绪变化进行深入的大脑分析,随后开发了性行为和各种其他社会展示的动物模型(70年代和80年代)。凭借相当丰富的想象力,麦克莱恩(1990)设想了情绪性(包括情感体验)如何与边缘系统中的各种原始结构相关联。事实证明,麦克莱恩并没有掌握所有正确的细节(谁能做到呢?),为此他受到了各种”少壮派”的不公正批评(关于反驳,见Panksepp, 2002)。例如,麦克莱恩认为海马体是最重要的情绪大脑结构之一,但事实并非如此。正如我们将在[第6章]中看到的,海马体对记忆形成非常关键:自传体记忆的编码和我们空间环境的映射。不过,它也促进对恐惧事件发生地点的学习,海马体的腹侧部分在情绪学习中相当重要,特别是与空间相关的问题,如位置条件反射。然而,人们也可以唤起某些强烈的情绪,例如,通过局部注射催产素(oxytocin)到海马体可以轻易使大鼠产生勃起(Melis et al., 1986)。
然而,缺乏证据并不是一些研究者拒绝皮层下边缘大脑产生原始情感体验这一观点的主要原因。一些研究人员极其不喜欢哺乳动物”情绪大脑”概念(即边缘系统)在解剖学上的不精确性,一些人也拒绝接受情感体验可以直接从皮层下系统的活动中产生的观点。事实上,如前所述,大多数情绪学者仍然更倾向于詹姆斯-兰格的观点,即情感是从更高的大脑皮层区域产生的,在那里情绪行为被新皮层解读。
在现代”解读”理论发展的同时,本书的资深作者正在发展基于进化的跨物种”情感神经科学”(affective neuroscience)概念,详见早期的一本书(Panksepp, 1998a)。麦克莱恩和潘克塞普的方法有很大的趋同性,尽管潘克塞普在职业生涯开始时就开始发展情感神经科学,而麦克莱恩在职业生涯末期越来越倾向于动物神经科学模型。同时且独立地,两人都对理解大脑的社会-情绪网络产生了兴趣——特别是分离痛苦、社会联结和嬉戏。两人都是坎农和达尔文的追随者,因为他们认识到情绪感受是不同大脑网络中特定活动的直接反映,而不是外周反馈或更高大脑的解读。根据这一逐渐成为少数派立场的替代观点,古老的情感大脑被设计为本质上以情感-本能的无条件反应来预测生活挑战事件,这有助于相应地引导习得行为和思维。
尽管现代解读理论在许多细节上与詹姆斯-兰格模型不同,但原则仍然相同:大脑的情绪状态是对较低大脑或身体过程的更高大脑反应或反映。著名的神经科学家强烈主张,我们与其他哺乳动物同源共享的古老皮层下大脑区域不具有内在的情感属性(Damasio, 1999; LeDoux, 1996; Rolls, 2005)。顺便说一句,当本书准备付印时,达马西奥(Damasio, 2010)做了180度大转弯,明确认识到皮层下功能在心智构建中的重要性,尽管他仍然认为情绪感受存在于高级皮层区域。就现代神经科学导向的解读理论家对情感(情绪的感受维度)表达的兴趣而言(这种情况很少见),他们倾向于得出这样的结论:情感体验仅在无意识的情绪信息被大脑的认知-思维部分(特别是新皮层)解读时才会产生。这导致了当前关于情绪感受和所有其他形式的现象意识的最流行观点,即它们只是更高认知过程的一种变体。用我们的术语来说,认知科学家中的主流观点是情绪感受是大脑的三级过程。有些人甚至认为没有基本情绪——所有情绪最终都反映了更高的概念行为(Barrett, 2006)。虽然这对于三级过程情绪可能是正确的,但这种观点忽视了来自人类的大量可用行为证据(Izard, 2007)和所有哺乳动物基本情绪的跨物种神经证据(Panksepp, 2007d, 2008a)。(新期刊Emotion Reviews的一整期以及最近的一篇专著[Zachar & Ellis, 2012]都专门讨论了这个话题。)
我们将跳过心理学在过去几十年中关于情绪的大部分理论探讨,因为其中很少是基于对大脑的理解。然而值得注意的是,达尔文关于情绪身体表达的开创性著作《人类和动物的情绪表达》终于在1970年代和1980年代被研究者Paul Ekman和Cal Izard重新引入现代科学。他们延续了基本情绪理论(basic emotion theory)的传统,这一理论由他们的导师、临床心理学家Silvan Tomkins开创,他鼓励他们研究人类固有的情绪行为模式,这些模式在发育过程和跨文化中都可复制,尤其体现在面部表情中。Ross Buck和Robert Plutchik等人在不同方向上发展了基本情绪理论,特别是制定了新的内省和临床测量方法。事实上,在这一时期只有少数心理学家愿意讨论基本情绪感受的本质。其中突出的”反叛者”包括前面提到的Silvan Tomkins(1962, 1963),以及最近的社会心理学家Ross Buck(1999)。尽管心理治疗师长期以来一直认识到情绪感受的重要性,但目前越来越多的临床医生正在以新的方式关注情绪,以帮助建立情感健康(例如,见Fosha等人,2009a; Greenberg, 2002)。我们不会详细介绍这些有影响力的心理治疗师的观点,因为他们的工作没有聚焦于对潜在大脑机制的理解,但他们对新情绪动力疗法演变的影响将在第十二章由Panksepp进行情境化阐述(即,年轻的作者不希望与这些观点有关联)。
我们现在简要描述由著名神经科学家提出的三种现代读出理论(read-out theories):Antonio Damasio(1994, 1999)、Joseph LeDoux(1996)和Edmund Rolls(1999, 2005)。尽管我们不同意他们关于情感基础的观点,但我们钦佩他们令人印象深刻的实验贡献。显然,在下面的简述中,我们无法充分展现他们出色实证工作的细节——但每个人都在上述引用的专著中详细阐述了这些成就。我们还想强调,我们在下文中关于Damasio观点的描述在本书写作期间已经有些过时,因为他接受了皮层下对情绪感受和意识的强大贡献,这与Panksepp三十年来倡导的观点相当一致。然而,仔细阅读表明,Damasio仍然认为情绪感受主要是由高级感觉过程构建的。因此,鉴于这一及时的发展(Damasio, 2010),我们保持我们的论述不变,特别是因为我们的目的只是传达主流的历史观点,而Damasio是强化这一观点最有影响力的人之一。
我们认为这些科学家关于初级过程(primary-process)情绪感受的观点并未得到充分发展。事实上,很少有人强调大脑和心智的进化分层。因此,对我们来说,他们关于情感体验的主张常常似乎偏离了方向,尤其是涉及其他动物时。但我们也不希望彼此误解。我们怀疑这些受人尊敬的同事可能一直在指基于学习的次级过程情绪(LeDoux和Rolls)以及当认知和基本情绪结合成复杂混合物时产生的三级感受(Damasio)。这些研究人员在很大程度上忽视了进化而来的初级过程情感的可能性。我们在本书中的主要关注点是那些构成人类情绪基础的古老感受的本质。为了勾勒该领域的现状,我们现在简要总结这些著名的当代情绪研究者的”经典”方法。
Damasio对情感过程进行了一些出色的人类大脑成像研究(Damasio等人,2000),他提出了一种詹姆斯-兰格(James-Lange)类型的事件序列,这一序列先于情感的出现。他提出存在两个主要映射(maps),其中一个(原始自我, protoself)存储关于身体状态的信息。另一个主要映射存储关于环境的感觉信息。第三个映射过程(核心意识, core consciousness)起到连接两个主要映射信息的作用,并确定环境的某种状态与身体状态的某种变化相吻合。这产生了一种认知感受(feeling of knowing)环境对象的体验。这种认知感受是一种意识体验,一种”内在感觉”;它是”发生了什么的感受”,但它不是情感。Damasio将这种认知感受称为躯体标记(somatic marker),因为原始自我的身体反应标记(评估)环境中的感觉刺激。核心意识结合这些刺激和反应,并产生认知对象的非情感性感受。
Damasio认为核心意识是一种转瞬即逝的现象,以连续但不连贯的脉冲形式表达。当将记忆和复杂认知的新皮层能力加入其中时,核心意识的脉冲就可以被记住,人们可以理解它们。然后意识在时间上得到延伸,变成自传式的,因为个体可以记住他或她生活中的事件。这允许对关于对象的感受进行智能反思的能力,这一过程产生了情感。因此,情感的个人意义生成是一种新皮层的成就。
Damasio认为,只有少数灵长类动物能够产生如此延展和自传式的意识。因此,人类和少数与我们亲缘关系较近的哺乳动物是唯一能够充分体验情感(affects)的动物。在他倒数第二本书《寻找斯宾诺莎》(Looking for Spinoza)中,Damasio(2003)进一步强调(重复了太多次):“动物有情绪行为,而我们人类有情绪感受。”Damasio的经典理论从根本上说是James和Lange的”读出”或”反馈”理论的一个变体,但它在富有成效的方向上发展了这些理论。就他谈到原我映射(protoself maps)存储关于身体状态信息而言,Damasio至少认识到大脑本身能够产生情感(即使他称之为”仿佛”情感,并将所有情绪感受定位在大脑的较高位置)。然而,如前所述,在他最近的著作中,Damasio(2010)已明确接受动物确实有情绪感受,皮层下脑区具有适当的条件为体验到的感受乃至意识做出重要贡献。这是Panksepp四十年来一直坚持的立场。
LeDoux在大鼠恐惧条件反射的大脑机制方面做了一些最出色的工作,他也区分了情绪(emotion)和情感(affect),认为情绪是一种纯粹的生理反应,不含情感。情感是某种情绪的后续产物,当情绪生理学被支持工作记忆(working memory)的前额皮层部分读出时就会出现。工作记忆的基质位于前额皮层的背外侧部分,这是大脑最智能或至少最善于思考的部分。工作记忆可以看作是一个用于思考当前信息的心理工作空间(详见[第6章])。例如,当你阅读这段文字时,你会记住一些要点,同时可能会想起上周读过的一篇相关文章。所有这些想法都是你工作记忆中的项目。因此,工作记忆是大脑一个高度智能的功能,能够理解输入的信息。当人理解事物时,会将许多信息片段整合成一个连贯的概念。LeDoux指出,工作记忆执行多种认知任务,其中之一就是创造情感。根据LeDoux的观点,情绪的生理学(行为、内脏和低级别无意识的大脑反应)在大脑的这些认知区域转化为情感感受状态。
需要注意的是,LeDoux的研究几乎完全集中在恐惧(FEAR)上,也指向古老的皮层下区域作为恐惧的情绪-行为和自主神经(但非情感)基质。他的研究揭示了杏仁核(amygdala)这一长期与恐惧相关的皮层下结构如何在恐惧条件反射的产生中发挥核心作用,但不产生感受。杏仁核由十几个专门化的细胞群或核团组成,每个核团执行略有不同的功能。杏仁核的中央核在无感受的恐惧反应的下游生成中发挥主要作用,尽管从情感神经科学家的角度来看,它与其他更深层的结构(特别是导水管周围灰质)一起构成了恐惧系统的一部分。杏仁核中的其他一些外侧核团在条件学习中发挥作用,但不在恐惧本身的产生中发挥作用(有关恐惧系统的更多细节,见Panksepp, 1991,以及本书[第5章]和[第6章])。
不知何故,在LeDoux 1996年的书之后,将杏仁核视为所有恐惧,实际上是所有情绪的源泉,已成为流行的说法——这是一种可悲的错误观点。杏仁核完全受损的个体(即患有先天性Urbach-Wiethe病的人,导致杏仁核逐渐钙化和破坏)仍然可以体验担忧、恐惧和许多其他情绪。此外,玩耍(PLAY)、悲伤(GRIEF)、照料(CARE)和寻求(SEEKING)唤醒并不显著涉及杏仁核。实际上,杏仁核的众多亚核中只有一个,即中央核,是初级过程情绪系统的一部分,该系统有助于将进化提供的恐惧状态与高阶学习过程整合(产生次级情绪)。相比之下,LeDoux和其他恐惧条件反射理论家认为杏仁核的中央核只是各种恐惧反应(如僵住、心跳加速、血压升高、恐惧诱发的排便和排尿,以及许多其他应激反应)的”输出系统”。LeDoux和其他恐惧条件反射研究者尚未明确考虑一个整合的恐惧系统,及其连接杏仁核与许多其他脑区的众多下行和上行成分,足以产生恐惧的原始感受。他们倾向于假设情绪感受从新皮层的更高区域出现(LeDoux声称他对人类情绪感受感兴趣,而不是动物的情感过程)。我们不同意这种观点,因为我们认为,如果不理解动物的情绪感受,就无法理解人类的情绪感受。
我们理解Rolls认为,在动物中,情绪是对各种刺激的非情感性评估,而感受只有在各种身体感觉被三级脑过程(即新皮层)重新解释时才会出现,这些过程精化出语言等符号功能。他的卓越研究专注于感觉处理,特别是味觉功能。他认为非情感性情绪反应发生在皮层下结构中,包括在早期表述中,一些在新皮层之前进化的较古老的大脑皮层区域。总的来说,认为情绪性感受在大脑较高皮层区域产生的假设,与证据相矛盾,这些证据表明能够精化奖励和惩罚的情绪系统位于更深的大脑区域。我们认为,更可能的是较深层结构对旧皮层结构进行编程(或教导)如何产生评估。例如,在恐惧条件反射中,正是恐惧系统(所谓的UCR)的唤醒使杏仁核中的条件反射得以发生。换句话说,较新的皮层结构能够产生评估这一事实,并不排除大脑较深区域在产生次级评估所基于的原始感受方面的基本参与可能性。
不过,我们暂时遵循Rolls关于较低脑区产生的环境刺激的非情感性评估如何转化为现象体验的表述。这种据称是非情感性的信息,由高位脑干(丘脑和下丘脑)组织,可以朝两个方向发送。朝一个方向发送的信息将到达基底神经节——控制无感觉本能行为的深层前脑结构,如进食、排泄时采取特定姿势、性和攻击性姿态等。因此,例如(根据Rolls的说法),如果一只老鼠碰到一块奶酪,老鼠较古老的大脑结构会评估食物的味道和质地。这种评估是非情感性的,它产生的信息会被发送到老鼠的基底神经节,后者会指示老鼠继续吃奶酪。由较古老脑区产生的非体验性信息也可以朝另一个方向发送,向上到达新皮层(实际上在这种情况下,是一个称为眼眶额叶皮层(orbitofrontal cortex)的较古老皮层区域,就在眼窝上方)。然而,在他对情绪感受的一般表述中,需要一个庞大而复杂的皮层,如大多数人类所拥有的,来为非情感性的低级脑评估构建符号解释。这种符号解释可以用语言表达。这些符号性和语言性转换创造了情感体验,Rolls(追随许多哲学家的观点)称之为”感质(qualia)“。然而,在新皮层发育较弱的动物(如老鼠)中,据称没有情感伴随情绪行为。这是因为这些动物的上层大脑很少有合适的类型来产生情绪评估的符号概念——这些概念据称是产生情感所必需的。因此,Rolls得出结论,”不聪明”的物种没有情绪体验——因此我们在实验室中常规研究的动物,特别是老鼠和小鼠,不是情感性生物。
总结一下,根据Rolls关于意识的一般概念,如果你品尝一勺美食厨师制作的芝士蛋糕,较古老脑区的各种结构(包括眼眶额叶皮层)会评估关于蛋糕味道和质地的非情感性信息。这些信息会被发送到你的基底神经节,后者会指示你吃更多蛋糕。此外,你的旧皮层会将信息发送到你的新皮层,新皮层能够符号化并因此谈论吃这种精致甜点的愉悦情感体验。因此,对Rolls来说,语言表达能力或至少概念化评估的能力是情感体验的必要条件。在他看来,只有人类以及少数其他聪明物种才有情感体验。
Rolls表述的最大问题可能是,他使用感官情感来讨论情绪情感,在我们看来这是一个范畴错误。同时,自从撰写他的第一本关于情绪的书以来,他提供了大量人类脑成像数据,显示眶内侧额叶皮层(orbitomedial frontal cortex)(一个古老的皮层区域)如何参与产生对食物味道和质地变量以及愉悦触觉的享乐价值(hedonic value)反应(Rolls, 2005)。简而言之,他的工作更适用于感官体验产生的情感,而不是我们在这里讨论的情绪回路类型。
简而言之,由于这一观点在全书中都有总结,经典的情感神经科学视角认为,古老的情绪回路集中在大脑的原始区域,但与高级脑区有大量联系。情绪系统是根据这些回路的特性来定义的,这些特性至少有七个特征,如[图2.1]所总结的,包括:(i) 少数几个能够初始激活情绪的无条件刺激,(ii) 独特的无条件行为反应以及触发多样化的自主性身体变化来支持这些行动,(iii) 通过基本学习机制(即控制激励显著性(incentive salience))来筛选和评估并发的传入刺激的能力,(iv) 在无条件刺激消失后仍持续的正反馈,(v) 受高级三级过程认知功能的调节,(vi) 情绪系统强烈影响高级心理过程,以及 (vii) 整个系统产生独特的情感感受,感受最重要的产生器位于皮层下指令回路(如[图2.2]所示)。我们要强调的是,人们永远无法对初级过程情绪有一个科学上充分的文字定义;这样的定义必须基于神经回路标准,随着越来越多的可复制证据的积累而不断完善。
当然,每个初级过程情绪系统(SEEKING、RAGE、FEAR、LUST、CARE、PANIC/GRIEF和PLAY)都有自己特定的基础结构,与其他情绪系统以及一系列一般性唤醒功能进行抑制性和协同性的相互作用,例如,由广泛分布的乙酰胆碱、去甲肾上腺素、多巴胺和血清素系统控制,这些神经元在所有脊椎动物的相同古老脑干区域中定位(见[图1.1]的大致近似)。每个系统都是纵向组织的,从较低的中脑区域延伸到大脑的较高内侧额叶皮质区域。所有情绪系统都倾向于位于中线附近,这突显了它们在大脑进化中的非常古老的地位。[图2.2]提供了SEEKING系统及其各种功能连接的卡通式总结(关于解剖连接,见[图3.1])。下一章将深入讨论这个极其重要的情绪系统。多巴胺位于这个庞大情绪系统的核心,几乎控制着有机体所做的一切。它与其他脑区的相互作用如此广泛,以至于它有助于促进大多数其他情绪冲动。
图2.1. 基本本能情绪系统的定义特征的示意性总结。它们都有少数几个 (1) 内在输入,行为主义者称之为无条件刺激(UCS);(2) 各种本能行为和身体,特别是自主-内脏的输出,行为主义者称之为无条件反应(UCRs);(3) 各种其他刺激进入高级脑区的输入——潜在的条件刺激(CS)——如果它们预测奖励和惩罚,则由情绪系统控制(产生一些人所说的”激励显著性”);(4) 情绪持续的时间超过激活系统的刺激,无论是外部刺激(UCS)还是内部沉思,例如那些来自 (5) 高级皮质区域的刺激,特别是额叶皮质激活或抑制情绪,以及 (6) 情绪系统显然有能力控制和修改高级大脑功能。情绪的情感感受主要由属性4总结的内部大脑过程产生。然而,正如属性7所强调的,系统的所有其他方面都可以修改和调节情绪反应的强度、持续时间和模式。因此,最终的情感是定义每个原始情绪网络的所有BrainMind属性相互作用的结果。
同样,去甲肾上腺素,一个更古老的系统(因为细胞在大脑中的位置更低),在每种情绪唤醒期间都能促进注意力,但对愉悦感受的促进作用更强。乙酰胆碱也起到同样的作用,但通常更多地针对负面情绪。对于我们将在后续章节中讨论的所有初级过程”情绪-指令”系统,都需要牢记这种通用复杂性。情绪反应的大部分特异性是由特定脑回路中特定类型的谷氨酸能(兴奋性氨基酸)影响所促进的,还有许多神经肽(氨基酸链,见[图13.1])为许多情绪提供特异性。
情感神经科学方法并不认为情感感受是被高级认知脑功能”读出”的,尽管这些功能与大脑心智的那些区域存在广泛的互动。情感状态是每个情感操作系统不可分割的一部分。然而,这并不意味着高级认知机制不与这些古老的力量互动或对其进行反思。不仅原始情感系统调节和激励高级认知活动,而且它们也必然是高级心智装置极感兴趣的状态,这取决于儿童的养育方式,这些状态常常显得非常令人困惑。例如,被诊断为边缘型人格障碍(BPD)的人,这是一种成年发展性情感问题,通常有动荡的社会关系,因为情感不安全感,如恐慌/悲伤系统的失调感受。这些感受可能导致”拼命避免被抛弃的尝试”,矛盾的是,这往往”伴随着淡化亲密重要性的努力和/或旨在惩罚重要他人的攻击性行为……导致频繁争吵、反复分手和整体情绪不稳定的关系”,并”难以维持与他人的合作”(见Bartz等人,2010年,第556页)。
显然,高级大脑可以与低级大脑”斗争”。在上述情况下,过度活跃的恐慌/悲伤系统可能导致人们试图以自我挫败的方式维持自尊。人们会认为催产素会让这些人变得温和,增加他们的信任感,但正如前述Bartz等人的论文所发现的,它实际上降低了他们的信任感和合作意愿。当高级、更理性的大脑试图应对低级大脑不断变化的情感地形时,像这样的矛盾发现并不少见,而且还没有人知道如何理解这些意想不到的结果。也许这反映出我们许多人对自己真实感受的强烈程度有点尴尬,所以我们掩盖它们,有时压抑它们到甚至感觉不到的程度(这种情况可能会导致述情障碍(alexythmia))。人们会期望在专业心理治疗师的帮助下,这些个体能够唤起催产素更加亲社会的感受(见第7章至第9章),以帮助情感心智与认知视角产生协同作用,这些认知视角可以有自己的想法,并且常常可以凌驾于情感心智之上。因此,似乎高级认知心智通常不愿意承认,也不准确读出低级情感心智中正在发生的事情。
图2.2. 大脑中完整寻求系统可能的半现实示意性概念描述,使用主要交互功能的解剖学近似(改编自Panksepp,1981年,经作者许可)。
Edmund Rolls教授,以及许多在非生物学领域工作的研究人员,认为我们使用词语来生成概念,从而产生情感的语义和概念构建(例如,Barrett,2006)。谁会否认高级心智可以极大地影响低级情感景观呢?然而,这里的大多数问题,可能导致激烈的争论,可能只是反映了一个事实,即不同的理论家在讨论超复杂、分层组织的心智大脑系统中不同的分析层次。似乎不可否认的是,所有哺乳动物都共享某些基本的初级过程情感系统。据我们所知,次级过程学习机制(例如,经典条件反射和操作性条件反射)在所有哺乳动物中也非常相似。然而,随着高级皮层认知区域在物种间的进化和分化,大规模情感-认知互动的门户出现了。这个门户在不同生物之间可能存在巨大差异。正是在心智发展的最后这个领域,出现了最大的科学困境。许多理论家(他们自己的大部分心理生活都在大脑心智处理的高级概念领域)倾向于将所有心理体验都置于心智的最高领域之内。这导致了一个不合理的假设,即低级大脑功能严格来说是无意识的。但这个结论根本无法从证据中得到证明(Merker,2007;Panksepp,1998a;Shewmon等人,1999)。
显然,科学家在得出情感网络位于何处的结论之前,需要考虑情感处理的所有层次,这些层次足够复杂以维持体验。我们认为,最好从嵌套层级结构的角度来设想大脑组织的各个层次如何共同构成完整的情感体验([图2.3])。在这种观点中,较低的脑心智功能被嵌入并在更高的大脑功能中重新呈现,这不仅产生传统的自下而上的控制,还产生情绪的自上而下的调节。这提供了双向控制途径,可以被视为”循环因果关系”的形式,尊重大脑作为一个完全整合的器官,可能存在剧烈的内心冲突。如果有时我们自己似乎没有尊重这一观点,那只是因为科学是一门旨在将事物拆解的智力学科,以便理解复杂机制和过程的细节。这是一种认识论,如果不将整体分解为部分,就无法获得详细的理解,尽管通常没有从这些部分重建整体的能力。每个曾经拆解过内燃机以了解其工作原理的人都知道,将散落在车道上的所有零件重新组装成一台正常运转的机器是一项更艰巨的任务。社会建构主义者通常没有机会详细研究大脑,但他们似乎相信,他们对情感概念”整体”的描述与那些试图理解大脑实际如何运作的人所处理的问题是相同的。
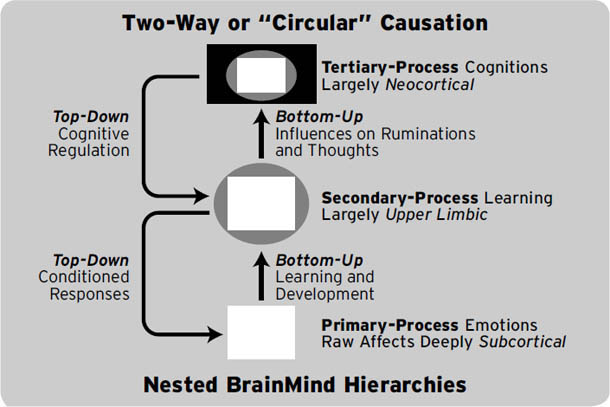
图2.3. 总结了在大脑的每个原始情感系统中运作的分层自下而上和自上而下(循环)因果关系。该图式总结了这样一个假设:为了使更高的心智脑功能运作,它们必须与较低的脑心智功能整合,其中初级过程被描绘为方形,次级学习过程为圆形,而顶部的三级过程为矩形。请想象每个符号都经过颜色编码,以更好地设想整合脑心智各个层次的嵌套层级结构(改编自Northoff等人,2011)。
公平地说,传统上倾向于心理生活的社会建构主义观点的社会心理学家和人格心理学家,最近开始假设情感的前概念基础。一些将视野局限于情感维度观点的人提出,某种原始的”核心情感”(Core Affect),其范围从消极到积极(效价),是构建所有其他情感感受的基本过程(Russell,2003)。他们观点的这一方面是发人深省且值得欢迎的,尽管他们往往没有充分考虑跨物种情感神经科学研究的现有证据(相关的已发表辩论及评论,见Zachar & Ellis,2012)。
社会建构主义者传统上认为,概念和语言是情感的标志,许多人现在仍然如此认为。如果动物无法概念化,它就无法体验情感。概念是一种抽象,通常从多种经验中获得。例如,椅子的概念是从看到许多不同种类的椅子中得出的,而椅子这个词代表了整体类别。第一次看到椅子时,你可能不知道它是什么,因为你肯定还没有一个概念知道它是一个休息的好地方。你必须学习到每一把单独的椅子都是更广泛群体的组成成员——这使你形成了椅子是什么的概念。
Rolls还提出,非情感性评价以某种方式也变成了概念,当你把这些概念用语言表达时,情感就产生了。只有聪明的动物才能做到这一点,这就是为什么他认为只有它们才能体验情感。我们认为这在进化上可能没有意义,因为我们知道人们在拥有疼痛概念之前就体验到了疼痛。对于我们将在这里讨论的所有初级过程情绪也是如此。
然而,有些词代表概念,而有些则不代表。正如我们在上一章中提到的,当你第一次看到红色时,你很快就知道了关于这种颜色你直接能知道的一切。你的视觉体验不是从其他经验中抽象出来的,除非在某种程度上你的视觉系统在发育过程中是逐步构建的。看到红色(或黄色或棕色)不是一个概念。因为你足够聪明,能够以语言的形式操纵符号,你可以使用诸如红色、深红、绯红和宝石红之类的词来区分和标记你体验中的细微差别。但看到红色的原始现象体验不需要智力。因此,像椅子这样的词代表智能概念,而像红色这样的其他词代表不需要智力的原始体验,当然,除非你想标记这种体验。
我们认为,基本情感属于原始体验的范畴,就像看到一种颜色一样,语言只是标记和表示这些体验。但情感体验本身,就像看到红色一样,不需要任何概念智力。人类可以使用词语来标记他们的情感,但他们不需要词语来体验它们。因此,我们使用词语并不一定意味着其他动物需要精通语言概念才能体验情感。原始情感确实是前语言体验——所有哺乳动物共有的体验,也许其他动物也是如此(Huber等人,2011)。
达马西奥(Damasio, 1999)关于情感意识的复杂观点——他在较低的无意识过程和大脑中相当高级的有意识过程之间划了一条线——仍有待实证证据的证实;他从未清楚说明其理论的关键测试应该是什么样的。尽管他的躯体标记假说(somatic marker hypothesis)——即来自身体的信息被转化为指导行动的感受——已经引起了大量实验关注(到目前为止记录好坏参半),但这些实验中很少真正监测所研究人类受试者的情感变化时间进程。此外,神经科学家对达马西奥假设的两个主要映射图(maps)的细节还没有清晰的认识:一个是关于身体变化的映射图,另一个是关于外部世界的映射图。他们也还不知道这些映射图是否通过他所称的核心意识(core consciousness)的高阶映射(higher-order mapping)进行整合。需要进一步的研究来检验这个观点:核心意识通过整合关于身体和环境的映射图信息,产生对正在发生的事情的内在情感感受。我们倾向于不同意他1999年的观点,不仅因为它没有得到足够详细的阐述。然而,我们很高兴看到,当本书付印时,达马西奥(2010)的观点发生了根本性转变:他现在接受皮质下结构(subcortical structures)确实对各种情感体验有贡献,这一观点在近半个世纪以来一直有坚实的实证支持。
勒杜克斯声称,情感源于支持工作记忆(working memory)的新皮质(neocortex)部分:背外侧额叶区域(dorsolateral frontal regions)。然而,现在有大量证据表明,在强烈情绪状态下,人类大脑表现出背外侧额叶区域激活的降低,而这些区域正是勒杜克斯和其他人确定为工作记忆基础的区域(Goel & Dolan, 2003; Liotti & Panksepp, 2004b; Northoff et al., 2009)。相反,当人们从事认知性、非情绪性活动时,这些背外侧额叶区域最为活跃。如果背外侧额叶皮质在情绪发作期间相对如此安静,它怎么能成为情感体验的源泉呢?我们同意这是大脑中我们人类以认知反思方式思考情感体验的主要区域,但更古老的内侧额叶区域(medial frontal regions)很可能是我们反复思考和沉浸于情绪困扰和其他感受的大脑区域。大脑的这个内侧部分在抑郁症患者中通常过度活跃(Northoff et al., 2011)。
那些支持读出理论(read-out theories)的人通常认为情感是认知构建物(cognitive constructs)。然而,据我们所知,新皮质(大脑的主要认知结构)在单独作用时无法产生情感。上述三位研究者似乎都同意,关于感知刺激和身体反应的信息是非情感性的。但是,非情感性信息如何通过本身无法产生情感的新皮质系统来创造有意识的情感体验呢?读出理论充满了问题和矛盾。这些当代理论中的每一个,在其经典形式中,都选择将其他动物排除在”情感圈”之外——即体验和以诸如热切期待、愤怒、焦虑、性感受、母性温暖、分离的心理痛苦或嬉戏的社交快乐等感受来响应事件的能力。我们将展示大量现有证据表明并非如此。事实上,如果仔细阅读勒内·笛卡尔的《灵魂的激情》(Passions of the Soul),很明显,即使是二元论(dualism)之父也可能接受其他动物确实有一些粗糙的感受;它们只是没有足够的”广延实体”(res extensa)(高级心理能力)来深思熟虑地反思它们的初级过程(primary-process)心理状态。如果许多其他意见领袖注意到并强调了这一点,也许对动物心智情感方面的研究就会蓬勃发展。如果詹姆斯-兰格理论(James-Lange theory)不是如此吸引人地反直觉,对创造性思维极具刺激性但迄今为止没有强有力的(因果)科学支持,其他动物现在不就已经被赋予了它们的情感感受吗(希望首先由行为神经科学家来做,因为当学者们无法处理这样的”显而易见的事”时,公众常常感到震惊,有时还会窃笑)?如果行为主义(behaviorism)对情绪的否认没有如此傲慢,我们现在可能已经对人类情绪有了丰富的理解,而不是仍然错误地相信情感只是高级心理能力的一种变体。我们的高级(新皮质)心理功能可以从我们的情绪中创造艺术和疯狂,但它们本身无法产生感受。这正是数据长期以来强烈表明的。
仅仅指出其他理论家立场中的弱点是远远不够的。我们可以解释行为主义的各种弊病和读取理论的失败之处。但这些批评都不能让我们有资格说情感是初级的、非认知的、前语言的体验。科学不是修辞。只有大脑研究以及精心设计的心理学实验才能让我们做出这样的断言。以下是通过这种方式获得的一些确凿证据的简要概述,我们将在后续章节中更详细地阐述,首先讨论SEEKING系统,然后依次讨论RAGE、FEAR、LUST、CARE、PANIC/GRIEF和PLAY。这些确凿的证据让我们能够得出结论:其他动物确实是具有情感的生物,并支持少数派观点,即如果我们理解它们的情感感受,我们将获得一门关于这些心脑能力在我们自己生活中的祖先来源的可靠科学。
如果这个论点是有效的,那么行为科学研究者在研究他们想要理解的生物时抛弃情感感受是一个重大错误。事实上,他们用于训练动物的主要概念——“强化”习得行为改变的奖励和惩罚——之所以能够成功运作,很大程度上可能是因为动物大脑中未被承认的情感原则。一旦我们认识到这是一个高概率的神经科学事实,而不仅仅是一个假设,我们就可以在利用临床前动物模型来理解人类情绪及其诸多障碍的知识应用方式上实现革命性转变。而这一切都不需要抛弃许多行为神经科学家收获的关于大脑行为、神经生理学和神经化学机制的出色知识——尽管这些科学家不能容忍关于动物感受的讨论。
那么生理心理学家是如何”偶然发现”让我们得出动物确实有情感感受这一结论的事实的呢?在上个世纪中叶,James Olds和他的同事Peter Milner做出了一个非凡的发现:所有动物,至少所有他们测试过的动物(以及此后测试过的所有动物),都会为了获得内侧前脑束-外侧下丘脑区(MFB-LH)的电刺激而努力工作到精疲力竭的程度,正如Jim Olds简洁描述的那样,这是一个极其广泛的系统(1977年,死后出版)。[图2.4]是这一系统在大鼠大脑中最早的真实描绘之一。该系统连接着大脑的下部、中部和上部区域。它是行为和心理连贯性最重要的大脑系统之一。
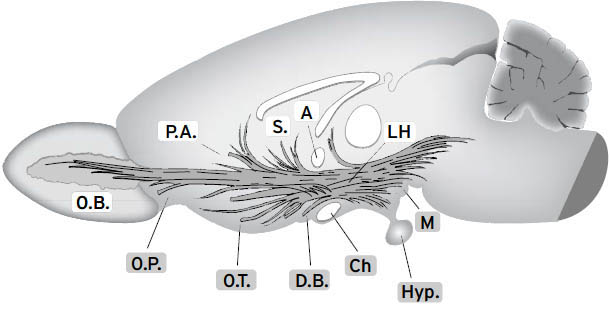
图2.4. 内侧前脑束(MFB)的示意图,连接中脑的中心区域与高级脑区。MFB穿过位于视交叉(Ch)正上方以及左右两侧的外侧下丘脑(LH),其余解剖学命名突出显示了嗅球(O.B.)、嗅脚(O.P.)、副嗅区(P.A.)、嗅束(O.T.)、Broca对角带(D.B.)、前连合(A)、垂体腺或以前称为脑垂体(Hyp.)和乳头体(M)。在中脑,MFB的下行分支部分投射到内侧区域,如导水管周围灰质。这幅经典图形改编自Le Gros Clark等人(1938)。
而且回想起来,刺激这个复杂网络会产生奖励性也就不足为奇了。但在1954年,这是一个惊人的发现,像野火一样席卷了整个心理学领域。动物倾向于通过执行诸如推杆之类的任务来持续努力获得这个系统的激活,这被称为自我刺激。这一发现完全是偶然的;Olds和Milner正在寻找用大脑刺激来增强学习的方法。但他们足够明智,转移了研究重点并深入研究这种新现象的本质(无疑是利用了他们自己的SEEKING系统)。显然,这种刺激具有高度的奖励性。否则动物为什么会如此努力工作呢?假设他们找到了大脑的”奖励系统”似乎是合理的,而这个过度的想法作为”权威”概念一直延续至今。尽管大脑中有许多奖励系统,但只有一个系统能够驱动动物积极地寻求所有其他类型的奖励,主要是感官和稳态奖励,这些奖励是它必须在世界中发现才能生存的。这就是为什么我们长期以来一直主张给它一个更恰当的、尽管不寻常的情感名称(Panksepp, 1981b, 1982)——首先是EXPECTANCY(期待),现在是SEEKING(寻求)系统。
当我们在下一章详细研究SEEKING系统时,我们将看到它的中枢位于大脑低级区域的神经网络中,包括腹侧被盖区(VTA)和外侧下丘脑(LH)。在那一章中,我们将解释SEEKING系统产生充满活力的探索和觅食,以及可以更好地描述为欣快兴奋而不是奖励或愉悦的情感——这种感觉是一种预期性的渴望和兴奋,在更认知的层面上,是离散期待的产生。正是这些高度充满活力的、欣快的觅食性与世界的互动,让动物觉得如此有奖励性。这些是位于某些人可能称之为快乐活力的核心的感受。
然而,在上个世纪中叶,SEEKING系统(寻求系统)还不为人知;科学家们当时考虑的唯一一种奖励是那些与恢复体内平衡相关的奖励。因此,食物、水、温暖、性满足等被视为有奖励性的体验,因为它们恢复了身体的平衡状态(这是驱力减少理论家的一个关键观点)。即使是像斯金纳这样激进的行为主义者也将体内平衡(驱力减少)视为奖励。体内平衡失调引发的饥饿和口渴的影响可以通过科学方法在生理上进行测量,例如,通过低血糖或低血容量来测量,也可以通过行为上增加的食物和水摄入量来测量。所有这些都可以在不需要提及饥饿或口渴等情感或动机状态的情况下完成(Skinner, 1953)。
行为主义者观察到,体内平衡失调,如低血糖,会使动物更倾向于为了食物而工作。然而,大多数行为研究人员最终发现,奖励的感官特性——诱因特性(incentive properties),如奖励的质量、数量和延迟——在控制学习方面比身体的平衡状态变化重要得多。换句话说,感官奖励越好,动物学习得越快。仅仅驱力减少并不那么有效。例如,虽然饥饿的动物会很容易学会为美味的食物而工作,但它们需要很长时间才能学会将食物直接自我注射到胃里,尽管经过长期训练,大多数动物最终甚至会学会这一点(Mook, 1989)。
由于SEEKING系统是研究最深入的情绪系统,尽管是在”大脑奖励系统”的名义下,让我们简要讨论一下它的特征,不引用科学文献,因为这些文献在许多资料中都能很容易找到详细内容(包括Panksepp, 1981b, 1998a),以及下一章。我们在这里介绍这个系统,因为这可能是科学家和感兴趣的读者都最难理解的系统,也因为它对所有其他情绪系统的正常运作都非常重要。因此,这个简短的概述是对本书其余部分许多内容的预示。
与只从行为角度考虑奖励的行为主义者不同,神经科学家对大脑功能感兴趣。因此,当奥尔兹和米尔纳发现动物会特别努力地工作以获得MFB-LH刺激时,这一消息席卷了心理学界。许多生理心理学家(当时他们被这样称呼)开始假设MFB-LH是所有类型的体内平衡和感官奖励的共同基质。带着这种想法,像潘克塞普这样开始研究这一现象的科学家最初假设,对MFB-LH的电刺激或药理刺激之所以具有奖励性,是因为它对应于各种消耗性奖励。换句话说,当MFB-LH的某一部分受到电刺激时,动物的大脑反应方式与动物享用一顿美餐时的反应方式相同。MFB-LH的另一部分会像动物解渴时那样反应。MFB-LH的另一部分会以动物参与有奖励性的性活动时的方式作出反应。
然而,实验证据并未遵循预期的模式。当动物找到它需要的资源并开始进行消耗性活动,如进食、饮水或性活动时,沿着MFB-LH的神经元放电会暂时但显著地减缓(Hamburg, 1971)。这表明MFB-LH刺激提供的奖励在体内平衡开始恢复之前是活跃的。实际上,当人类和动物处于体内平衡需求状态,并且在环境中有机会找到良好感觉时,MFB-LH最为活跃。
那么MFB-LH刺激实际上可能提供什么样的奖励呢?它当然不仅仅是体内平衡或感官奖励,尽管该系统确实对这些事件作出反应。一个线索可以直接从动物在接受此类大脑刺激时表现出的非条件行为中获得。大鼠在进行自我刺激时会变得超级兴奋。如果只是给MFB-LH”免费”的电击,而不需要大鼠为此工作,动物会四处移动,急切地探索它们的环境,即使是单调的环境,比如一个空盒子。它们探索所有环境,就好像在寻找什么东西。此外,一直令人困惑的是,为什么动物在MFB-LH自我刺激期间按压杠杆的次数远远超过获得所有”奖励”所需的次数。它们这样做似乎只是因为它们过度兴奋,这与消费奖励产生的愉悦状态不同。
MFB-LH 并不是大脑中唯一能让动物进行自我刺激的部位。例如,动物会按压杠杆来自我刺激隔区(septum)。但它们这样做的方式更有条理,通常每次电击只按压一次杠杆,而不是按压次数远超必要的次数。换句话说,MFB-LH 刺激会引起某种兴奋状态。事实上,动物可以区分——它们能够辨别——隔区和 MFB-LH 刺激之间的差异(Stutz et al., 1974)。显然,这两个刺激部位产生的是不同的体验。当人类大脑的隔区受到刺激时,人们通常会报告性感受。当他们的 MFB-LH 受到刺激时,他们报告的是更普遍的兴奋和期待感——难以用言语表达的感受。虽然隔区刺激确实参与了性活动的消费-高潮奖赏(consummatory-orgasmic reward),但 MFB-LH 的寻求系统(SEEKING system)则详细阐述了性欲的食欲渴望阶段(appetitive eagerness phase)以及对所有其他奖赏的期待。
结论是不可避免的。在认知层面上,MFB-LH 以一种欣快的普遍期待状态的形式提供情感奖赏,最初并没有明确的目标。MFB-LH 的刺激当然不会产生与我们在身体失衡恢复到体内平衡时所感受到的脑状态相对应的状态(即满足的感觉)。当动物满足时,它们往往会入睡。MFB-LH 刺激使动物保持清醒。在 MFB-LH 刺激下,动物显得热情高涨,渴望探索它们的环境。因此,人们会对世界感到更有兴趣,并制定未来计划——这显然是一种充满期待的兴奋状态。没有人报告过体验到明确的感官愉悦感,比如美妙的味道。MFB-LH 的唤起产生的奖赏更接近欣快感,而不是任何感官-身体愉悦。
此外,即使动物被去皮层(decorticated)——通过手术剥夺其新皮层——它仍然会工作到精疲力竭的程度,以获得 MFB-LH 刺激。因此,这种奖赏性情感不可能来自新皮层,因为这些动物没有任何新皮层。人们不得不得出结论,皮层下结构产生这些情感奖赏,以欣快性情感意识的形式——一种主观感受状态,人类和动物如此渴望,以至于他们会工作到精疲力竭来获得它。
在日常生活中,MFB-LH 以及寻求系统的其余部分,通常在动物处于体内平衡失调状态时更容易被唤起,但真正激活该系统的是世界上现成可得的好东西(“激励刺激”(incentive stimuli),正如科学家所说)。每个人都知道,身体的所有主要体内平衡失调都会让人感到不愉快。相反,与激励刺激互动,会引发摄入奖赏的愉悦感受,不仅预示着体内平衡的恢复,还会激发愉悦体验(Cabanac, 1992)。但”奖赏系统”并不是在为我们做这件事。它在做同样重要的事情——它让我们能够热情地追求奖赏。寻求(SEEKING),这个系统更好的名字,产生一种压倒一切的期待性欣快感,促使人类和动物寻找他们需要的资源。这个系统不仅帮助动物满足身体需求,而且正如我们现在所知,还满足许多其他高级情感需求,从对金钱和信息的渴望到音乐和其他审美体验。
其他六个情感系统不适合这种体内平衡解释,因为它们与满足身体需求的联系不那么紧密。其他情感更严格地与脑心(BrainMind)的内在方面相关,但所有这些都需要寻求环境资源。因此,在某种程度上,所有其他情感也依赖于寻求冲动的心理行为推动。从某种意义上说,寻求是所有情感系统的”鼻祖”。要满足欲望(LUST),人们必须寻求关系。要感受到温柔的关爱(CARE),人们必须寻求帮助那些需要帮助的人,尤其是婴儿。要感受到完全的愤怒(RAGE),人们必须寻求伤害那些想从你那里夺走资源的人。要很好地应对恐惧(FEAR),人们必须寻求安全。要让你的恐慌/悲伤(PANIC/GRIEF)为你服务,你必须寻找那些会支持你需求的人。要充满喜悦地玩耍(PLAY),你必须找到朋友。
显然,伴随寻求系统人工唤起的情感来自皮层下区域,正如大规模前脑损伤后自我刺激的存在所强调的那样(Huston & Borbély, 1973, 1974; Valenstein, 1966)。这长期以来一直质疑所有读出理论(read-out theories),这些理论声称情感体验是新皮层的成就。事实并非如此。当然,新皮层可能有助于从更原始的情感现象构建复杂的情感(三级过程情感(tertiary-process emotions)),这本身就是一个非常有趣的神经科学话题,但目前我们对这实际上是如何发生的知之甚少。上述分析也应该结束动物研究中长期存在的行为主义偏见:其他动物不是情感生物。
尽管关于SEEKING系统的相关数据比其他任何系统都多,但对其他每个初级过程情绪系统的研究都支持同一总体结论——原始情绪感受源自大脑皮层下网络,这些网络产生本能的情绪行为。所有其他哺乳动物都像我们一样具有情感生命。但我们不应声称它们的感受与我们的完全相同——进化总是在细节上产生变异——但我们都确实拥有属于相同大类的原始感受。在某些物种中,某些感受比其他感受更强或更弱,但它们在某种程度上都存在。它们位于我们心智的基础。如果是这样,我们可以理解自己情绪的一般原则和来源,如果我们详细研究这些系统在我们动物同伴身上的表现。大鼠和小鼠对这类研究来说非常合适。这项工作可以做得很好,对动物造成的压力很小。许多这些本能情绪系统可以在麻醉动物中研究(Panksepp, Sacks, et al., 1991; Rossi & Panksepp, 1992)。事实上,去皮层动物展现出所有七种初级过程情绪行为(Kolb & Tees, 1990; Panksepp et al., 1994)。
所有被研究过的动物都明显喜欢或不喜欢本书讨论的情绪系统人工激活所产生的情绪感受。这帮助我们理解为什么各种无条件刺激(UCS)和引发的无条件反应(UCR)对学习如此重要。正如动物会趋向于它们先前有过积极激励体验的地方(表现出条件性位置偏好),如进食、饮水或性行为,它们对接受促进这些行为的回路人工激活的环境也表现出类似的偏好。相反,它们会回避(表现出条件性位置厌恶)有过不愉快情绪体验的地方。它们远离曾经受到惊吓或伤害的地方;这些情绪是由环境事件产生还是由产生这些类型情绪行为的大脑系统人工激活产生并不重要。
其他相关实验表明,情感源自哺乳动物大脑的皮层下区域。例如,动物对它们服用过滥用药物的地方表现出偏好——这些药物在人类中诱发愉悦或令人向往的情绪状态。这些效应的关键网络位于皮层下。正是因为它们影响大脑情感系统,成瘾药物才能用于动物研究来理解人类成瘾的大脑机制。大多数研究者的隐含假设是,动物寻求这些药物是出于类似的情感原因,而不仅仅是学习”奖赏”,但这很少被承认(例外见Kassell, 2010)。我们现在知道,这些药物通过模拟在我们大脑中产生特定类型感受的神经化学物质来实现其在人类中的效应。在其他动物中不太可能有所不同。然而,成瘾有一个额外的特性——一个对抗过程,即当药物从系统中清除时会留下一个黑暗的情感空洞。而且这种可怕的后效应会随着某些药物(如安非他明、可卡因和阿片类药物)的消耗增加而变得更大。摆脱这些负面感受可能比某些药物产生的良好初始感受在成瘾中更重要(Koob & Le Moal, 2001)。
滥用药物分为两类:药理学上刺激SEEKING系统的药物和介导感官愉悦的药物,包括神经化学抑制PANIC/GRIEF系统,这会产生社会联结的温暖感受。像可卡因和安非他明这样的药物主要增强多巴胺的效应,多巴胺刺激SEEKING系统,唤起与外侧下丘脑(LH)电刺激所提供的相同的热切期待感。阿片类药物,如吗啡或海洛因,在化学上类似于介导感官愉悦和积极社会关系形成的内源性大脑化学物质(Panksepp, 1981a, 1998a)。这就是为什么梳理毛发对猴子来说是有奖赏性的(Keverne et al., 1989),以及为什么好友和爱人的陪伴会唤起我们的舒适和放松感受。正如我们将在第10章看到的,当阿片类药物直接施用到大脑中时,它们刺激的情绪感受类似于积极社会联结所体验到的感受,以及许多其他情感上令人向往的激励。其他大脑系统,如基于催产素(oxytocin)的系统,最近也被发现产生类似的效应。
但阿片系统遍布整个大脑。为什么我们应该相信这些美好的感觉仅仅由新皮层(neocortex)以下的大脑系统产生?通过动物研究,我们可以通过将阿片类药物注入特定大脑区域来直接评估这些问题。动物对注入原始皮层下大脑区域(如导水管周围灰质(PAG)和腹侧被盖区(VTA)——这些大脑区域通过内侧前脑束-外侧下丘脑(MFB-LH)发送通路)的吗啡表现出偏好,但它们对注入其他更高级大脑区域的吗啡不表现出偏好,尽管所有这些区域都有丰富的阿片受体(Olmstead & Franklin, 1997)。当这种药物被注射到深层皮层下区域时,动物表现出位置偏好这一事实表明,这些深层结构产生了奖赏性情感(affects)——即动物喜欢体验的情感。当相同剂量的吗啡被注射到大脑的许多更高级区域(包括皮层)时,动物不表现出这种偏好,这一事实表明这些区域可能没有同样高的能力来产生奖赏性情感感受。
除了对阿片类药物表现出位置偏好外,动物还表现出愿意工作以获得直接给予寻求系统(SEEKING system)的深层内侧皮层下位点的吗啡和可卡因剂量(Ikemoto, 2010)。因此,化学和电刺激大脑的发现是一致的。当其他药物被放置到其他情绪系统时,也会看到类似的效果。但随着人们从介导寻求冲动的神经网络转向其他情绪的神经网络,总体数据量会减少。这并不反映矛盾的证据;它只是反映了许多必要的研究仍有待完成这一事实。
动物也可以通过发声来表达它们的喜好和厌恶(它们的偏好)。从观察我们的宠物中我们都知道,动物的发声表明特定的快乐或不快。我们认识到当我们下班回家时家犬欢快的吠叫,我们也理解当陌生人在附近时愤怒的咆哮声。我们很容易区分抚摸猫时满足的呼噜声和不小心踩到它尾巴时的尖叫声。我们毫无问题地理解当我们把狗留在狗舍时的悲惨哀号和恼怒的猫发出的嘶嘶声。所有这些情绪发声都来自大脑的皮层下区域,在不同物种中具有非常相似的解剖结构和神经化学(Burgdorf et al., 2007; Brudzynski, 2010; Jürgens, 2002; Newman, 1988中有早期工作的广泛总结)。
最近关于情绪发声的科学工作大多是用大鼠完成的。例如,当大鼠彼此玩耍或被挠痒时,它们会发出50千赫兹(kHz)的高音(超声)啁啾声。当大鼠(雄性和雌性)期待性行为或任何其他各种奖励时,也会发出类似频率的发声(Knutson et al., 2002; Panksepp, Knutson et al., 2002)。因此,无论在大脑的哪个地方通过电刺激人工诱发50-kHz叫声,大鼠都会自我刺激这些电极位置(Burgdorf et al., 2007)。相反,当大鼠在社交中失败或周围有危险时(例如,附近有猫),大鼠会发出长长的22-kHz”抱怨”或”警报叫声”。这些在恐惧条件反射研究中连续施加足部电击之间尤其突出,当安全信号响起表明没有疼痛即将到来时,大鼠会叹气(Soltysik & Jelen, 2005)。令人惊讶的是,交配后,雄性大鼠也会发出22-kHz的发声。也许,仅仅是也许,这是让雌性知道他不再有社交情绪的声音报告。或者,也许这只动物正在发出虚假的”警报叫声”来让其他雄性保持距离(大鼠是滥交的),从而增加他成为雌性下一窝”宝宝”父亲的机会(当然,无需思考)。
这些关于自我刺激、位置偏好以及情绪发声和其他本能-情绪行为回路的事实,使我们能够得出哪些大脑区域对于产生原始情绪体验最重要的结论。这种证据对于基于事实理解大脑如何产生所有初级过程情感状态(无论是感觉的、稳态的还是情绪的)至关重要。
动物是否体验除基本情绪之外的初级过程情感?有足够的证据最终可以考虑感觉愉悦和不适的性质(即感觉情感,如味觉的愉悦和疼痛的痛苦),以及源于身体失衡的情感(稳态情感,如饥饿和口渴)。然而,我们不会在本书中广泛关注感觉和稳态情感,尽管我们认为动物很可能强烈地感受到它们。
为什么?很大程度上是因为这些情感的数据库不如情绪情感那么广泛,而且这些系统可能不是意识本身的必要基础。科学家也缺乏精细的操作手段,例如对相关脑区进行局部电刺激和化学刺激,以明确在动物身上唤起这类状态,从而对这些状态的情感特质进行因果实验。大部分可用证据属于相关性而非因果性或构成性范畴,因此我们知道当潜在的享乐刺激呈现给动物时会发生什么样的行为和脑变化,但我们不知道这些变化中哪些实际上导致了相关的情感。在缺乏此类数据的情况下,人们只能陷入基于相关性观察来论证因果联系的逻辑困境。
无论如何,关于稳态情感的实质性科学文献正在不断增长,特别是通过人类脑成像技术收集的研究。来自人类的脑成像证据表明,口渴、饥饿以及所有其他”身体-内脏”感觉都是在调节动物这些相同过程的深层皮层下结构中形成的(Denton, 2006)。同样,关于味觉电生理相关性的引人入胜的文献(Rolls, 2005)以极其精妙的方式揭示了感觉情感的可能本质。但认为这些发现能够解释情绪是一个范畴错误。
我们尚不确定感觉情感是在哪里产生的。最可能的答案是它们在神经系统的多个层次上生成,甚至可能在新皮层中。一些研究得最清楚的系统是介导味觉的系统(Berridge, 2000, 2004; Steiner et al., 2001)。通过对这一系统的研究,可以清楚地看到,在实验室大鼠中,甜味等积极的味觉特质在一定程度上是由基底前脑和苍白球周围的深层脑干结构介导的。密歇根大学的Kent Berridge和Susana Peciña已经确定了基底前脑的特定区域(腹侧苍白球(ventral pallidum))作为甜味神经处理的中枢(Peciña et al., 2006)。有时,研究人员暗示这些皮层下区域只是处理味觉信息,而这些信息在大脑更高处(如脑岛)转化为味觉感受,脑岛显然对厌恶感很重要(Craig, 2003a, 2003b)。人们普遍认识到,许多传入感觉系统在到达丘脑时会一分为二,刺激的情感方面分流到各种皮层下系统,而让我们能够有思考地区分各种感觉的更多认知信息则继续传向新皮层(关于味觉见Sewards, 2004,但同样的原理也适用于疼痛、触觉等)。我们怀疑较低的脑区本身足以产生他们研究的原始情感性味觉体验。但是,对于大多数感觉-情感体验的脑基质问题,目前还不像初级过程情绪情感的脑基质问题那样容易解决。
随着大脑的进化,较新的结构补充了较旧结构的功能([图2.3]),导致了随发育而变化的层级控制([图1.4]和[图1.6])。因此,在进化史的早期,与情绪、稳态和感觉体验相关的情感很可能严格来自大脑的深层皮层下区域,随着大脑进化,它们已经被更新添加的脑网络所精细化。也许某些情感的产生甚至已经被更高的新皮层区域”接管”,但我们在这些假设上还处于摸索阶段。我们只能确定一个事实,即在婴儿发育早期,所有动物都更依赖于较低而非较高脑结构的功能(Chugani, 1998)。
在成熟过程中,大脑的深层部分很可能可以编程——或”教导”——更表层的结构如何以特定方式发挥功能([图2.3])。因此,某些初级过程情感可能最初是在皮层下形成的,在个体发育过程中,这些功能得到完善,在某些情况下可能被新进化的更高脑区所接管。如果是这样,那么大多数情感很可能受到大脑更新进化功能的重大影响。这对于某些高度由认知介导的感觉情感尤其相关(例如,为专业品酒培养的那些感觉)。但在允许感受的微妙性方面,这往往是以感受的强度为代价的(即认知调节功能更多时候是抑制而非放大初级过程感受)。但在这里我们完全处于推测之中。
相比之下,我们可以确信,在大脑的低级区域,情绪感受比在高级区域更强烈——原因很简单:在所有被研究过的哺乳动物中,包括人类,电刺激在大脑低级区域用更少的电流就能诱发更强烈的感受。因此,刺激杏仁核(amygdala)在人类和其他动物中产生的情绪感受,不如刺激脑干区域(如位于中脑中心的导水管周围灰质PAG——大脑最古老的区域之一)产生的强烈。此外,如前所述,当新皮层(neocortex)缺失或在发育早期被移除时,人类和动物长大后在外在表现上都会比那些拥有高级脑区来抑制初级过程情绪性的个体更加情绪化。在没有新皮层(特别是额叶区域)的动物身上,比在拥有完整大脑的动物身上更容易唤起情绪表现。
这些事实对我们对情绪情感的科学理解来说无异于一种恩赐。产生原始情感的神经回路与本能情绪表达之间的紧密关系,使我们能够用可见的代理物(情绪行为)来研究我们无法直接看到的东西(情感)。为什么神经科学和心理学一直缺失这一洞见?也许是因为我们太习惯于将运动过程视为”单纯的输出”,而不是将其视为对整个有机体的整合过程。除非动物的大脑中有复杂的行动图式(action-schema),比如基本情绪,否则它们根本没有生存的机会。当我们开始思考地球上生命的本质时,这种复杂的”运动”大脑功能能够构成情绪感受的事实似乎很有说服力,而且现在的数据令人印象深刻地证明了大脑内情绪行动和情绪感受系统的一致性。
这使得基于动物大脑研究的大量可检验预测成为可能,特别是关于神经化学因素的研究,这些因素可以类似地应用于动物情感和人类体验的研究。所收集的知识对于生物精神病学和心理治疗科学将至关重要。显然,情绪情感对心理健康和疾病有着强大的影响。调节愤怒(RAGE),发展抵消恐慌/悲伤(PANIC/GRIEF)的能力(通过形成温暖的社会依恋),适应性地应对恐惧(FEAR),享受游戏(PLAY)的能力,优雅地满足欲望(LUST)的追求,以及以乐观的期待、同情和宽恕来面对生活,这些都是良好心理健康的基本要素。
情绪系统出错并不需要太多——这就是为什么情感失调(affective dysregulation)现在是、可能一直都是人类的常见体验。直到最近,心理学家才开始对积极情绪产生强烈兴趣(最全面的近期总结见Sheldon等人,2011年),甚至神经科学家和精神病学家也开始触及积极情绪的表面(Burgdorf & Panksepp, 2006; Vaillant, 2008),而不仅仅是研究大脑的”奖励系统”(这是一个误称,正如我们已经看到的,但我们将在下一章进一步阐述)。在初级过程情感的神经本质被阐明之前,精神病学和心理治疗将仍然缺乏严格和透明的科学基础。目前还没有公认的策略来解决这一困境。但是跨物种三角验证(cross-species triangulation)的情感神经科学方法(在行为、神经和心理证据线之间)已经建立了良好的记录,能够从我们大脑的深处揭示我们最基本情绪感受的来源。
人类情绪感受的机制不再需要保持神秘。如果我们在理解人类情绪情感本质的努力中采用从动物模型中获得的洞见,也许我们可以开始填补目前仍然相当大的经验空白。到目前为止,心理健康专业人员依赖的是各种不同的理论,没有一个是完整的或完全有效的。精神病学依赖的诊断类别与脑科学或我们对情绪大脑的理解几乎没有关系;相反,它们来自对外在认知迹象和症状的描述,大部分是通过语言报告的。精神病药物的发现主要是偶然的——当治疗其他疾病的药物的副作用被意外发现能产生有益的情绪变化时。在过去40年里,几乎没有发现新类型的精神病药物。随着对情感大脑更清晰的神经科学认识,新的医学发现应该会更快地到来(见Burgdorf等人,2011年)。
我们相信,一旦我们开始认真对待初级过程情绪行动系统,将其视为所有哺乳动物大脑中可预测组织的情感实体,我们在经验和理论理解方面就会取得重大进展。然后,我们可以在统一的理论框架基础上开发新药物和新疗法,而不是零碎地、偶然地开发。换句话说,我们可以使用临床前(动物)模型来研究精神疾病,在这些模型中我们操纵大脑的不同情感系统,并监测其他情感系统如何被改变(关于抑郁症建模,见Panksepp & Watt, 2011)。
简而言之,全面理解情绪影响背后的大脑机制似乎是当代精神病学的一个重要项目。这些知识还可以为心理治疗艺术提供更坚实的基础。在第12章,Panksepp将探讨由我们对情绪和情绪记忆神经基础的新兴理解所提出的心理治疗实践的新思路。一些发现完全出乎意料。再巩固(reconsolidation)的发现(第6章)表明,我们可以提取旧的和令人困扰的记忆,然后用较少情感困扰的阴影重新塑造它们。
对哺乳动物大脑主要过程情感过程的神经科学研究可以打开现象意识的潘多拉魔盒——即原始情绪体验实际上是如何在大脑中构建的。它可以同时为人类和许多其他动物做到这一点。我们对其他动物这些过程了解得越多,就越能更好地理解我们自己。
虽然动物从观察中学习了许多知识,但也有许多知识来自大自然的原始之手……我们将其称为本能,并倾向于将其视为非常不寻常的东西……生活的整个行为都依赖于它……它教导人类避开火焰;就像教导鸟类如此精确地孵化,以及其育雏的整个经济和秩序。
——大卫·休谟,《人类理解研究》(1748/1910)
大脑最重要的本能-情绪系统之一是允许动物搜索、发现和获取生存所需的所有资源的系统。这个探寻系统的唤起产生各种接近行为,但它也以一种特殊的方式让人感觉良好。这不是我们在享用美食时体验到的那种愉悦,也不是之后感受到的满足感。相反,它提供的是当我们期待享用那顿美食时产生的那种兴奋、欣快的期待。当厨房里美味的香气飘到你鼻子里时,你难道不欢迎饥饿的刺痛感吗?与心爱之人的短暂分离同样可以带来特殊的魅力,在重聚的喜悦之前。对性的期待往往比完成时的兴奋更令人激动。即使是对热水澡的期待也可能是一种精致的想象愉悦,尤其是在忍受寒冷天气的寒意时。还有赌博、探索的刺激,更不用说许多审美愉悦了。这种期待渴望的积极感觉(欣快感?),这种探寻冲动,与完成时的愉悦释放完全不同。而且这种感觉作为一种情绪存在于哺乳动物大脑的某些皮层下网络中,远在大脑发展出与世界的旺盛客体关系(如上述描述)之前。最初,它只是一种没有目标的驱动。
如前一章所述,探寻系统传统上被称为”大脑奖励系统”,因为Jim Olds和Peter Milner(1954)发现,大老鼠会过度兴奋地自我刺激这个系统,直到精疲力竭——老鼠强迫性地向这个大脑区域施加微小的电击,仿佛世界上没有什么比这更重要的事情。图2.4显示了这个系统的早期描绘,在解剖学上称为内侧前脑束(MFB),它穿过外侧下丘脑,连接低级脑干和中脑的许多区域到大脑的许多高级区域,一直到内侧额叶皮层。这个庞大的系统向许多其他大脑区域发送连接,因此,如果这个系统在大脑两侧受损,动物就无法再照顾自己。它们看起来极度抑郁(也许是第一个动物抑郁症模型,而研究人员没有意识到这一事实);这些动物通常会在没有密集护理的情况下死亡。
行为神经科学家不习惯给这个生存必需的网络起一个像探寻这样的名字,因为这意味着动物具有一定程度的意向性,但这是因为他们没有过多考虑主要过程情绪力量确实有自己简单思维的可能性——一种使动物在其自然环境中成为主动代理者的原始思维。这些祖先大脑系统自动介导”行动中的意向”,这可能是人类最终”行动意向”的必要前提。行为主义导向的心理学传统称其为食欲性”接近动机系统”,甚至设计了人格测试来测量这种冲动及其一般对立面——“退缩”,或”回避动机系统”(Elliot, 2008)。
英国伟大的人格理论家汉斯·艾森克(Hans Eysenck)最早在一项针对外向性和内向性/神经质的人格测试中概念化了这些维度。他的学生杰弗里·格雷(Jeffrey Gray)以更具神经科学风格的方式,为行为激活系统(Behavioral Activation System)和行为抑制系统(Behavioral Inhibition System)开发了自己的人格测试(综述见Larsen & Augustine, 2008)。其他测试很快跟进,包括积极情感和消极情感量表(著名的PANAS; Watson et al., 1988)。看到科学家们用不同的词汇表达基本相同的意思,真是令人惊叹,这些术语旨在聚焦于一个多面向过程的两个方面。认识到这些测试都没有评估基本的情绪气质后,潘克塞普及其同事着手开发了情感神经科学人格量表(Davis et al., 2003),其中统计上独特的SEEKING(寻求)、CARE(关爱)和PLAY(玩耍)量表共同构成一个积极情感超因子,而FEAR(恐惧)、ANGER(愤怒)和GRIEF/SAD(悲伤)量表则构成一个消极情感超因子。
因此,这个系统与以下方面有关:(i)一般行为激活;(ii)控制”诱因显著性”的”想要”状态(Berridge & Robinson, 1998);(iii)行为的”持续性”(Salamone et al., 2009);(iv)行为模式之间的转换(Oades, 1985; Redgrave et al., 1999);(v)简单的接近行为(Ikemoto, 2010);(vi)可能最为晦涩的是,那些主要迷恋学习理论的人提出的”奖励预测误差”(Schultz & Dickinson, 2000; Schultz, 2010),我们将在后面广泛讨论。遗憾的是,这些术语并未告诉我们SEEKING系统帮助促进的许多不同的食欲行为,它们也没有说明这个系统促进的特定积极情感特征—预期性欣快感—而不是消费的任何”愉悦”。
我们认为SEEKING标签目前是这个初级过程系统的最佳总体名称。已发现这个系统参与大鼠的大量行为,一些发现已扩展到人类(Knutson & Cooper, 2005)。然而,我们在此使用的许多例子实际上并未被神经科学家研究过,因此它们是启发性假设,旨在使我们的理论观点清晰明了。我们预测,当我们描述的所有类型的行为都被研究后,我们将一次又一次地确认SEEKING系统在我们沉迷的每一种积极食欲行为中的作用。
从情感角度来看,该领域一个持续存在的困境是,许多对这些问题感兴趣的科学家(例如,可能最突出的是达马西奥Damasio, 1994, 1999)似乎相信,所有类型的良好感觉都是由我们的感官介导的。也许他们忽略了我们古老的脑内本能情绪行动系统也能产生心智的情感品质。这一重要信息似乎在大多数关于情感感受的科学分析中缺失了。无论如何,证据表明情绪行动系统产生的感受可以完全在大脑内部被触发,尽管每个系统都有某些感官触发点(例如,疼痛会唤起FEAR恐惧)。在学习之后,这些系统通常会被许多其他事件唤起。
尽管有许多感官输入进入维持自我刺激奖励的大脑区域,但我们需要考虑,每个动物的基本情绪,以及它们的核心自我(见Panksepp, 1998b),都是以运动坐标布局的。这种可能性并不排除伴随SEEKING唤起的渴望和欣快体验可以整合来自身体和外部世界的各种感官反馈;它只是表明,有机体的连贯性锚定在原始行动装置上—即位于脑干核心SELF结构中心的内在”行动意图”(见第11章)。无论如何,这个SEEKING系统帮助激励我们所做的几乎每一件充满活力的事情。
当SEEKING系统被唤起时,动物会表现出对世界的强烈、热情的好奇心。例如,大鼠会带着目的感四处移动,用力嗅闻,并停下来调查有趣的角落和缝隙。大鼠经常发出我们在没有特殊设备的情况下听不到的兴奋小声音:50千赫的超声波啁啾声,尤其是在它们玩得开心时特别持久(见第10章关于PLAY玩耍)。这些行为与大鼠在寻找奖励时表现出的行为相同,而不是在享用美食时。人类报告一种急切的期待感,以及一种增强的自我效能感,认为自己可以在世界上让事情发生。人和动物显然喜欢这种感觉,尽管它也可能变得过度。他们会不懈地工作,直到精疲力竭(有时甚至会导致死亡,例如实验室大鼠被允许每天只在它们也被允许自我激活大脑”欣快”系统的同时吃一顿饭)。动物会花费大量精力来获得对这一回路的电刺激或化学刺激(Ikemoto, 2010)。我们将这个关键的动机系统命名为SEEKING-EXPECTANCY(寻求-期待)系统,或简称SEEKING系统。这一名称比经典的”奖励系统”概念更能说明这个系统的整体功能。大脑中有许多情感奖励过程。
行为科学家传统上一直在做我们已经在这里做出的区分,即消费性行为和食欲性行为之间的区别。在动物能够消费奖励之前,它们必须经历食欲阶段:它们必须搜索、找到并占有它们需要的资源。而这不仅仅适用于寻求消费性资源。SEEKING(寻求)系统可能参与所有其他情绪系统的食欲阶段,尽管以下大多数尚未从神经科学角度进行研究。例如,当一个孩子在出去和朋友们在泳池里玩耍之前急切地穿上泳衣时,她的SEEKING系统可能会帮助激发她的准备工作。当我们策划对那些激怒我们的人进行报复时,肯定是SEEKING系统促使我们制定这些计划。因此,一些霸凌者会急切地渴望打架。当充满希望的恋人为重要约会选择完美的餐厅时,他们的SEEKING系统可能正在为浪漫邂逅铺平道路。当你为你关心的人烤蛋糕时,SEEKING系统帮助你预期他们的惊喜和喜悦。当你害怕时,你必须寻求安全。这些体验中存在许多认知差异,但所有这些活动中的预期紧迫感都共享一种共同的积极的想要做和能够做的感觉。同样,从消极的一面来说,当SEEKING系统长期不活跃时,我们会经历一种绝望的抑郁形式,其特征是倦怠和缺乏进取心。
SEEKING唤醒也会让我们在困境中继续前进——当我们饥饿、口渴、寒冷或孤独时。也许我们甚至因此感觉好一些。这是因为SEEKING系统提供积极、热情的情感,至少在某种程度上可以抵消这些负面感受——这种状态我们通常称为绝望。假设一只动物饿了。饥饿感觉很糟糕,但来自SEEKING唤醒的那种令人鼓舞的目标感仍然使动物对其环境保持好奇,并足够乐观地进行集中而充满活力的食物搜寻。换句话说,找到食物的”愉悦”预期和能够这样做的积极感觉提供了一种充满希望的期待感,这将抵消饥饿的负面感觉,如果幸运的话,最终会消除它们。然而,当每个计划都失败时,绝望最终会降临,这就是通往抑郁的大门。
所有令人不快的内稳态失衡状态都会自动使SEEKING系统对奖励(以及预测它们的线索)更加敏感。被称为内感受器(或”需求探测器”)的特化神经细胞,存在于大脑的古老内侧区域以及一些其他身体器官中,测量导致口渴和其他身体需求情感指标的内稳态失衡。例如,当血液水浓度降低时——无论是由于细胞脱水还是血容量减少——特定类型的内感受器会做出反应,从而唤起口渴的感觉。当糖和体脂水平下降时,其他的会跳入行动,促进饥饿感。其他系统促进困倦,在睡眠中我们有由多巴胺驱动的SEEKING冲动所激发的梦。动物也有SEEKING梦吗?蜂鸟每天必须大量进食,否则它们会死亡;进化教会它们每晚进行微型冬眠,以节约早晨搜寻花蜜所需的关键能量。但我们不知道它们是否像我们一样拥有由希望和恐惧激发的梦,而且我们也没有办法找出答案。
一些内部传感器测量性激素的变化,这可以促进LUST(性欲)感受。还有其他传感器监测核心体温。尽管我们还不知道所有涉及的确切机制,但神经科学家已经取得了巨大进展,并开始了解神经肽(neuropeptides)如何向SEEKING系统传递这种特定的内稳态信息,促进行为激活。然后SEEKING唤醒激励动物热情地搜寻它们需要的多种类型的资源。当动物饥饿、口渴或寒冷时,特别是当环境中有可用资源的迹象时,它们的SEEKING系统会超速运转,因为它们觅食、寻水和寻找庇护所。同样,当它们有社交需求时,它们可能会寻找配偶,或者,如果非常年幼,会寻找它们的母亲。
除了响应体内平衡失调外,SEEKING系统还会在动物体验到与更复杂的社交需求相关的负面情绪时被唤起。这些社交需求不像简单的体内平衡需求那样由内感受器来监测。然而,正如我们将在后面章节中学到的,未满足的社交需求,比如对陪伴的需求或玩耍的需求,会造成情感上的痛苦。我们不知道不愉快情绪唤起SEEKING系统的确切机制,但研究表明许多神经肽再次参与其中。例如,心理痛苦和孤独感是由大脑中高水平的促压力皮质激素释放因子(CRF)和内啡肽的缺乏所促成的,内啡肽是大脑自身制造的内源性舒缓阿片类神经肽。当人类(和动物)大脑中有充足的内源性阿片类物质时,他们会体验到积极情绪和舒适感,非常类似于与好友和爱人在一起时的感觉。当这些化学物质水平低,而CRF水平高时,人和动物会感到孤独、痛苦,常常很悲惨。当他们找到陪伴时,这些痛苦的情绪会得到缓解,部分是因为内源性阿片类物质的释放,但也部分是因为大脑内催产素和催乳素活性的提升以及许多尚未确定的分子。最近确定的一种能够促进SEEKING功能的额外分子是胰岛素样生长因子1(IGF-1;Burgdorf等,2010)。
可能仅仅是内源性阿片类物质的缺乏就会唤起SEEKING系统,进而促使人和动物去寻找让他们感觉更好的社交陪伴;但目前关于这类问题的确凿证据仍然很少。我们也知道,看似矛盾的是,我们可以通过将微量阿片类物质直接置于多巴胺细胞附近来强化SEEKING活动,这些剂量可以激发欲望并刺激食欲。也许低剂量的阿片类物质实际上是通过抑制附近的GABA神经元来促进SEEKING的,这些神经元通常会抑制SEEKING冲动(Ikemoto,2010)。还有其他可能性。SEEKING系统也参与缓解其他负面情绪,比如恐惧(Salamone,1994;Blackburn等,1992)。当人和动物处于危险中时,他们的SEEKING系统促使他们寻找安全庇护所。
SEEKING系统既响应贪婪也响应需求。它最初对任何触手可及的奖励都极其敏感(Schultz等,1993)。当某人非常饥饿时,即使是一块干面包皮也可能是一种享受,正如许多囚犯在古拉格集中营和我们有时极其残酷的社会世界的集中营中发现的那样。但即使身体需求得到满足,动物和人类也会被诱人的刺激所吸引。例如,如果一只猴子刚吃饱,当它发现美食——一根香蕉或其他喜爱的食物时,它仍然会变得兴奋。然而,当我们饥饿时,我们会被美食更加吸引。我们哺乳动物同样容易受到各种诱惑的影响。谁能抵挡那额外的一块蛋糕或其他喜爱的食物?当涉及到像酒精、可卡因和海洛因这样的药物时,正是SEEKING系统巩固了我们成瘾的欲望。动物会对与人类完全相同的药物上瘾。一些研究人员认为这种情况发生时动物对此没有感觉。事实上,当我们开发出监测动物感受的方法时,例如通过它们的情绪化发声,我们发现那些声音可以自发地表明动物的感受,突显了成瘾冲动的潜在情感本质(Browning等,2011;Burgdorf等,2001;Panksepp,Burgdorf等,2002;Panksepp,Knutson等,2002)。
在野生动物中,很容易看到SEEKING系统在起作用。资源不易获得,动物必须持续寻找它们才能生存。它们必须狩猎或觅食,寻找水源,找到树枝或挖洞来建造遮蔽巢穴。SEEKING系统促使它们养育幼崽,寻找性伴侣,当动物生活在社会群体中时,还要寻找非性伴侣,形成友谊和社会联盟。然而,在现代人类生活的舒适环境中,SEEKING系统的作用并不那么明显,这在发达国家尤其显著。我们在超市的过道里悠闲地进行”狩猎”。只要水龙头里有水,就不需要主动寻找水源。我们可以轻松获得温暖舒适的家。我们在安排好的聚会上见朋友和寻找爱人。
但即使身体需求得到满足,这个系统仍对诱人的可能性保持警觉。因此,很容易理解这个系统如何在提供如此多诱惑的现代社会中引发各种过度活动。我们倾向于暴饮暴食,在不明智时吸烟,过度饮酒。我们中的许多人是工作狂。药物成瘾泛滥。我们过度渴望查看电子邮件、赌博,并沉溺于不明智的性放纵。简而言之,我们的SEEKING系统可能太容易促使我们沉溺于各种活动,而不会停下来仔细考虑我们在做什么。
尽管这个系统能强有力地响应体内平衡需求、情绪冲动和诱人的诱惑,但当人类和动物不需要特别的资源或没有迫切需要解决的问题时,它会在较低的水平上持续在后台运作。这个系统让动物不断探索环境,以便记住资源的位置。这样,当它们需要食物、水、同伴或安全时,就能做好行动准备。SEEKING系统对人类来说也或多或少持续运作。我们定期扫视周围环境,看商店橱窗,翻阅杂志和目录,浏览网页和回复电子邮件。我们总是在寻找可能需要或想要的东西,或者可能仅仅让我们感兴趣并满足好奇心的东西。我们的SEEKING系统让我们保持与世界互动的一般状态。
在智力不如我们的动物中,SEEKING系统的运作不会掺杂人类特有的预见和战略规划。在人类中,战略思维在SEEKING唤醒中起着重要作用,因为这个系统和我们所有的情绪系统一样,与额叶新皮层有丰富的连接,而额叶新皮层是认知心智大脑(MindBrain)中最高度发达的部分。当SEEKING系统唤醒人类的新皮层时,它激活思维过程——一种虚拟世界——产生复杂的习得行为,这些行为不是本能的,甚至可能是反本能的。
想想正在扑灭火灾的消防员。情况很危险,他们会感到一定程度的恐惧,这会自动唤醒他们的SEEKING系统。在正常情况下,通过FEAR系统的共同激活,SEEKING唤醒会促使消防员寻找逃生方法。然而,由于他们接受过帮助他人和扑灭火灾的训练,SEEKING唤醒会通过激活新皮层的思维和规划能力,激发这些习得的技能。我们已经注意到,当动物饥饿时,它们的SEEKING系统会产生热切寻找食物的冲动。但当消防员的SEEKING系统被唤醒时,它有助于抵消她的恐惧,让她能够专注而有活力地完成工作。她所有的训练、经验和聪明才智——她所有的认知和身体能力——都将用于寻找灭火和帮助人们逃生的方法。
除了促进消防员所从事的那种实用战略思维外,SEEKING系统还能唤醒新皮层纯粹的智力能力。例如,你可能买这本书是因为你在智力上好奇,想了解大脑如何创造情感体验。我们已经确定,新皮层不能提供自己的动机;新皮层是由皮层下情绪系统激活的。正是你皮层下的SEEKING系统帮助激活你的新皮层——你的智力——并促使你做诸如买这本书的事情,如果书很吸引人,还能从书中学习。同样,建筑师、作家、艺术家、政治家和科学家的SEEKING系统促使他们发现新的和更好的方法来解决问题和表达自己。这个系统激发所有人类的创造力——它一直是所有文明的精神引擎。
这绝非小事。它强调了一个事实,即在许多方面,新皮层——我们人类智力的来源——是我们情绪系统的仆人。SEEKING系统推动新皮层寻找满足我们需求和欲望的方法:耕种农场、饲养动物、建造舒适的庇护所、编织保护性衣物。SEEKING系统促使新皮层做让我们感到重要和掌控自己命运的事情;我们试图以让我们更有影响力或更强大的方式操纵社会关系。我们为自己和神灵建造纪念碑,并通过艺术努力表达自己。SEEKING系统促使我们满足对新奇事物的喜好。我们从事揭示自然奥秘的科学研究。SEEKING系统还促使新皮层设计满足我们每一个欲望的方法。我们不仅耕种和挤牛奶;我们还制作巧克力。我们的衣服不仅是为了保护,也是为了美丽和性吸引力。人类伟大而独特的成就,我们巨大新皮层的产物,牢牢植根于这个系统提供的精神能量。
显然,SEEKING-EXPECTANCY系统是一个通用系统,用于获取世界上存在的各种资源,可以说从坚果到知识。简而言之,它参与所有在满足之前的欲求行为;它产生搜寻环境中任何和所有”果实”的冲动;它激发从美味食物到性可能性到政治权力等积极体验的动态渴望;它激励人类和动物通过对抗或逃到安全地带来克服危险;它振奋人类并促使我们投身于创建文明的伟大任务。但在开始时,在出生时,它只是”一个没有目标的驱动”(Panksepp, 1971),打开了与世界互动的大门,从而获得知识。
SEEKING系统由大脑多巴胺驱动,但它远不止产生这一种激活性神经递质那么简单。它是一个复杂的知识和信念生成机器。难怪这个系统仍被称为”大脑奖励系统”。事实上,这是一个古老的大脑系统,使我们和所有其他动物能够获取世界上的所有奖励。这可能就是几乎导致世界在一个世纪内遭遇第二次重大金融萧条的系统——2008年的经济危机,其中自私的贪婪超越了更广泛的人类和社会关切。显然,这个系统需要良好的训练才能减少人类悲剧。它没有内在的道德。它只是一个超高效的起身去获取的系统。人类的认知抱负,无论是善还是恶,都源于其巨大的情感”能量”。
从解剖学上讲,SEEKING系统的路径从腹侧被盖区(VTA)延伸到三个主要目的地:(i)内侧前脑束和外侧下丘脑(MFB-LH),(ii)向上到伏隔核,以及(iii)通过中脑边缘和中脑皮质多巴胺通路到内侧前额叶皮层。解剖结构的总体概述见图3.1。该系统的一些主要神经元,即位于VTA的多巴胺神经元,接收来自大脑其他部分的大量输入。正如我们提到的,该系统也有大量输出到大脑的几个高级区域,特别是伏隔核,这是食欲学习的主要中转站。在某些”低等”哺乳动物如大鼠中,激活该系统的上行多巴胺通路不会投射到额叶皮层区域之外。然而在人类中,该系统延伸得更远,进入集中在大脑后部的感觉-知觉皮层。这与以下事实一致:SEEKING系统在人类中唤起的认知功能在其他动物中没有明确的同源物。
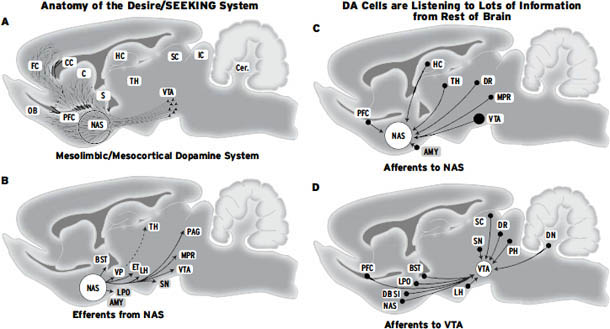
图3.1. 大鼠大脑示意图。A. 位于VTA的A10 DA(多巴胺)神经元的上行投射,支配边缘区域,包括NAS(伏隔核)、中脑边缘DA系统,以及通过中脑皮质DA系统支配皮层区域。B. 来自NAS的主要传出投射。C. 到NAS的传入投射。D. 到VTA的传入投射。缩写—AMY,杏仁核;BST,终纹床核;C,尾状核-壳核;CC,胼胝体;DB,Broca对角带;DN,齿状核;DR,中缝背核;ET,脚间核;FC,额叶皮层;HC,海马;IC,下丘;LH,外侧下丘脑;LPO,外侧视前区;MPR,中脑桥网状核;OB,嗅球;PAG,导水管周围灰质;PFC,前额叶皮层;PN,臂旁核;SC,上丘;SI,无名质;SN,黑质;TH,丘脑;VP,腹侧苍白球;VTA,腹侧被盖区(改编自Ikemoto & Panksepp, 1999)。
在所有哺乳动物中,伏隔核与内侧额叶皮层相互作用,促进简单的食欲学习(和成瘾)。因为SEEKING系统激活额叶新皮层区域,特别是关注即时情感需求的内侧区域,我们能够设计策略来获得生活的恩赐并逃避其陷阱。当体验特别愉快时,我们会记住它们,这为成瘾的可能性奠定了基础。如前所述,该系统的多巴胺部分在人类中比在大多数其他动物中更广泛地延伸到整个皮层。当然,该系统与许多其他大脑区域协同工作(图3.1 B、C、D),包括控制整体唤醒的区域(全局运作的去甲肾上腺素和5-羟色胺系统)以及更具体的大脑注意功能,如由乙酰胆碱、GABA和谷氨酸介导的功能。因为SEEKING系统也参与本书将讨论的所有其他情绪的实施,我们不会在每一章重复这些复杂性,但我们认为读者会理解,对每个系统的讨论都是从每个系统所嵌入的更大脑复杂性中抽象出来的。没有大脑其余部分的帮助,任何情绪系统都无法发挥太大作用。
SEEKING系统主要由神经递质多巴胺(DA)提供能量。DA在刺激该系统中的作用已得到最深入的研究,但还有其他关键化学物质使该系统能够执行其所有功能。神经科学已经积累了关于多巴胺功能的大量分子细节——足以让普通读者头晕目眩。滥用药物,如可卡因或苯丙胺,之所以令人上瘾,是因为它们直接增强多巴胺的作用,从而唤起SEEKING冲动。如果过度刺激,动物的行为会变得刻板,人类会对非常平凡的事物产生强烈兴趣。例如,女性可能会反复整理她们的手提包——把东西拿出来然后再放回去,似乎无休止地,似乎入迷了。如果这种唤起持续太长时间,个体可能会变得多疑,大多数人会发展出偏执倾向。正如我们将看到的,该系统的过度活跃会导致精神疾病,如偏执型精神分裂症。
其他大脑化学物质,最显著的是谷氨酸(glutamate)(Heidbreder et al., 1992; Yeomans et al., 1993),这是大脑的主要兴奋性神经递质,在寻求系统(SEEKING system)的习得功能(学习)中发挥着重要作用。在很大程度上,当伏隔核(nucleus accumbens)将来自内侧前额叶皮层的认知影响与来自寻求系统较低区域的情绪能量整合在一起时,就会发生食欲性学习(appetitive learning)(Kelley, 1999, 2004)。谷氨酸是推动食欲性学习过程的主要大脑化学物质,正如它推动所有其他情绪系统的学习一样。
除了多巴胺和谷氨酸之外,各种神经肽(neuropeptides)也是调节寻求系统的明确化学参与者。例如,神经肽食欲素(orexin)使稳态失衡以及其他情绪系统(如恐惧系统)能够唤起寻求系统。动物通常对获得激活寻求系统的神经肽如神经降压素(neurotensin)充满热情,而它们通常不喜欢使系统失活的化学物质如强啡肽(dynorphin)。这强调了一个事实:人类和动物喜欢寻求唤起的感觉,不喜欢这个系统降得太低的感觉。现在很清楚的是,当这个系统崩溃,强啡肽产生的厌恶感开始占上风时,人们会感到抑郁。研究人员目前正在开发新的抗抑郁药物,可能会减少这一通路上过多强啡肽带来的可怕感觉(Bruchas et al., 2010)。
我们之前提到,只有极少数刺激会固有地(无条件地)唤起大多数情绪系统。老鼠对捕食者的气味、明亮照明的开放空间等有固有的恐惧。其他哺乳动物有不同的固有喜好和厌恶。然而,寻求系统也会被所有新奇事件短暂唤起,这意味着它会被环境中的大量变化在短时间内唤起。当刺激不再新奇(当动物习惯了它)时,寻求系统不再响应。这种现象被称为”习惯化(habituation)“。该系统也固有地响应意外的奖励刺激,如食物的递送(Schultz, 2006)。如果奖励偶尔或时不时地递送,该系统会继续反复响应——也就是说,它会发展出持续的预期冲动(或慢性渴望)。在某些动物中,这可能包括猎物的气味或红色成熟水果的景象。
这个本已庞大的刺激库会随着学习而扩展。假设一个婴儿被挂在婴儿床上方的闪亮悬挂玩具所激发。当这个吸引人的景象移动时,各个部件相互接触并发出叮当声。也许在一个炎热的夏日午后,婴儿坐在厨房的高脚椅上,刚刚吃完午饭。他的母亲给自己倒了一杯冰茶,当婴儿听到玻璃杯中冰块的叮当声时,他变得兴奋起来。也许听起来像悬挂玩具的叮当声。当婴儿第一次看到悬挂玩具时,它是一个新奇的刺激,唤起了他的寻求系统。现在习惯了它,婴儿的寻求系统有些习惯化了。尽管如此,叮当声仍然可以唤起婴儿的寻求系统,尽管不如它是新的时候那么强烈。现在,任何让婴儿想起悬挂玩具的东西,比如母亲杯中冰块的声音,或者甚至当婴儿可能想象这个声音时,都可以唤起他的寻求系统。但我们无法真正在人类身上研究这些问题。对于每个观察到的行为总是有替代解释。例如,也许冰块的声音唤起系统是因为它是一个新奇的声音,而不是因为它唤起了对悬挂玩具声音的某种记忆。
无论如何,各种这样的联结(associations)在一生中发生,导致高度个性化的唤起模式。动物研究实际上可以追踪因果级联,而人类脑成像可以提供类似过程的不太精确的证据。因此,我们有充分的理由相信,强迫性赌博和性冲动是寻求冲动的精妙挑逗者——当一个人越来越兴奋时,伏隔核会越来越亮。似乎生活中所有渴望的兴奋都会唤起这个系统。然而,有些路径导致过度,而另一些则引导人们获得实质性的人生成就。这留给受过良好教育的新皮层(neocortex)来决定追求哪些生活选择。但如果条件反射足够强烈,通常高级心智无法抵抗低级心智想要追求的诱惑。
寻求系统通过消费被渴望的东西而平静下来,但如果满足感不持久,它不会平静太久。当饥饿的动物觅食时,它的寻求系统被唤起,但当它开始进食时,寻求系统变得平静。尽管如此,该系统可以被特殊美味的可能性迅速唤起。然而,当系统受挫时,也许是因为其他小动物得到了美味,愤怒可能会爆发。考虑一下在自动售货机中投币但机器没有履行其交易部分的常见挫折。人们会摇晃有时还会踢这台机器。就神经生理学而言,寻求系统在没有获得满足(没有美味)的情况下关闭,然后这会唤起愤怒系统(RAGE system)。
一个运作良好的SEEKING系统对身体和情绪健康至关重要。然而,当该系统受到过少或过多刺激时,可能会引发情绪障碍,从抑郁症到精神病不等。Oliver Sacks在他的著作《觉醒》(Awakenings)(1973)中描述了帕金森病导致的多巴胺耗竭使SEEKING系统受到刺激不足,患者因此遭受的严重抑郁。左旋多巴(L-dopa)药物在一段时间内纠正了这种化学失衡,产生了显著效果。Sacks引用他的一位患者Leonard L.的话写道:“我感到被拯救了……复活了,重生了。我感受到一种相当于恩典的健康感……我感觉像一个恋爱中的男人。我打破了将我与爱隔绝的障碍。”可悲的是,过量的多巴胺最终过度刺激了这些患者的SEEKING系统,产生了过度的渴望和欲望以及不切实际的命运感——简而言之,就是精神病症状。正如我们将看到的,在这种精神状态下,人们会开始在事件之间看到妄想性的联系;动物也表现出类似的错误归因。
正如已经提到的,当SEEKING系统长期活跃不足时,例如在反复挫折之后或从安非他明和可卡因成瘾中戒断期间,就会出现抑郁情绪。另一方面,精神分裂症、躁狂症和精神病妄想出现在SEEKING光谱的另一端,反映了当系统被多巴胺严重过度刺激时的过度心理倾向(Grace, 1991)。安非他明和可卡因等滥用药物是SEEKING系统非常有效的刺激剂,因为它们增加了突触间隙(神经元之间的交流通道)中多巴胺的可用性。这类药物很容易被滥用,它们使SEEKING冲动变得超敏,使人们对成瘾药物更加敏感。动物对其他奖励也变得更加敏感,从美味食物到性接触(Nocjar & Panksepp, 2002)。精神科医生很清楚,这类药物如果服用时间过长且剂量过高,最终会导致精神病症状——对任何人都是如此。有些人很快屈服;其他人恶化得更慢。但每个服用过量这些药物的人最终都会陷入精神病性、偏执性思维(Snyder, 1972)。然后,在药物戒断期间,抑郁会占据主导。
我们已经提到,SEEKING系统在唤醒内侧额叶皮层的认知区域方面特别有效。新皮层的功能之一是其产生因果概念的能力。当它受到过度刺激时,负责构建”工作记忆”(working memory)(见第6章)的额叶皮层会产生大量关于世界如何组织的新想法。它经常会激发某人在只有相关性或根本没有有意义联系的地方看到因果关系和其他有意义的联系。当这种情况发生时,思维失控,导致大量且经常错误的结论。现在,思维成为妄想滋生的肥沃土壤。SEEKING唤醒的典型特征——增强的自我感——同样可能呈现出不切实际的比例,导致夸大的精神病妄想。
例如,一个精神分裂症患者可能怀有妄想信念,认为他的行为,比如打破一面心爱的镜子,导致了一个重要的世界事件——比如2001年9月11日世贸中心双塔的爆炸。这将构成因果关系的妄想信念,因为患者的个人行为并没有在更大的世界舞台上引起某些事情。这其中也包含夸大妄想的成分,因为患者相信他有能力导致这些重要事件发生。这些精神病幻想是由严重过度唤醒的SEEKING系统产生的。值得注意的是,压力可以提高额叶皮层中的多巴胺活性。这可能解释了严重压力如何帮助促进偏执性、精神分裂症思维模式。事实上,有些人已经设想了这种思维模式与梦境之间的关系(Panksepp, 1998a; Solms, 2002),最近的研究已经证实,SEEKING系统中的多巴胺神经元在REM睡眠期间以非常高的速率放电(Dahan et al., 2007)。因此,合理的结论是,大脑中丰富的多巴胺活性在梦境和精神分裂症中都会发生(Léna et al., 2005; Panksepp, 1998a; Solms, 2000)。
多巴胺是唤醒SEEKING系统的主要化学物质——虽然它不是唯一的,但它肯定是我们最了解的。多巴胺通过在突触处以全局方式释放并与接收神经元上称为受体(receptor)的分子结合来唤醒该系统(受体有五种主要类型,分为两个家族,即D1和D2,我们在这里只考虑其中之一:D2受体,它在精神分裂症等精神疾病中特别重要)。结合以钥匙和锁孔的方式发生,其中多巴胺充当钥匙,受体充当锁孔。除了多巴胺,大量其他化学物质(即神经肽和其他神经递质)也可以充当它们各自特定受体的钥匙。
通常每种脑部传递物质化学物质都可以与多个不同的受体结合——每种化学物质都有不止一个可以”对话”的受体。另一方面,受体通常更具专一性;它们只能”倾听”并与特定的传递物质化学物质结合。一个能够嵌入受体”钥匙孔”但无法打开(或激活)它的化学钥匙被称为受体阻滞剂。当受体被阻断时,通常与其结合的化学物质无法这样做,这种脑部化学物质的活性因此降低。因此,如果使用多巴胺阻滞剂(许多此类药物是抗精神病药物),那么在突触处释放的多巴胺就无法再与SEEKING系统中的受体结合,该系统变得唤醒不足,导致抑郁症状,如上述所描述的那些。
研究人员发现,多巴胺在其一种受体(即D2型)上的过度活动会导致(或至少与之相关)某些精神分裂症状。几乎所有治疗精神分裂症状的药物,即那些能够平息妄想和幻觉的药物,都会在D2受体处阻断多巴胺活动。如果上述打碎镜子的患者接受抗精神病D2阻滞剂治疗,他妄想的认知方面不会完全消失,但妄想的动机力量会显著减弱。他可能仍然认为自己与那些灾难性事件有关,但这些想法不再具有同样强烈的确信程度。换句话说,抗精神病药物通常会降低妄想的强度,但不会改变其内容。这就是为什么谈话疗法有时也有助于帮助患者重构其妄想认知。就这位患者而言,如果他的妄想源于过度的愤怒,那么理解是什么使他如此容易愤怒可能会有所帮助。阻断多巴胺信号的抗精神病药物也会抑制动物探索环境并因此获取新信息的倾向。探索倾向是SEEKING系统的正常表达。妄想位于SEEKING连续体的病理性远端。
当SEEKING系统被较轻度地过度刺激时,它会产生附属行为,这些行为是强迫性的,但通常没有明显的外在目的。在实验室条件下,例如当非常饥饿的动物定期收到少量食物时,人们会看到附属行为。它们收到的少量食物不足以让它们满足,而且它们无法自己获取更多。由于这些动物处于持续的饥饿状态,它们的SEEKING系统持续处于过度唤醒状态。当它们等待下一次少量食物投放时,这些动物通常会从事附属行为。例如,一只饥饿的实验室大鼠可能会在跑轮中过度奔跑。另一只大鼠可能会撕碎纸张、啃咬木头或饮用大量的水。这些行为与它们的身体需求无关,因此被称为附属行为。在日常生活中也可以看到附属行为。非常饥饿的人倾向于来回踱步。踱步是一种附属行为,它既不能滋养身体,也不能获取食物。事实上,如果它消耗稀缺的能量,可能会适得其反。
附属行为通常是重复性的,并且看起来是仪式化的。行为主义创始人之一B. F. Skinner指出,饥饿的鸽子在接受小份食物之间的长时间间隔期间,会进行重复且可预测的昂首阔步、拍打翅膀的”舞蹈”(Skinner, 1948)。它们不是在刚收到一小块食物后表演舞蹈,也不是在非测试期间表演。相反,鸽子在等待下一批食物时跳舞,通常处于极度饥饿状态——这种状态无条件地唤醒SEEKING系统。我们并不是说鸽子在认知上”认为”它们可以通过跳舞让食物出现。相反,似乎当SEEKING系统被过度刺激时,它会自动促进重复性和仪式化的行为。这些附属行为通过多巴胺阻滞剂以及外侧下丘脑的损伤而显著减少,这些操作会使SEEKING系统失活或受损(Wayner et al., 1981)。
更难理解的是,为什么动物会表现出一种重复的附加行为而不是另一种。例如,为什么一个饥饿的人会在地板上踱步,而另一个人吹口哨,还有一个人用拳头捶打手掌?在对动物的研究中,所表现出的行为类型似乎是被研究动物的特定属性——也就是说,它是其个性的一个属性。因此,性格更具攻击性的人可能会捶拳头,而更顺从的人会吹口哨。或者,附加行为可能看起来更有目的性——它们针对通常预示奖励的刺激。例如,定期获得小块食物的饥饿老鼠会开始啃咬投放食物的食槽,尽管这丝毫不会影响食物投放的速度。这几乎看起来好像某些类型的行为给动物一种专注的目标感。换句话说,仪式化的附加行为似乎是以一种让动物感觉它正在做一些有成效的事情的方式形成的,即使它并没有。类似地,服用大剂量可卡因和安非他明的人和动物,这两者都会强烈刺激SEEKING系统(探寻系统),表现出看似无止境的重复行为。如前所述,当人类有如此强烈的刺激时,他们经常报告说,做一些平凡的事情,比如翻找手提包,突然变得非常有趣。
这种表现出重复和仪式化行为的倾向可能具有适应性价值。学习一项新技能需要重复,有时甚至到成为一种仪式的程度。当体操运动员学习如何完成双后空翻时,她可能会准确地走四步,以特定的方式低头,总是用同一只脚起跳,等等。我们有许多涉及重复和仪式的习惯。我们每晚把钥匙放在同一个挂钩上,我们以特定的方式叠衣服。即使当我们洗澡时,我们也倾向于以某种顺序清洗不同的身体部位。看起来SEEKING唤醒有助于产生这些习惯。然而,一旦一种行为成为习惯性的,它就被储存在多巴胺控制的大脑区域,比如伏隔核正上方的背侧纹状体(例如,尾状核),其唤醒由VTA外侧的黑质纹状体多巴胺系统控制。这些大脑区域的刺激奖励性要低得多,因为习惯就是习惯。许多习惯是无意识完成的。在这种情况下,当以前通过SEEKING冲动建立的令人兴奋的行为变得常规时,人们不再需要变得情绪激动。因此,动物对激活那些更新的多巴胺系统不会表现出太多自我刺激行为也就不足为奇了。
自动塑造是指一种实验室现象,当动物非常饥饿时(这也意味着其SEEKING系统高度唤醒),并且当动物还暴露于一个短暂的、无关的刺激时,例如,在投放食物之前点亮食物托盘上方的一个按键,这种现象会逐渐出现(Brown & Jenkins, 1968)。这种预测性刺激似乎与动物获得奖励有因果关系。一个冷静的哲学家可能只会决定耐心等待每次奖励到来,而不会变得充满渴望、满怀期待,从而开始与预测食物的刺激互动。这样的行为无论如何都无法帮助缓解饥饿;然而,动物会逐渐开始与这些刺激互动,几乎就好像它们相信这种互动会获得奖励。在反复暴露于这样的配对之后,动物,在这种情况下是鸽子,开始啄在食物投放前会被点亮的按键。鸽子会在实验者停止投放食物很久之后仍然坚持这种活动,尽管啄击什么也完成不了。自动塑造现在已经在所有被研究的哺乳动物物种中被观察到。这显然是一种SEEKING行为,因为多巴胺会阻断自动塑造的效应(Phillips et al., 1981)。这对那些认为动物会明智行事而非情绪化行事的人来说是一个挑战。
对于我们这些从因果关系角度思考的智慧头脑来说,似乎被自动塑造的鸽子在啄按键和食物投放之间建立了一种有用但虚幻的心理联系。也许确实如此。然而,大多数行为研究者怀疑鸽子是否足够聪明来做出这样的心理跳跃。那么是什么导致了这种行为?这可能只是一个盲目学习的问题。也许鸽子并不”认为”啄按键会确保食物投放,就像斯金纳的跳舞鸽子不”认为”它们的舞蹈会获得食物一样。当然,我们永远不会知道,因为我们无法了解其他哺乳动物的想法,更不用说鸟类了(但参见Clayton et al., 2003),至少不像我们能够衡量它们的情绪那样清楚。那么,我们该如何理解这种行为?为什么鸽子会啄按键?嗯,也许它已经被条件反射训练得过度情绪化,而产生迷信行为是在一个非常无聊的环境中度过时间的好方法,就像任何其他方式一样,特别是当实验者不时用食物碎片诱惑它时,这些奖励相当一致地由一个线索预测。
当SEEKING系统被唤起时,动物会对它们的环境产生好奇心。似乎当饥饿的鸽子看到发光的按键时,它的好奇心被唤起,并通过啄击来探索按键。换句话说,SEEKING唤起使人和动物注意到并检查任何可能帮助它们理解世界的刺激。动物不需要”思考”外部刺激与食物传递之间可能存在因果联系。条件性SEEKING唤起确保它们会以有规律的方式对环境产生好奇。这种好奇心是适应性的,因为有时这些外部刺激确实是资源的线索。事实上,在现实世界中,这种”洞察”可能有一半的机会是有效的。例如,如果公园里的一只饥饿鸽子碰巧注意到并调查地上的一些闪亮纸张,它可能会找到美味的薯片残渣。此后,任何闪亮纸张都将作为可能预测食物的线索,看到它将唤起鸽子的SEEKING系统,使其集中接近并与这种刺激互动。
自我塑造(autoshaping)和附加行为(adjunctive behaviors)在严格的实验室条件下是分别发生的。然而,在现实生活中,自我塑造和附加行为通常是并行的。动物进行重复性附加行为,通常使用外部物体——条件刺激——来执行附加行为。例如,自我塑造实验中的鸽子重复啄击按键,公园里的鸽子可能以重复的方式啄击薯片包装。人类也表现出附加行为和自我塑造的组合。假设你的经理一直专横且不公正,唤起了你的RAGE(愤怒)系统。你想和他摊牌,但他推迟到下周,你的愤怒不得不暂时搁置。那天晚上晚饭后,你读报纸,希望从恼怒的专注中分散注意力,你注意到填字游戏,这是你通常会忽略的东西。然而今晚,你尝试了一下,变得异常投入,熬夜到了平常就寝时间之后。当你做填字游戏时,你感觉好些了,甚至可能享受这个活动。然而,一旦你把报纸放在一边,你可能又会想到你的老板并感到愤怒。
从神经科学的角度来说,你的RAGE系统被唤起是因为你的老板让你难堪。据我们所知,一些SEEKING唤起最初伴随所有类型的情绪唤起,在这种情况下,它可能会促使你计划如何接近你老板的策略。然而,因为你的老板一直不可接近,你的困境类似于自我塑造实验中的鸽子。鸽子想通过进食来满足饥饿,而你想通过给老板一番教训来满足你的RAGE。你们两个都没有办法做你们想做的事。所以你的SEEKING系统和鸽子的SEEKING系统在没有有用活动可能性的情况下被唤起。在这些条件下,你和鸽子对外部刺激执行附加行为。鸽子啄击按键,你热切地做填字游戏。也许这有点牵强,但希望观点是清楚的。如果我们想到配偶而不是无生命的填字游戏,会更有意义且与临床更相关。人们可能很容易把愤怒发泄在错误的人身上。我们喜欢有一种控制世界的感觉,即使我们并没有控制。这可能是那么多人祈祷的原因之一吗?或者为什么他们会错误地将情绪发泄在”无辜旁观者”身上?
在本章后面,当我们讨论条件学习时,我们将再次考虑自我塑造和附加行为如何在提供学习发生所必需的环境中发挥重要作用。自我塑造的倾向确保人和动物注意到似乎有因果关系的外部刺激。这是条件学习的必要先决条件。附加行为的倾向使人和动物学会如何执行有效的重复性行为,这些行为通常也会在动物被条件化时出现。自我塑造和附加行为都是SEEKING唤起的表现,两者都可能是条件学习在现实世界中发生方式的基础。
这其中还有一个对科学也至关重要的方面——归纳法(induction)在产生可检验假设中的作用。归纳逻辑不过是沿着相关事件看到关系,以及这种相关性暗示因果关系的”洞察”。当然,这导致了实验,在实验中独立操纵关键相关变量,以查看是否可以证明因果关系。通过这种方式,避免了将相关性视为因果关系来源的许多潜在缺陷,而自我塑造正是基于此。预测和可检验性使科学免于归纳思维可能导致的许多错误线索——从表明地球是宇宙中心的观察到潜在的祈祷改变世界物理事件的力量。这种批判性思维模式将科学从看似无休止的错误信念循环中拯救出来,这些错误信念源于不加批判地按表面价值接受表面观察,这经常是人类思维和文化的特征。
我们已经看到,SEEKING 唤起可以产生持续的仪式化行为,如鸽子在可预测奖励之间的舞蹈或自塑的啄键行为。SEEKING 系统不思考个人事务,但新皮层会,尤其是与该系统相连的内侧额叶皮层。当人们反复思考时,这个大脑区域通常会被激活(Northoff et al., 2010)。人类拥有巨大的新皮层,新皮层有能力从因果关系的角度解释和理解事件。想象一个在科学知识有限的时代遭受干旱的部落民族。在他们的挫折中,人们可能会进行仪式化和附加的情绪行为。他们可能会持续走动,时而踢腿和呼喊,以类似鸽子舞蹈的方式踢起干燥的地面。最终,雨水会来临。注意到他们附加的跺脚行为与降雨之间的相关性,他们可能会相信存在因果关系,这为创造祈雨舞提供了动机,希望促成未来的降雨。此后,他们可能会定期使用舞蹈仪式——一种祈祷形式——作为一种文化认可的努力来产生降雨。
我们现代西方世界的大多数人认为这是一种妄想的思维方式。但我们中的许多人在痛苦时期有祈祷的倾向。有些人祈祷时并不真的相信这会有帮助。然而,这似乎让他们感觉好些,因为他们正在采取某种行动。因为人们通常足够聪明,知道什么时候无法掌控自己的命运,这种行动往往采取向更高力量——上帝——发出言语呼吁的形式,而上帝能够控制命运。附加行为似乎让人和动物感觉更好,因为它们提供了一种幻觉,即他们是有效的行动者——这也是 SEEKING 唤起的一个特征。有些人明确地请求上帝的力量,例如,在看不到停车位时找到一个停车位——有时它”奏效”了!这是否可以部分解释为什么祈祷是如此受欢迎的活动,尤其是在压力时期?祈祷是否可能是一种附加行为,让人类产生一种幻觉,认为他们能够以某种方式神奇地改变自己的命运?
我们也可以想象自塑如何参与宗教象征的创造。假设一个部落的酋长在干旱期间漫无目的地在一块木头上雕刻。当雨水来临时,有人注意到这种随意的雕刻像狼的脸。这块木头可能会吸引部落长老的注意,就像点亮的圆盘吸引了鸽子的注意一样。它将是一个新颖而重要的物体,他们的 SEEKING 系统会专注于它。因为他们巨大的新皮层能够从因果关系的角度思考并编造叙事,他们可能会认为狼拥有带来降雨的超自然力量。然后他们可能会在木头上雕刻狼脸,并将它们用作宗教象征,在困难时期向它们祈祷。当然,我们只是在这里想象这种情况。然而,如果祈祷可以被视为一种附加行为,如果自塑在宗教象征的创造中发挥作用,SEEKING 系统可能会在很大程度上解释宗教信仰的神经根源。在这方面,宗教信仰是许多精神病疾病的核心特征也许并非偶然。
但情感生活还有许多其他方面,这些触角深入宗教传统。我们同意 Thandeka (2009) 的观点,认为人类宗教背后的一个驱动力是我们的情感本质,尤其是我们对养育和理解的迫切需求,通过社区抵御悲伤,并且常常渴望寻求更高的善。我们将在关于 PANIC/GRIEF 系统的章节中再次讨论这个革命性主题。
如前所述,SEEKING 系统最初由 Olds 和 Milner 于 1953 年在加拿大麦吉尔大学研究,尽管他们并没有用这个名字称呼该系统。在寻找其他东西时(即人工诱导的大脑唤起/注意力如何促进学习),他们偶然发现了这样一个现象:动物会为了接受大脑特定部位的微小电击而工作。有时,Jim Olds (1922-1976) 称这为”快乐系统”。但其他研究人员相当拘谨,至少直到 1980 年代,当自我刺激研究的宏大探索时代结束时,大多数研究人员开始专注于这个复杂系统的多巴胺成分。
从那时起,几乎每个人都称其为”大脑奖赏”甚至”强化系统”。但人们不得不怀疑,当奥尔兹在撰写他的最后一本书时(Olds, 1977),他已经意识到这个系统远不止于创造愉悦那么简单。他开始研究食欲驱动的经典条件反射,通过将声音与向饥饿大鼠投喂食物配对,并监测整个大脑的神经元活动。他发现大脑中许多许多地方都学会了预期即将到来的食物,但最快的神经元条件反射,以及最早表明动物正在预期食物的信号,来自我们这里称之为SEEKING系统的MFB-LH走廊沿线的神经元。细胞的放电通常预测即将到来的奖赏,但奥尔兹似乎从未下定决心,他实际发现的是热切期待奖赏的大脑系统,而不仅仅是记录消费奖赏所带来的愉悦。在奥尔兹英年早逝前几年,潘克塞普在飞往欧洲的航班上与他讨论了EXPECTANCY/SEEKING假说,他很感兴趣,注意到他的电生理学工作与这一想法是一致的。
奥尔兹和米尔纳对”大脑奖赏”的发现无疑是二十世纪最伟大的神经科学发现之一,最终导致揭示了学习和成瘾的神经基础的研究。动物会通过MFB-LH刺激学习各种各样的东西,从按压杠杆到沿着迷宫中的特定路径奔跑,这些正式称为操作性条件反射和工具性条件反射。他们称这种效应为自我刺激,因为动物扮演积极角色(它们工作)以接受”快乐”的电击——人们甚至可能认为他们发现了一种精神自慰的形式。动物刺激它们大脑中那些在进化上被设计用来获取其他好处的区域。毕竟,一个自慰的人真正想要的是一段性爱关系,但由于各种原因,最终独自满足了他或她的需求,这与成瘾非常相似(Zellner et al., 2011)。现在我们知道,这个通用的SEEKING系统对于各种成瘾都至关重要,从可卡因和吗啡等药物,到酒精和尼古丁依赖,甚至性成瘾(Wise & Rompre, 1989; Robinson & Berridge, 1993)。该系统也是各种创造性活动背后的驱动力(Reuter et al., 2005)。
仍然存在一个大问题。大多数”继承”了这个迷人大脑系统研究的年轻研究者几乎不质疑前几代人传给他们的”奖赏”和”强化”等单一概念,就好像它们是单一现象一样。事实上,“强化”这个概念可能只是我们无知的一个概括性术语——也许是行为科学的”燃素”2——它只是训练动物的一个方便的程序。然而,作为一个大脑过程,这只不过是掩盖(和隐藏)了一座无知之山。在这个过程中,这个”大脑奖赏系统”的大多数研究者都未能考虑到当SEEKING系统被人为唤起时,动物自发表现出的实际”自然”行为模式,即特征性的探索活动。
动物会自我刺激大脑的许多区域,主要是隔膜区和LH区,通过它们流经包含上行多巴胺系统以及许多其他神经网络的MFB。我们已经知道,动物对MFB-LH和隔膜区的自我刺激体验是完全不同的,尽管沿着MFB-LH的刺激点对动物来说往往感觉相同,因为它们很难区分这条通路上的两个远距离点(Stutz et al., 1974)。因为像这样的辨别工作做得很少,我们必须假设许多其他介导自我刺激的大脑部位也会产生不同类型的奖赏。无论如何,大脑包含许多奖赏系统。
现在已经有好几代科学家对MFB-LH刺激所产生的奖赏类型”推卸责任”,我们可能不必重复,“大脑奖赏系统”的全局概念相当偏离了目标,尽管那些了解得更清楚的人继续使用这个术语(Haber & Knutson, 2010)。这个描述根本无法捕捉到这种大脑刺激在动物身上引发的自然行为模式,而且电生理学一致表明,该系统的设计是首先对新发现的奖赏感到兴奋,然后如果有更多即将到来,就会迅速预期它们。底线是,获得MFB-LH奖赏的动物的行为表现并不像是在享用美味佳肴并体验感官-情感奖赏。刺激隔膜区产生的行为更接近这种情况。
当动物自我刺激外侧下丘脑时,它们会以一种疯狂的方式进行。它们疯狂地按压杠杆,鼻子”每分钟嗅个不停”,几乎就像在试图看清杠杆背后是什么——去探索它——而且它们通常会比获得所有”奖励性”电击所需的工作量要努力得多。相比之下,动物以一种非常不同的行为”态度”自我刺激隔区——它们以有条不紊的节奏工作,通常每次电击按压一次杠杆,并且没有激动表现。从各种测量来看,隔区刺激似乎并不比外侧下丘脑刺激的愉悦感更少,而且根据人类的自我报告,隔区刺激实际上确实会引发愉悦感(Heath, 1996)。那么为什么动物会如此过度努力地工作来获得外侧下丘脑刺激呢?最合理的假设是,SEEKING系统诱发了一种强烈的心理和行为激活——这种唤起是动物在获得即将到来的奖励之前所表现出的,因为它们体验到某种欣快的热情。想想一只饥饿的狗在你端出食物碗时上蹿下跳,有时还转圈的样子。
然而,在发现自我刺激的早期,行为概念作为讨论动物行为的唯一有意义的科学方式占据主导地位。即使是偏好在野外研究动物自然行为的动物行为学家,也将他们的分析限制在对行为的准确描述上,没有任何心理构念的暗示,情绪问题也很少被讨论。诺贝尔奖获得者、动物行为学家尼科·廷伯根指出,由于”主观现象无法在动物身上客观观察到,因此声称或否认它们的存在都是徒劳的”(1951, p. 5)。对于讨论大脑主观方面存在一种禁忌(Wallace, 2000)。因此,动物心智在很大程度上被科学家忽视也就不足为奇了,至少直到唐纳德·格里芬(1984, 2001)开始有力地再次谈论动物意识的可能性——这种探讨在十九世纪后期相当流行,也许过于流行(例如,Lindsay, 1880),特别是达尔文的门徒乔治·罗曼斯(1882)的研究。
当奥尔兹和米尔纳发现自我刺激现象时,行为主义正处于顶峰。行为主义运动最大的成就是发现,当奖励(通常以食物或饮料的形式)在特定时间按照特定程序(即各种奖励传递程序,称为”强化程序”)传递时,可以诱导动物以特定方式工作(它们会表现出操作性/工具性行为),并呈现高度可预测的模式。例如,在一种程序下,当食物在固定次数的反应后传递时(固定比率程序,有点像劈柴),动物会尽可能快地按压杠杆,吃掉奖励,然后放松一会儿,再开始另一轮最快速度的操作性行为。当奖励以不可预测的方式传递时——在不同次数的操作性杠杆按压后(变动比率程序)——它们的工作速度会慢一些但保持稳定的节奏。当奖励以固定的时间间隔传递,而不管操作性行为的次数(固定间隔程序)时,动物在接收到奖励后会缓慢按压杠杆,并随着奖励即将再次传递的时间临近而越来越快地按压(看起来像一条期待递增的曲线)。如果奖励在各种不可预测的时间传递,而不管动物按压杠杆的频率(变动间隔程序),动物的工作速度相对较慢但相当稳定。如果难以想象这些文字描述,请参见图3.2中动物在这些不同程序下工作时的”累积记录”(Panksepp, 1998a, p. 22)。
图3.2. 在固定间隔程序下(每20秒给予一次刺激),给予大鼠半秒钟的免费奖励性外侧下丘脑电刺激后,大鼠自发产生的表示积极情感的50-kHz超声波发声。在仅适度接触这种免费大脑奖励模式后,动物开始表现出预期曲线,这是动物在固定间隔”强化程序”下为获得传统奖励(如食物)而工作的特征。在嗅探行为的测量中也自发获得了非常相似的模式,这反映了由潜在SEEKING系统介导的本能探索反应(数据改编自Burgdorf et al., 2000, Figure 1, p. 321)。
使用强化或奖励程序(比上述描述的基本程序复杂得多的程序)会在所有被测试的动物(包括人类)中产生高度特征性和可预测的行为模式。这种一致性使行为科学家相信,他们正在揭示关于人类和动物在简单学习情境中如何对奖励做出反应的规律。当然,作为激进的行为主义者,他们对情绪和动机的更深层神经本质不感兴趣。相反,他们只考虑刺激和反应,以及与奖励、惩罚和一个称为”强化”的过程相关联,这个过程被认为将它们全部粘合在一起。奖励刺激通常是按特定程序传递的食物或水,而反应则是动物的模式化行为。然而,食物并不是唯一的奖励——雄性动物会为获得性交机会而工作,母鼠会为接触幼鼠而工作,等等。
这些行为发现并没有被那些经营和设计赌场的人所忽视。他们对老虎机进行编程,使其按照最终能最有效地让顾客交出辛苦赚来的现金的模式来提供现金奖励(即,可变比率强化计划表)!当然,从长远来看,赌场总是赢家。事实上,当使用现代脑成像技术监测时,这种贪财活动和心态非常有效地点亮了大脑的”奖励中心”(Knutson & Cooper, 2005)。
Olds和Milner注意到一个事实:当LH刺激按照模仿上述食物递送计划表的方式施加时(例如,固定比率、可变比率),动物工作时表现出的模式几乎与它们为获得食物而工作时表现出的可预测模式相同。不同之处在于,动物为LH刺激奖励而工作时,从不像饥饿的动物为食物奖励工作时那样持久。例如,在固定比率计划表下,老鼠可以轻松地为每一小块食物按压杠杆数百次,但为了大脑奖励,它们很少会超过这些水平的十分之一。如果实验者停止给予动物LH刺激以回报它按压杠杆,按压行为也会很快减弱并停止。动物通常会转而进行放松的自我护理活动,如梳理毛发。自我梳理是动物在进食或性行为后会非常积极地进行的活动——在这些时刻,SEEKING系统相对平静。因此,当实验者停止给予动物大脑奖励时,其SEEKING系统会相对快速地停止活动。然而,如果停止给饥饿的动物食物颗粒,动物会继续按压杠杆更长时间。这是因为动物处于稳态失衡状态,这会自动使SEEKING系统敏感化,促使动物继续积极地按压杠杆。Panksepp发表的研究(Panksepp & Trowill, 1967a, 1967b)发现,非饥饿状态的动物为获得非常高激励的奖励(即,直接注入口中的巧克力牛奶)工作时,往往表现得像自我刺激的动物,这表明缺乏任何身体需求可能与自我刺激动物的异常行为有关。
当然,Olds和Milner必须在当时行为主义者使用的有限概念框架内思考自我刺激现象。这一发现足够令人兴奋,或许是二十世纪最重要的心理学发现,但没有动力去思考像SEEKING-EXPECTANCY系统这样的激进想法,最终研究者很少再关注自我刺激动物与那些为常规奖励而工作的动物之间的差异。大多数研究者认为,自我刺激在某种程度上必然反映了从常规奖励中获得的快乐,也可能反映了导致动物寻找奖励的稳态失衡。因为有不同的方式来恢复稳态(进食、饮水,或通过刺激MFB-LH可以引发的许多行为),各种研究者还假设LH必须包含每种消费活动的子系统。一个子系统会激发进食,可能通过诱发短暂的饥饿感然后是进食的满足感;另一个会激发饮水;第三个会激发性满足;等等。然而,大量实验表明这些假设并不正确。
如果LH是大脑中记录消费快感的部分,那么当动物体验到消费世界上美好事物的愉悦时,它就会被激活。那里的神经元会在动物进食、饮水、交配等时放电。然而,实验数据并不支持这一观点。LH中的神经元通常在动物寻找食物时活跃,但当动物找到食物并开始进食时,这些神经元会立即停止活动(Hamburg, 1971)。其他实验也得出了类似的结果,表明与LH有强烈连接的大脑结构(构成SEEKING系统其他部分的结构)对奖励的预期而非奖励本身做出反应(Blackburn et al., 1992; Fibiger & Phillips, 1986; Schultz & Romo, 1990)。因此,在现实世界中,即在没有人工脑刺激的情况下,在动物正在消费奖励的那些精确时刻,SEEKING系统似乎并不特别活跃。相反,SEEKING系统通常在动物获得它们所期待的奖励之前最为活跃。事实上,正如我们所指出的,当动物开始进食时,MFB-LH中的神经元倾向于停止活动。
然而,在消费阶段确实会释放一些多巴胺,一些研究者认为这意味着LH激活是进食、饮水和其他活动快感的神经相关物。但对这一事实有一个更合理的解释。多巴胺的释放可能是因为当我们吃东西时,存在一种预期和消费的衔接模式。如果你饿了并坐下来吃汉堡,当你开始咀嚼第一口食物时,你LH中的神经元会停止放电。然而,当你吞咽时,你开始期待下一口。在这段短暂的预期期间,你SEEKING系统中的细胞再次开始放电,多巴胺被释放。即使在你吃饱之后,一想到加冰淇淋的苹果派,LH中的细胞可能会再次放电。
因此,合理地推测,即使你正在享用美食的过程中,多巴胺释放和LH唤醒也会以循环模式发生。然而,作为一般规律,LH中的许多神经细胞以及构成SEEKING系统的相关结构,通常在获得满足之前比在获得满足期间更活跃地放电。这些数据支持这样一种可能性:你的SEEKING系统在你期待汉堡时分泌的多巴胺,比你实际带着满足感咀嚼它时要多。此外,我们必须记住,多巴胺只是构成SEEKING系统的复杂神经网络的一部分。
但还有其他令人困惑的现象需要解释。为什么这种自由给予的脑刺激(无需工作)会产生各种消费性行为——进食、饮水、啃木头、交配等等?如果脑刺激产生的是从这些行为中获得的满足感或奖励,那么这是相当令人困惑的行为。但这个普遍观察到的事实引出了另一个合理的猜测:MFB-LH可能包含与每种消费性活动相对应的特定神经子回路。但当这一假设被测试时,结果证明是错误的。如果LH包含所有这些不同的子系统,那么当在同一只动物体内移动电极穿过LH(即游走刺激探针)时,不同的子系统应该被唤醒,动物会首先表现出一种消费性行为,比如饮水,然后如果探针再向下移动一点,动物会开始另一种行为,比如进食或交配。但这并不是实际发生的情况(Wise, 1971)。当使用这样的”游走”电极时,动物会坚持它碰巧表现出的第一种行为,然后无论你把电极放在”活跃区域”的哪里,它都会继续表现出这种行为。如果动物在进食,当电极在整个LH中移动时,它会继续进食。此外,动物有时会坚持那些非消费性的活动。有时它们啃木头,叼着自己的尾巴,收集自己的幼崽,强迫性地啃咬自己的粪便等等。而且动物会以非常相似的方式自我刺激所有这些脑区。因此,研究人员逐渐发现,LH的”奖励回路”并没有针对许多不同消费性活动的独立神经回路。相反,该系统准备好对任何维持生存的活动做出反应。
得出这个系统服务于某种一般行为功能的最重要研究,是艾略特·瓦伦斯坦(Elliot Valenstein)及其同事的研究,他们发现了动物在该系统受到刺激时表现出的各种食欲行为的一些非常显著的特性。这些行为非常灵活且可互换。如果一只动物优先大量进食,而不是饮水或啃咬可用的木块,然后通宵持续刺激这只动物,但现在让动物无法接触任何食物(它们最初偏好的”目标物体”),第二天早上,动物要么在饮水,要么在啃木头,就像前一天它们进食一样热切(Valenstein, Cox等,1970)。更令人惊讶的是,当瓦伦斯坦及其同事归还食物时,动物坚持了它们新发现的行为。他们还发现了许多其他令人困惑的行为模式。例如,如果动物在脑刺激期间首先开始从吸管中喝水,而研究人员只是把水放在盘子里,那么动物开始进食或啃木头的可能性与它们去取容易获得的水源的可能性一样大。还有许多其他同样令人困惑的行为变化例子(见Panksepp, 1998a, pp. 153-155)。研究人员思考了这些显著的发现,并得出结论:MFB-LH只是一个非常可塑的学习系统。
在同一时期,潘克塞普通过使用脑刺激来激发捕食行为,发现了类似的模式(Panksepp, 1971)。瓦伦斯坦认为这些发现支持他自己的结论,但潘克塞普提出了一个相当不同的理论观点:他认为所有这些都是MFB-LH中统一情绪系统的证据,该系统介导通用的食欲渴望和觅食行为。该系统是一个没有固定目标的驱动力,用于SEEKING所有奖励,并且随着学习逐渐形成对所有奖励的期待。用更行为化的术语来说,它是一个通用的、激励动机性的、食欲行为系统。如果这种系统反复被唤醒,那么动物最终会选择任何手边的食欲反应并坚持下去。这是一个情绪系统,而不仅仅是奖励系统。
多年后,Valenstein开始思考LH的唤醒是否标志着一种泛化的、非特异性的愉悦,这种愉悦使许多种类的消费变得令人愉快。Roy Wise(1982)也提出过这个观点,他是该领域的另一位先驱,最初认为证据支持存在许多沿着MFB-LH运行的消费子系统。为了测试这个假设,Valenstein请他在密歇根大学的年轻教师研究合作者Kent Berridge做一个关键实验。Berridge已经完成了他的博士研究,研究了一个迷人的现象:通过仔细观察大鼠的面部表情,特别是舌头运动,可以测量大鼠对糖水愉悦味道的水平。当糖水直接注入大鼠嘴里时,随着Berridge增加糖的浓度,动物会越来越用力地舔嘴唇,舌头伸得越来越远,几乎像卡通片中的滑稽角色。简而言之,愉悦感(“甜味”——一种感官情感)越强,大鼠舔嘴唇就越激烈。
Valenstein和Berridge推断,与其增加糖水的浓度(对人类来说相当于令人愉悦的甜味),不如通过对LH施加一点额外的电刺激来简单地增加愉悦感,LH是一种普遍的愉悦基质。换句话说,这种刺激应该增强Berridge擅长监测的消费性”喜欢”反应。在这种脑刺激的小”脉冲”期间,大鼠应该过度舔嘴唇,就像少量中等甜度的糖直接注入它们嘴里一样。这个实验做得很好(Berridge & Valenstein, 1991)。遗憾的是,对于Valenstein的理论来说,Berridge发现了完全相反的结果。当施加LH刺激时,舔嘴唇的行为并没有增加,反而急剧减少。显然,LH刺激并没有增加大鼠的消费性愉悦反应。因此,这种反应一定是由于某种其他类型的奖励。
Panksepp及其同事对这一结果感到高兴,因为他们已经认识到LH刺激会唤起SEEKING(寻求)冲动,这反映了强烈的觅食行为,这种行为通常发生在动物找到令人愉悦的东西之前。事实上,Berridge本人也得出了基本相同的结论,并继续培养他自己版本的SEEKING-EXPECTANCY(寻求-期待)假说。他认为该系统介导的是”想要”而不是”喜欢”(Berridge, 1996)。有相当长一段时间,他很难说服同事接受这一观点。直到今天,最普遍的观点仍然是,这个自我刺激情绪系统可以恰当地被称为”大脑奖励系统”,最近这个误称被大多数人脑成像研究者采用,他们的知识根源可以追溯到认知心理学。他们一直发现,这个系统的主要终端区域之一,伏隔核(见图3.2),在响应人类渴望和享受的一切事物时会像圣诞树一样亮起——从动人的音乐到一个好笑话(Knutson & Cooper, 2005)。Panksepp曾问Brian Knutson为什么不把它称为SEEKING系统,他表示如果使用这么激进的名称,他的工作将很难发表。换句话说,食欲学习的研究者更愿意将他们的动物视为感觉信息的被动整合者,增强”激励显著性”(incentive salience),而不是拥有大脑系统以欣快地参与世界以满足其需求的主动有机体。这是看待有机体如何在进化熔炉中构建的根本不同的方式。
一路上还有其他理论,但不会详细讨论(最近的总结见Panksepp & Moskal, 2008)。但没有任何理论像SEEKING系统假说那样具有包容性、行为学性和情绪性。如果有人仍然认为这个系统只介导美食或极佳性爱所产生的良好感觉,那么他或她没有关注所有的证据。最引人注目的观察,也是该领域大多数研究者仍然没有关注的,是接受这种脑刺激的动物会疯狂地探索它们的环境,注意它们遇到的所有新刺激。事实上,通过以某种方式组织环境,受刺激的动物往往会成为囤积者,当刺激开启时拾起各种物品,当刺激关闭时又丢下它们。因此,如果在测试箱的一半放置我们认为是垃圾的物品堆(软木塞、瓶盖等),然后安排当大鼠进入那一侧时SEEKING刺激开启,动物会把所有这些东西搬到箱子的另一侧,当脑刺激关闭时把它们放下。只是另一种强迫性的、附属性的行为!
总之,LH刺激不会产生独特的内稳态身体需求的感觉——它不会产生饥饿或口渴。相反,它促进了一种有利于自塑造(autoshaping)和大量附属行为的情绪”能量”。食物和饮料成为附属冲动的目标,导致疯狂地消费或与手头任何足够有趣的东西互动。因此,LH刺激也可以导致热衷于消费食物和水的事实,并不表明产生了任何特定的身体需求感觉(即内稳态情感)。相反,它是能够响应许多需求的基质,包括探索世界和追逐环境中有趣选项的需求。这项工作的大部分是在大脑的多巴胺网络被发现之前完成的。
20世纪70年代初,当所有这些实验进行时,Urban Ungerstedt(1971)发现了一个上行的多巴胺系统,该系统起源于腹侧被盖区(VTA),通过内侧前脑束-外侧下丘脑(MFB-LH)传递信息,上行至伏隔核,一直延伸到额叶皮层的内侧区域(图3.1A)。换句话说,很明显多巴胺通路是庞大而复杂的MFB-LH环路的重要组成部分,从中脑延伸到新皮层。这一环路被称为中脑边缘多巴胺通路和中脑皮层多巴胺通路,以及许多相关的神经通路,我们现在对它们的奖励性质有了相当多的了解(Ikemoto, 2007, 2010)。我们只是对它们总体上为有机体做了什么,或者如何谈论大脑的这种全局性情感功能(使动物和人类自发成为”主动”的有机体)还没有达成一致。
在20世纪70年代,Panksepp提出了这样一个观点:这些通路构成了一个寻求-期待(SEEKING-EXPECTANCY)系统。他的理论与之前所有将外侧下丘脑视为某种稳态或广义快乐-奖励基础的理论不同,他将其概念化为一个情绪脑系统,该系统产生期待行为和欣快-热情的情感,激励动物获取自然的恩赐并逃离危险。在这一观点中,附加行为和自动塑形是寻求过度刺激的自然结果。对MFB-LH自我刺激奖励所特有的狂热活动的这种替代性解释认识到,传统行为主义的”奖励”概念将这一系统的功能隐藏在一个模糊的通用标签之下。如果人们接受了”奖励”的概念,显然就不再需要思考该领域中的所有悖论了。
同样,Berridge得出结论,这个系统并不产生一种完成性满足感(“喜欢”),而是一种奖励性的食欲”想要”(Berridge & Valenstein 1991; Robinson & Berridge, 1993)。他和他的同事们继续论证,这个上行多巴胺系统增加了一种叫做”激励显著性(incentive salience)“的东西,这是一个略显模糊的概念,本质上意味着环境中的刺激吸引注意力的程度。事实上,这是许多情绪系统的一个特征—它们都帮助将感觉和认知信息输入大脑(见图2.1)。因此,这个想法类似于寻求-期待概念的一个关键方面—该观点认为哺乳动物有一种内在的冲动去伸手抓取吸引人的刺激,并逃离那些具有威胁性的刺激。然而,”想要”这个术语倾向于关注动物如何感知世界,而寻求假说则包括动物如何被设计为世界中的行动者—一个主动的代理者,而不是被动的信息处理器。
Panksepp和Berridge的假说之间还存在另一个关键差异。Berridge将”想要”和”喜欢”这两个词加上引号,以表明它们只是隐喻。他没有承认外侧下丘脑-多巴胺(寻求-“想要”)系统的活动会产生任何真实的内在情感体验。相反,他关注的是奖励的感觉特性以及预测奖励的刺激的效力/强度(“显著性”)。因此,Berridge曾一度提出,“喜欢”本质上是一个非体验性的过程,可能会在人类大脑的更高新皮层区域影响心理体验(即,这是另一个”读出”假说)。从这个角度看,很难想象为什么动物会进行自我刺激,也许只是因为这个系统的某些方面帮助在配备良好的人类大脑的更高部分创造了感觉,当然,这并不能解释为什么没有新皮层的动物会发现大脑刺激具有”奖励性”—它们的自我刺激同样很好(Huston & Borbély, 1973)。
Berridge选择将大鼠的”喜欢”设想为人类情感体验的无意识前因;这样他似乎绕过了心理过程或体验如何在其他动物中存在的问题。如果他的解释是正确的,Berridge是最复杂的读出理论家之一(见第2章)。他相信外侧下丘脑”奖励”系统的唤醒,特别是多巴胺能部分,是从人类新皮层产生的”想要”意识体验的前身。如果我们关注的是对未来遥远的特定对象和审美体验的预期,即三级心理过程,而不是欲望本身,这可能是合理的。
另一方面,Panksepp 比 Berridge 更早提出,热切渴望的原始情感体验——一种增强的纯粹欣快期待感——直接来自所有哺乳动物大脑中都存在的这些皮层下结构。换句话说,他提出其他动物是完全具有情感的生物,能够以热情的方式体验它们的寻求冲动。动物自我刺激外侧下丘脑并不是因为它感觉愉快(就像美食令人愉悦那样),而是因为它促进了一种内在的寻求状态,这种状态不仅产生对资源的搜寻,同时还产生一种非常特殊的积极感受,这种感受非常类似于我们人类对世界所包含的美好事物充满积极兴奋时的感受。但该系统最初并不知道它想要什么,这使得”想要”这个概念过于认知化,而情感性不足。无论如何,实际上每个研究者都隐含地同意这个系统介导大脑中某种类型的积极感受,但目前对于这种感受是什么样的还很少有共识或讨论。一个关键问题是,作为一个群体,行为神经科学家还没有准备好同意动物具有情感体验。事实上,大多数人还不愿意公开讨论动物情感感受的本质。我们认为,这不必要地贬低了其他动物,从而损害了我们自己的智识诚信。
当今大多数神经科学家并不太关心动物可能具有的情感感受。很少有人承认,研究其他动物的相关情感大脑机制是理解我们自己基本情感感受的唯一明确科学途径。大多数研究者更感兴趣的是这个”奖励”系统——我们更愿意称之为寻求系统——如何帮助动物大脑学习。因为神经科学家研究大脑,他们不只是像行为主义者那样关注环境中外部刺激的奖励和强化效应。相反,他们将注意力集中在可能介导奖励和强化的大脑区域、回路和神经化学上。他们最终在寻求神经科学答案来解决那些本应困扰行为主义者的问题,如果他们对学习机制是什么感兴趣的话。但大多数神经科学家没有认识到,情感体验的机制,即无条件刺激以及情绪无条件反应所唤起的感受的神经机制,都是允许大脑学习的”强化”过程的组成部分。
目前,大多数行为神经科学家同意该系统的主要化学物质多巴胺是条件学习的基本基质。在神经科学革命之前,行为心理学家提出了奖励/强化模型来解释条件反射是如何发生的。在经典实验中,像食物或饮料这样的奖励会在动物执行操作性行为(如按压杠杆)之后立即提供。行为心理学家提出,食物是强化学习的奖励。他们不知道强化的过程到底是什么,尽管他们都知道强化的程序(即一个反应之后跟随一个外部奖励)确实非常有效。过程问题显然需要大脑研究,而自我刺激”奖励”似乎是20世纪70年代初理解这一问题最明显的入口。
然而,在发现”大脑奖励”之前,奖励/强化学习理论中一直存在一个主要问题:没有人能够从大脑活动的角度有意义地解释奖励或强化实际上是什么,除了那些激发学习的东西。行为主义者将奖励定义为动物愿意为之工作的食物或饮料。但为什么动物会为食物和饮料而工作?因为它们是”有奖励性的”!强化也以同样循环的方式定义。像食物这样的刺激强化学习。但它是如何做到的?简单地说动物会学习行为模式以获得奖励是不够的。这类论证除了显而易见的事实之外什么也没有告诉我们。当”大脑奖励”被发现时,神经机制的研究终于对许多行为主义者产生了吸引力,但他们试图保留他们的旧术语。为了处理麻烦的情绪概念,他们提出情绪是通过学习产生的实体,即通过强化偶联性产生(例如,Gray, 1990),这引发了一些争论:Panksepp 提倡的另一种观点是,强化是情感感受和其他情感在大脑中促进学习的方式(Panksepp, 1990a)。
当新出现的神经行为学家开始以新的方式思考学习——从大脑回路和神经化学的角度——时,他们继续坚持他们传统的奖励和强化行为理论及其所有固有的模糊性。许多人认为他们在外侧下丘脑-多巴胺系统中找到了基本的学习基质,因为当动物被条件化时,多巴胺神经元总是以有趣的模式活跃。
行为主义流派中最现代的理论源于一系列多巴胺学习理论,即由瑞士电生理学家、现任职于英国剑bridge大学的沃尔夫拉姆·舒尔茨(Wolfram Schultz)提出的”奖励预测误差”假说。舒尔茨以精湛的技艺探测了饥饿猴子大脑中多巴胺神经元的放电模式,这些猴子期待着信号传递的食物。例如,如果闪光信号表示将送来猴子喜爱的食物,舒尔茨可以监测猴子首次接触到光线时、接触到食物时、光线预测食物时,以及光线亮起但猴子什么也没收到时(无疑会感到沮丧)的多巴胺活动。
舒尔茨观察到,猴子大脑中的多巴胺神经元最初对未预测到的食物传递做出反应,但随着猴子逐渐习得将提示(如闪光)与食物传递联系起来,多巴胺细胞逐渐停止对食物传递的放电,转而开始对光线放电——换句话说,对预测即将到来的食物的提示放电。然而,如果光线亮起而食物没有到来,多巴胺细胞的放电会出现轻微减少,这被认为发出了”奖励预测误差”信号,有助于完善学习。
请记住,行为主义者认为像食物和饮料这样的奖励刺激会强化学习。舒尔茨认为,从神经科学的角度来看,奖励最初表现为多巴胺神经元的快速放电。这种快速放电强化了学习。因此多巴胺放电的速率会教导猴子光线是食物的信号。当食物被省略时,多巴胺放电的减少是无奖励的(是一种惩罚),这进一步完善了学习。如果实验者在光线闪烁时不再提供任何食物,多巴胺神经元放电会更慢,这就是猴子学会光线不再是食物一致信号的方式。通过这种方式,舒尔茨得出结论,我们称为SEEKING系统中的多巴胺细胞构成了一个”教学信号”。但重要的是要注意,这些勤奋的猴子被束缚在它们的”课桌”上,因此无法展示出如果它们自由时肯定会表现出的许多有趣行为。这是环境如何控制行为的行为主义观点,而非动物的内在冲动。
舒尔茨假设多巴胺强化学习,因为多巴胺活动的变化总是伴随着学习过程。然而,科学的一个重要教训是”相关性不等同于因果关系”。通过将多巴胺神经元放电与特定的习得行为联系起来,人们看到的是相关物,而不一定是原因。当舒尔茨观察到多巴胺神经元放电随着动物条件反射而系统性变化时,他假设这些神经元在引领学习过程中扮演关键角色。同样可能的是,它们只是在跟随大脑其他地方发生的学习。我们相信这是对他精细数据的更正确解释。事实上,某些其他证据表明与似乎引导舒尔茨理论化的假设不一致(我们在上面关于自动塑形、附加行为和埃利奥特·瓦伦斯坦(Elliot Valenstein)小组的杰出工作的讨论中提到了一些)。
行为主义者的主要关注点是”学习如何在大脑中发生?“在这个重要问题上已经取得了很大进展。例如,最近关于FEAR系统和恐惧条件反射的研究(LeDoux, 2000),如第5章和第6章所总结的,揭示了学习在很大程度上依赖于谷氨酸(glutamate)递质。谷氨酸提供了一个门户,允许关于中性(条件)刺激的信息进入FEAR系统——这是它以前没有的通道。假设一只老鼠反复接触铃声,片刻后它的爪子会受到痛苦的电击。电击的疼痛无条件地唤起了FEAR系统。然而,在条件反射之前,铃声的响起没有唤起明显的情绪,既不焦虑也不担心,只有一个注意性的定向反应。条件反射之后,每当铃声响起时,老鼠明显变得害怕——大小便失禁的概率增加,自主神经指标(心率和血压)飙升。
这些实验表明,在条件反射之前,传递铃声信息的神经通路无法接入恐惧行为。条件反射之后,这条通路确实能够接入各种”恐惧”反应。通路是由一个特定的分子学习机制提供的,它是通向行为和自主神经输出的条件门户,简称为”恐惧”。这就是为什么条件反射的老鼠在铃声响起时表现出恐惧反应。这个分子机制是条件学习的神经关键(LeDoux, 2000)。在这些方案中完全没有为FEAR感受的神经机制提供位置。这是因为我们理应对动物拥有体验(即心智)的可能性保持永久怀疑。但如果大脑情绪活动的体验方面在许多学习过程中至关重要呢?
在伏隔核中存在类似的谷氨酸介导的学习机制,多巴胺系统向该区域发送其最重要的”奖赏”信息——即SEEKING(寻求)冲动(Kelley, 1999, 2004)。总体而言,Schultz收集的大多数证据与一个简单直接的可能性相一致,即当饥饿的动物获得可预测的食物时,条件性线索能够进入SEEKING回路。“奖赏预测误差”是一种复杂的说法,实际上是在表达另一件事——即动物能够区分那些持续预测奖赏的线索和那些不能预测的线索。据我们所知,这种区分发生在大脑的高级区域,而不是在Schultz记录多巴胺神经元放电的低级区域。但这些高级区域中的许多会让多巴胺神经元知晓其他地方正在发生的事情(见图3.1D)。
如果多巴胺活动不”强化”条件性/情绪性学习,那么为什么它们如此密切相关?简短的答案,正如刚才提到的,是它们接收关于大脑其他部分正在发生的学习的信息。这种学习正在启动SEEKING系统或在没有奖赏即将到来时抑制兴奋。这并不意味着多巴胺活动的变化直接参与了学习过程本身的介导,但这是一个合理的假设,需要在多巴胺轴突的终端场直接评估,特别是在伏隔核。然而,多巴胺活动也追随大脑高级部分中心理上令人满意的高阶状态的出现,如聆听动人的音乐、赌博和其他日常”成瘾”。这些是认知介导的预期状态,可能最初是由多巴胺的模式化释放构建的。
此外,当动物在食欲学习情境中首次获得食物时,多巴胺驱动的SEEKING冲动的初始唤起确保动物注意到条件刺激。SEEKING系统在食欲性条件反射期间总是被唤起,因为条件反射要求动物一开始就处于情绪唤起状态(也就是说,没有这种非条件反应,学习就不会发生)。在典型的实验条件下,情绪唤起是由非条件刺激引发的(即疼痛唤起FEAR(恐惧);奖励唤起SEEKING),而且这些非条件反应很可能能够打开大脑其他地方的学习通道。如果是这样,这将突出情绪唤起对动物模型中研究的许多类型学习的关键作用(例如,见第6章)。
这种通用的SEEKING反应不仅帮助动物自发地寻找并且凭借运气和技能找到它们需要的资源,而且还能帮助它们找到逃离危险的方法,它们最终需要学会避免危险。所有这些都需要环顾四周和探索环境。因此,如果你处于一种恼怒的愤怒状态,你的SEEKING系统也会被唤起。在不那么文明的社会中,你可能会以高度消极的方式表现出来。如果你的经理要求你承担更多任务,尽管你的工作量很重,你很可能希望大喊大叫并让他听听”你的想法”,但无论如何你保持沉默。你希望能设计一种优雅的方式来口头分享困扰你的事情——但当然,这需要自律,而自律通常是由过去的情绪教训培养起来的。无论如何,在所有这些情况下,通常在动物身上研究的简单学习自动来自激活RAGE(愤怒)和SEEKING系统的大脑机制的复杂性。这些情绪机制可能与”奖赏预测误差”假设设想的底层大脑系统相当不同。我们认为每个初级情绪过程的唤起对于实际创建一个将相关刺激”吸引”到其网络中的大规模神经动力学至关重要(见第6章)。换句话说,在情绪学习中,对非条件刺激的非条件反应在建立学习过程方面与非条件刺激本身同样重要,而许多研究者选择性地偏爱非条件刺激。请记住,大脑内的原始情感性情绪体验来自非条件情绪反应系统的唤起,与相关的环境事件共同作用。尽管这种食欲性学习机制尚未得到详细阐明,但正在取得很大进展(Alcaro et al., 2007; Kelley, 2004)。
我们怀疑食物”奖赏”的真正情绪学习机制与已经破译的厌恶性”惩罚”机制相似。例如,近年来,关于FEAR条件反射的神经科学研究(更多细节见第6章)发现,使条件刺激(即在足部电击之前响铃)进入FEAR系统的分子机制是关键的学习机制,这需要谷氨酸传递的变化,以便恐惧预测信号能够进入”恐惧输出”,如大多数人所说,或者如我们所声称的”FEAR回路”。在前一种观点中,人们对”输出”机制几乎没有什么兴趣。相反,在情感神经科学观点中,由于被条件化的是FEAR系统本身,人们密切关注FEAR系统本身的直接研究,以理解原始情绪学习(Panksepp, 1998a; Panksepp et al., 2011)。换句话说,条件反射机制可能与FEAR系统本身的非条件唤起密切相关。
如果我们能将这些知识应用到Schultz所研究的那类食欲性学习中,那么用情感性的情绪系统术语而非认知性的(“奖赏预测误差”)术语来概念化这种低层次的许可性”教学”过程会更明智。我们认为,SEEKING系统的视角为理解大脑低层区域(介导欣快性自我刺激奖赏)的组织方式提供了更连贯的整体图景。许多研究者仍在寻找一个独立于情感-情绪功能、值得被标记为强化的大脑过程,但在大脑的强大自动学习过程中尚未明确发现这一过程。也许看待简单经典条件反射的更好方式是设想非条件刺激和大脑的非条件反应倾向(两者都具有深刻的情感性)如何将外部信息吸引到它们的轨道中,使那些先前中性的刺激能够以越来越有模式和结构化的方式触发适应性情绪反应。
当我们真正理解原始情感体验的神经机制时,我们预期将对动物学习实际发生时我们所讨论的内容有更好的整体理解。行为主义者只用刺激和反应来表述。他们没有准备考虑看不见的神经心理过程。然而,大脑中确实存在各种初级过程的情感过程,除非我们恰当地将它们概念化,否则我们将无法理解生物体学习时真正发生了什么。根据这种更符合常识的观点,如果我们能从学习情境中的动物大脑里抹去情感,那么无论我们如何奖励或惩罚它们都无关紧要,因为它们不会学习。这听起来似乎过于基本,但当然,理解情感的真实神经本质却绝非如此简单。一个关键事实是,所有原始情绪系统都支配着那些学习发生的基底神经节脑区。
对大多数类型的动物学习的更准确理解应该需要理解大脑的情感机制。有可能行为神经科学家在试图理解非情感性的”强化”机制时,一直在追寻一个”斯纳克”——一种不存在的生物。要理解学习,我们需要更好地理解在大脑中拥有”奖赏”和”惩罚”意味着什么,我们需要确定这些神经机制如何促进学习。这种策略几乎是传统处理这些问题方式的镜像。我们不应只是”使用”奖赏和惩罚来促进学习,而需要理解使物体和事件成为奖赏和惩罚的大脑机制。这直接将我们带向大脑的情感本质,我们推测这最终将包含理解学习机制的关键要素。
当然,大脑中并非只有单一的奖赏或惩罚过程;它们有许多不同类型。但总体原则可能是相同的:更原始的情感脑机制明确控制着更高级脑功能的运作,从学习到思维([图2.3])。不幸的是,由于这个科学领域的历史,这些极具吸引力的替代可能性几乎未被讨论。
SEEKING唤醒和学习在多个方面紧密交织。SEEKING唤醒促使动物前往可能学习的新地方。SEEKING唤醒还诱导它们注意外部刺激,这通常是条件学习的必要条件之一。但这个方面可能完全无意识地发生。SEEKING系统最终还会产生重复的行为模式,伴随着现在由条件学习引导和构建的热情。然而,与SEEKING唤醒和学习的这种密切关系并不表明多巴胺活动是一个情感中性的”教学信号”;它是允许学习发生的情感丰富的神经状态。因此,我们预测,学习的产生将通过情感回路神经生物学的某些尚未理解的方面,可能通过波动的谷氨酸能传递,其中沉默突触(在年轻大脑中尤其丰富)在某些脑区被重构(即变成活跃突触)(参见[第6章])。再次强调,与其寻找强化信号,更有成效的观点可能是初级过程情感回路将相关的信息事件”拉入”它们自己的”轨道”,产生越来越结构化和有效的情绪行动系统。但这也可能导致各种附加行为、躁狂症状,以及妄想的自动塑造,这是偏执型精神分裂症的核心症状。而这个系统的持续低活性肯定会导致抑郁。
似乎像Schultz这样的神经科学家仍然将大脑设想为一个根据某种潜在强化原则学习的器官,该原则与产生多巴胺活动波动的刺激有关。然后,基于它所学到的,大脑指示生物体参与或脱离环境。这是一种被动的大脑观,将其视为首先学习、其次才产生行为的器官。SEEKING系统是一个自发的、无条件的行为生成器,它主动且探究性地将动物带到各个地方,在那里相关的学习机制允许它们发展知识结构,引导它们最重要的进化行动工具(内置的情绪系统)创造更多结构——更多更高级的心理过程——促进生存。
因此,作为另一种观点,我们认为多巴胺驱动的探寻系统不仅仅是一个学习系统,而是一个本质上促使人类和动物主动接触和积极参与世界的系统,从而促进学习。有时这种参与有助于准确学习;有时则不然。所有人都会同意Schultz的观点,即学习是大脑的主要功能之一——这一功能反映了许多其他相互作用的功能。然而,我们认为大脑是一个更具内在主动性的器官,在条件反射形成之前,它就促使生物体以好奇的方式参与世界。最终我们都会基于所学知识来参与世界,但最初的参与倾向,如婴儿所表现的,是一种非条件性的情绪情感反应,从根本上独立于个体学习。它是一种”祖先记忆”,使学习得以发生。
探寻系统反映了如此重要的祖先学习,以至于它被构建到我们的大脑组织中。换句话说,我们的初级过程祖先情感工具是编码在基因中的记忆,在我们的大脑中构建了生存的基本工具。因此,情感神经科学的观点是,所有哺乳动物生来就有以各种方式参与世界的冲动,这是探寻系统为神经科学带来的最根本贡献。如果没有这个系统,世界上任何有个人价值的事情都无法向前推进。父母和教育系统需要更有效地利用大脑的这种力量。
根据Schultz所遵循的经典行为主义观点,哺乳动物的大脑主要是一个用于学习的器官,其自发行为以及内在的情感和其他心理倾向似乎是次要的。在我们看来,哺乳动物的大脑以硬连线的方式促使我们以各种不同的(特定情绪的)方式主动与世界互动。这些祖先记忆(基本情绪)通过经验得到完善,但并非由经验创造。因此,尽管学习很重要,但我们不认为它是年轻人和动物最初参与环境的主要原因。相反,学习是一个自动的、无意识的过程,随着我们心智的成熟,它增强和完善了我们以越来越微妙的方式参与世界的自然倾向。另一方面,情感从来都不是无意识的。起初它是无知觉的(anoetic)——没有知识;但它迅速变成有知觉的(noetic)——被环境可供性的印记所充满,这构成了知识的开端。
探寻唤醒是大自然赋予的一种预期性礼物,它提供了看似无限的学习机会;随着高级心理过程的发展/表观遗传学出现,它逐渐微调合理的期望、工作假设,就像在科学研究中一样。这不是一个微妙的区别。但只需要稍微转换一下视角,就可以将Schultz关于多巴胺神经元波动放电模式的精细神经生理学数据视为对初级过程探寻系统的直接支持。
然而,探寻系统能够自发学习的一种非常特殊的方式。这不是我们一直在讨论的那种传统的条件性学习,而且似乎也不涉及思考。相反,它反映了这个系统能够衡量时间流逝的方式。这个系统可以学会自发地预测各种事件,特别是高度可预测的奖励事件。当我们讨论行为实验中常用的经典”强化程序”时,我们提到了一个对当前讨论特别有意义的程序。这些是固定间隔实验,在这些实验中,动物被允许通过按压杠杆、将鼻子戳入孔中(孔中有光电管自动记录这些探查行为)或执行各种其他任务来在固定的时间间隔获得奖励。在固定间隔程序中,动物在刚刚获得奖励后按压各种”操作按钮”——杠杆按压、鼻子戳等——的速度相当缓慢,但它们逐渐加速,直到在间隔的后半部分,它们以越来越高的频率按压杠杆。当这些操作行为模式在图表上绘制时,它们形成扇贝形状——一条明显向上的预期曲线。而这是自发发生的。
动物在为自我刺激奖励工作时也表现出这种扇贝式反应。但当奖励免费给予时,这种模式也会在动物的大脑和身体中自发出现。假设一只大鼠在规律的固定间隔(比如每20秒)免费获得LH刺激,因此它无需做任何事情就能获得每次奖励。在这个实验中,大鼠没有被给予杠杆或任何其他用于执行操作行为的装置。所有奖励都是免费的。动物可以选择”冷静如黄瓜”、像哲学家一样坐下来放松。然而,一种显著的预期模式出现了。固定间隔的大脑”奖励”产生了相同扇贝模式的自发嗅探行为(Clarke & Trowill, 1971; Panksepp, 1981a)。实际上,唤醒的嗅探是大鼠探寻唤醒的主要非条件性标志之一(Ikemoto & Panksepp, 1994; Rossi & Panksepp, 1992)。因此,似乎在高度周期性的探寻唤醒过程中发生了某种内在学习,逐渐产生了嗅探反应的扇贝模式。
作为同一过程的另一个自发出现的指标,最近我们还发现大鼠也表现出扇形模式的50千赫超声波发声(Burgdorf等,2000)——这是幼鼠在玩耍时发出的兴奋啁啾声(见第10章)。这些声音,就像活跃的探索性嗅闻一样,被认为是多巴胺激活的SEEKING系统的非条件反应(Burgdorf等,2001)。换句话说,在经历了一段时间这种固定间隔的免费奖励安排后,动物在大脑刺激后立即几乎没有嗅闻和啁啾声;但随着固定间隔的进行,嗅闻和啁啾的频率都系统性地上升,速度不断加快,直到接收到下一次大脑刺激为止(见图3.2)。然后这些指标又下降到非常低的水平。换句话说,该系统自动形成一条预期曲线,而没有任何东西被明确”强化”。显然,大脑是一个被设计为自发预测未来的器官,也许是因为这个系统介导了本章末尾描述的”心理时间”。
由于被唤起的SEEKING系统既产生升高的嗅闻又产生啁啾声,而这自然地形成一种预期模式,那么似乎SEEKING系统在某种程度上对奖励性情感的时机具有内在反应性。随着奖励传递时刻的临近,它变得越来越兴奋。SEEKING系统如何能够衡量这段时间的流逝?没有人确切知道,但众所周知,许多神经元具有自我生成的放电模式。虽然有些神经元只在受到某种外部影响刺激时才放电,但其他神经元具有一定水平的背景活动,这种活动来自某种”内部起搏器”——换句话说,一种计时机制。
SEEKING系统的含多巴胺神经元具有这种内源性起搏器,通常使它们以稳定单调的速率放电,就像钟表的滴答声,特别是当动物没有发生什么特别的事情时。这些神经元甚至在动物睡觉时也持续放电,但背景活动通常不伴随多巴胺的释放。SEEKING系统中多巴胺神经元的规律活动似乎几乎像钟表的秒针一样,以有条不紊的方式标记相当准确的心理时间。当系统以这种方式滴答作响时,它处于静止但具有信息性的状态。然而,当系统被唤起时,多巴胺神经元开始”爆发”并在快速连续放电几次时释放多巴胺。现在动物变得警觉并开始探索它的世界。或者如果动物在睡觉,它开始做梦,或者至少表现出REM模式,这是一种以高多巴胺活动为特征的状态(Dahan等,2007;Solms,2000)。
尽管研究尚未完成,但我们可以假设,在固定间隔实验期间,这种类型的神经元爆发和多巴胺释放增加正好发生在嗅闻和啁啾开始自发增加的时候。如果这个系统有一个内部计时机制,可以帮助动物预测何时表现出热切的期待——可以说是”第一个排队获取资源”——这对于理解生物体的基本行为和心理学将具有重大意义。大概正是SEEKING系统内活动的这种内部塑造,有助于解释当动物被要求在固定间隔时间表上为食物工作时所表现出的扇形行为模式。也许正是这一过程在我们的大脑中记录心理时间的流逝。
因此,当SEEKING系统被唤起时,多巴胺神经元的规律放电转变为更快速的爆发模式,可能也会导致动物的内部时间感加速。我们都听说过这样一句格言:快乐时光飞逝,这现在已经得到了实证证明(Droit-Volet & Meck,2007)。当我们愉快地从事某项活动时,特别是当我们有效地工作并朝着期望的目标努力时,时间似乎自由流动,没有磕磕碰碰的无聊感。也许这是因为在我们的SEEKING系统被唤起、我们的多巴胺神经元呈现爆发模式的这些时期,我们对主观时间的体验加速了——时间似乎过得更快,并伴随着一种令人愉悦的心理轻松感。
同样,多巴胺神经元对某些厌恶性事件的反应是抑制基线放电(Schultz,2006),但各种厌恶性事件也可以增加这种放电(Ungless,2004),这与各种负面情绪被唤起时SEEKING冲动的激发相一致。事实上,如果动物面对情感上的负面情境,多巴胺末端区域往往表现出可塑性,使它们更能够维持负面情感,因为这个系统可以介导”渴望和恐惧”,正如Kent Berridge及其同事所指出的(Faure等,2008,2010)。糟糕的时光会强化大脑中的负面情感回路。
当我们处于痛苦中或被忧虑困扰时——当我们度过艰难时光时——我们对时间本身的感觉往往会减慢。同样,众所周知,帕金森病患者的多巴胺神经元退化,他们对时间的感觉也发生了改变。没有促进多巴胺传递的药物,这些人会陷入清醒的”睡眠”——他们感觉自己被冻结在时间中,生活在一个看似没有事件的无聊、倦怠和心理空虚的宇宙中(Sacks,1973)。
除了这些关于多巴胺放电模式的重要观察之外,我们还不知道多巴胺神经元的放电如何计算时间感。此外,我们也不知道这种时间感如何调节SEEKING系统的唤醒,使其在固定间隔时间表的前半段保持相对静止,然后在后半段变得越来越活跃。尽管这些想法的许多方面仍有待正式测试,但来自大鼠的数据越来越多地表明,它们的时间感与人类一样,是由多巴胺控制的(Meck et al., 2008)。
我们开始理解为什么生物体在奖励传递的固定间隔计时期间会变得如此奇妙地具有预期性,这阐明了关于时间流逝感知与我们SEEKING系统产生的预期渴望之间关系的深刻奥秘。目前,我们可以确信,我们对时间流逝的感觉是一种基本的心理功能,它使我们能够预测环境中的变化事件。时间是否也是宇宙的基本属性则更具争议性(Barbour, 2000),但很明显,如果没有这种进化而来的心理过程,我们就无法连贯地讨论宇宙的本质或我们在其中的位置。
我们只是触及了这个迷人系统的一些特征。还有很多需要学习的。例如,有人提出,REM睡眠——做梦睡眠——可能通过SEEKING系统的过度唤醒产生其一系列充满情感和兴奋的幻觉事件(Panksepp, 1998a; Solms, 2000)。事实上,最近的研究表明,多巴胺细胞在REM睡眠期间比在安静清醒期间表现出更多的爆发,并分泌更多的多巴胺(Dahan et al., 2007; Léna et al., 2005)。看来我们梦境的情感景观是由与生活中食欲兴奋相同的化学物质所激发的。这表明做梦的一个功能是帮助预测和应对我们面临的情感挑战。尽管取得了巨大进展,但关于这个系统仍有许多未解之谜,包括它参与精神病妄想、幻觉、梦境和对未来预期的确切方式。
尽管存在这些局限性,我们对这个系统的深入理解使我们能够有效地思考人性的一些神秘方面——例如,被称为”感觉寻求(sensation seeking)“的心理特质(Daitzman & Zuckerman, 1980; Zuckerman & Kuhlman, 2000)。为什么人们喜欢从事危险的工作和运动?攀岩者报告了这样的体验。即使在危险中,他们也全神贯注于寻找下一个支撑点,找出最佳的身体定位方式,并在危险的垂直地形上前进。似乎SEEKING系统带来的快乐使他们保持活力,并分散他们对运动危险性的注意力。
想想消防员。许多人热爱这份工作,尽管他们自愿定期将自己暴露在危险之中。恐惧是一种非常负面的情感,人们会期望消防员害怕去上班。当然,消防提供了对社区有价值的服务,这可能是一种自豪感的来源,这显然是一种三级过程情绪,可以抵消每天面对恐惧的痛苦。消防员与同事分享宝贵的同志情谊。这也可以建立社会纽带,从而补偿长期恐惧的痛苦。我们将永远铭记在9/11事件中为帮助他人而失去生命的数百名英雄,以及那些每天冒着生命危险营救火灾和其他灾难受害者的人。
一些有创意的心理治疗师可能会说,消防员正在掌控童年的恐惧,而掌控的快感是他们可能热爱工作的原因之一。这种观点可能有其价值:对情绪的认知掌控可能帮助消防员克服对受伤或死亡的恐惧。但我们也可以尝试从SEEKING系统提供欢快情感的能力来理解消防员对工作的热爱,这种欢快情感可以抵消甚至有时完全消除恐惧的持续痛苦。此外,由于SEEKING系统唤醒新皮层,促使其制定策略和解决方案,消防员的欢快情感可能很容易与他们危险工作的细节联系在一起,这些工作穿插着长时间的单调和重复性例行公事。然而,当他们的SEEKING系统被唤醒时——当他们奋力扑灭危险火焰时不可避免地会发生这种情况——消防员将全神贯注于灭火和从燃烧的建筑物中救人的工作。这种强大而专注的投入提供了令人兴奋的冒险体验,增强了消防员作为世界上一股有效和重要力量的自我感觉。这些都是SEEKING系统提供的积极情感。但我们必须怀疑,他们的经历在多大程度上也强化了他们思想中某些负面情感回路。
人们理论上也可以想象许多其他与精神问题的联系。例如,我们想知道SEEKING唤醒是否会导致”自恋”问题。自恋是指人们对自己的感觉。在日常用语中,自恋通常带有贬义:它意味着某人过度关注自己。然而,只要一个人的自我评价是现实且积极的,自恋在情感上可以是健康的。病理性自恋通常发生在早期生活经历损害了一个人的价值感时。这些人试图通过以某种方式过度补偿和高估自己来让自己感觉更好。如果孩子从父母或老师那里得到了太多不切实际的表扬,他们也可能变得自恋。
SEEKING唤醒的表现之一是增强了自己作为世界中有效行动者的感觉。在社会世界中,这需要感到重要、有吸引力、成功和优越。多巴胺产生增强的自尊;这可能是理解自恋问题的一个神经化学关键,这也有助于解释为什么自恋问题如此难以治疗。如果自恋是由多巴胺驱动的,这是一种高度成瘾的脑化学物质,会助长重复强迫,那么自恋症状将特别令人满足且难以放弃。自恋也可能支撑一种虚假的自信和主导感,这种感觉可能像纸牌屋一样崩溃,导致一个人走上一种可能特别顽固的抑郁之路,因为它直接打击一个人的自尊核心。
同样有趣的是,一些自恋患者倾向于进行马拉松式的幻想,幻想自己是主角的辉煌壮举。这在青春期尤其明显,并且可能是正常的。这些患者经常说,当他们如此投入时,他们不知道时间去了哪里。过度唤醒的SEEKING系统可能解释了时光飞逝的感觉。如果过度自恋和时间飞逝都是SEEKING唤醒的迹象,那么这些轶事报告就有了新的意义。
如果这些想法是正确的,那么在某些情况下,轻度剂量的抗精神病药物可能会促进对自恋患者的心理治疗。这些药物可能会抑制多巴胺带来的令人振奋的愉悦感,使患者更愿意为他或她的问题寻找现实生活中的解决方案。当然,有效药物的剂量需要谨慎衡量,因为多巴胺活性过低也会促进抑郁。
我们可以写一整本关于SEEKING-期待系统的书。在这最长的一章中,我们试图提供一些描述这个非凡大脑系统的广泛轮廓。我们相信它的功能多年来一直被误解。许多行为科学家仍然误解它,他们用被动的”信息处理”术语来理解学习。他们专注于研究在受控实验的监狱式禁锢中的动物,而不是在真实世界中”信息寻求”的主动框架中,在那里所有哺乳动物、鸟类、爬行动物和复杂无脊椎动物在过自然生活时必须主动照顾它们的身体需求。在1950年代被发现后的几十年里,奖励-SEEKING系统被视为一种完成性/稳态奖励基质。近年来,它被视为一种学习强化系统。
我们认为它两者都不是。这是一个促使我们主动——积极主动地——与世界互动的系统,以便找到我们茁壮成长所需的资源,以及避免危险和威胁。它自动促进食欲性学习,通常以错觉的方式(例如,自动塑形)。它激发我们所有的能力,从最基本的冲动到抽象思维的最高境界。因此,SEEKING系统对所有动物的健康和福祉至关重要,包括人类。然而,这个系统的故障可能导致病理状况,从系统长期不活跃时的极度抑郁到过度活跃时的妄想性躁狂和偏执性精神分裂症。在特殊条件下,它甚至可能促进负面情绪。
这个系统在本质上所有其他积极感觉的食欲阶段都起着至关重要的作用,以及逃离特定惩罚和从更持久的其他糟糕时期中解脱。这是成瘾药物导致强迫行为模式的原因之一。例如,在戒断成瘾药物期间可能经历的烦躁不安,这种类似抑郁的状态,可以通过再次服用缺失的药物来迅速缓解(行为主义者称这种现象为”负强化”——惩罚的缓解——而不是使用直接的情感概念”解脱”)。换句话说,药物成瘾如此难以治疗的原因之一是戒断效应如此强烈且消极,以至于人们学会了自我治疗。
当我们期待任何事情时,当我们为任何事情努力时,当我们竭力逃避任何事情时,SEEKING系统会激发我们的行为和态度。除了成为渴求行为的核心之外,它还创造了许多学习形式所必需的条件,包括操作性条件反射(operant conditioning),因为它促使我们探索新的物理和智力领域,并且因为它将平凡的活动转变为令人兴奋的追求,即使在情绪动荡之中也是如此。该系统还促进行为模式,最终被整合到习得性预期条件反应中。动物的新训练程序,如”响片训练”(clicker training),利用了动物想要做似乎在自己控制之下的事情的自然倾向(Pryor, 2005)。特别令人着迷的是,多巴胺系统衡量心理时间的流逝,这是急切期待的必要成分。
SEEKING系统,虽然通常仍被称为”奖励系统”,但已成为新兴神经经济学(neuroeconomics)领域未被承认的宠儿(Knutson & Cooper, 2005),通过分析SEEKING系统和岛叶(insula)的厌恶反应——这个隐藏在额叶和颞叶之间及其下方的组织岛——可以预测人们何时选择购买物品或不购买它们(Grosenick et al., 2008)。如果岛叶的厌恶系统被激活,这个人就不会购买;相反,当伏隔核(nucleus accumbens)的SEEKING冲动被激活时,这个人就会掏钱包。当我们聆听情感动人的音乐时,中脑边缘多巴胺系统的同一终端区域也会被唤醒(Blood & Zatorre, 2001)。该系统激发我们的梦想(Solms, 2002)以及许多其他心理上的愉悦,有时也包括恐怖。正如我们将在下一章看到的,该系统在掠夺性行为中也非常重要,比如性跟踪。当然,我们对互联网的沉迷反映了SEEKING系统的作用。未来的研究可能会揭示这个非凡的通用系统的许多其他特定能力,它被设计用来SEEKING任何东西和一切。
任何人都可以生气,这很容易;但是对正确的人生气,生气到正确的程度,在正确的时间,为了正确的目的,以正确的方式,这不是每个人都能做到的,这并不容易。
——亚里士多德(公元前320年)
人类似乎对爱有无尽的渴望。如果有人”夺走”了我们这种情感宝藏,我们就会发现自己同样无限的悲伤和孤独能力——以及原始的愤怒(RAGE),它可以转化为嫉妒和仇恨。在这种激情的控制下,我们体验到一种强烈的欲望,想要伸手去打某人——不是随便什么人,而是我们认为应该为释放我们的愤怒负责的那个人。我们体验到涌入思想的愤怒(outRAGE)是一种祖先留下的宝藏,它帮助保护我们,现在仍然如此。但我们的RAGE初级加工能力不需要一个有意的仇恨对象;它是一种纯粹的感觉。当然,我们的愤怒(一种次级加工情绪)总是有某个被认为是RAGE原因的对象。凭借我们丰富的思考大脑空间,我们在心智的更高层次孵化仇恨——充满各种复仇计划。有时我们会制定惩罚敌人的现实计划。但更多时候我们在幻想中这样做,不会产生持久的满足感,往往还会毒害我们的心灵。
主要对我们心智的三级加工层次感兴趣的心理学家,在列举我们愤怒的许多细微差别时没有困难,甚至到了它似乎完全溶解成一种认知态度的地步。所以它通常被这样定义。Jim Averill(2010)的定义指出,愤怒是指一种情绪状态,既包括对某种感知到的错误的责任归因,也包括纠正这种错误或防止其再次发生的冲动;攻击是试图强迫他人违背其意愿采取或不采取某种行动,而不是为了他或她自己的利益([p. 4],原文强调)。
然后Averill提到了很少有人问过的十个问题。它们非常值得一读,从”狗会生气吗?“到”当你发泄愤怒时,你在发泄什么?“简而言之,对于第一个问题,他建议”如果我试图拿走我的狗的骨头,它可能会对我咆哮和咬;但它并不生气,因为它不懂愤怒的语言和概念。然而,我的狗正在体验某种东西;它不是一个自动机器,它的攻击性行为让人想起愤怒。如果不是愤怒,那么,我们可以叫它什么?“([p. 8])。他接着明智地使用这里建议的分析层次,并将狗的愤怒置于”次级加工”习得性激怒类别中,而我们人类显然具有”三级加工”愤怒(如上所定义)。对于另一个问题,他建议”什么都没有,我认为。然而,确实有些东西在改变……在宣泄期间,不需要失去任何东西,而且可能会获得很多东西,即对事物真实面貌的新见解,也许不是绝对的,但是潜在的。如果这是对宣泄的准确解释,它也意味着一种新的情绪观点,在这种观点中,情绪对创造性变化的可能性是开放的”([p. 20])。这本质上陈述了心理治疗的主要目标——清楚地看到你的情绪并学会使用它们来改善我们的生活。
但我们如何了解狗的”愤怒”的认知方面呢?其他动物是否会计划和幻想击败和杀死对手?其他动物是否像人类那样体验仇恨?我们不知道。但如果大脑发达的黑猩猩和大象的心智中不存在怨恨,那将是令人惊讶的。尽管我们可能永远无法真正知道它们在想什么,但有大量轶事证据表明它们有”报复”的倾向。从科学角度来说,理解它们的初级过程情感比理解它们的高级心理活动要容易得多。基于确凿的数据,我们可以确信其他动物拥有能够产生高度激怒行为和值得被称为愤怒(RAGE)的负面情感感受的脑系统。正如艾弗里尔所认识到的,用通俗术语”愤怒(anger)“来称呼它会让我们陷入无法解决的困惑。但我们可以预测,动物愤怒(RAGE)的机制确实会激发人类愤怒的感受,而现在我们处于主流科学范畴:我们的想法可以被证伪。它们也可以得到支持。例如,脑内阿片类物质会抑制愤怒(RAGE)回路,我们可以预期显而易见的结果:阿片类物质应该在减少人类愤怒感受方面非常有效,从而也应该削弱仇恨和复仇欲望的力量。
然而,在本章中,我们主要关注的不是仇恨或愤怒的想法或复仇计划。仇恨和复仇是三级过程,反映了我们思考所经历的不公并设计详细报复方案的能力。也许大多数其他哺乳动物不具备进行这种反复思考的认知能力。尽管如此,它们确实表达愤怒(RAGE),这从根本上不是为了惩罚,而是为了迅速让其他个体符合自己的隐含(进化)欲望。据我们所知,所有哺乳动物都会对那些竞争资源的其他个体体验愤怒(RAGE)。因为愤怒(anger)和仇恨(hatred)是愤怒(RAGE)在我们认知中展开的方式,所以在我们的日常语言中很难将这些相互作用的概念区分开来。这突出了所有初级过程情感的一个重要观点——我们有许多情感术语是建立在我们基本情感的神经能量之上和之外的认知阐释。本论述的总体前提是,初级过程情感唤起始终参与我们高级情感过程的各种体验,但我们都可以同意,没有人设计出好的科学方法来探究那些心理微妙之处,它们反映了我们的认知如何被我们的激情所改变。
人类的愤怒(anger)总是在困难时期增加,当有许多挫折时——在经济衰退时期,或者当某些看似必需的资源,从汽油到工作到爱的感受都稀缺时。在稀缺时期,脾气爆发的频率必然比在富足时期更高。在认知层面,令人恼火的分歧可能是日常生活的一部分。复仇的感觉很容易爆发,特别是在那些互相欺凌但从未成为朋友的年轻人之中。所有这些人类问题在Pahlavan(2010)中都有很好的讨论。但我们伸手去打某人的冲动,无论是言语上还是身体上的脑源在哪里?动物脑研究能告诉我们很多关于这些问题的信息吗?我们的答案是有条件的肯定。许多与人类愤怒(anger)和仇恨(hatred)相关的高级认知过程在神经科学上仍然难以渗透,特别是当人类使用语言的方式,从沮丧和和解的语调到指责和认知性和解,可以放大或减弱愤怒(anger)的激情时。但愤怒(RAGE)的原始状态可以通过困难的动物脑研究得到详细理解(Panksepp, 1998a; Siegel, 2005)。
因此,动物脑研究不会让我们理解文化化的人类价值观的更微妙方面——那些可以对抗我们动物本能的存在方式。例如,跨物种情感神经科学不能告诉我们太多关于能够平息被激起的愤怒(RAGE)的安抚姿态和宽恕的质量。宽恕的能力,就像感到悔恨的能力一样,是基于大多数动物可能不具备的复杂认知过程。然而,动物研究可以阐明在大脑内部愤怒(RAGE)爆发意味着什么。当然,这种初级过程感受可以导致人类之间许多应受谴责和伤害性的行为;这些行为可能以矛盾的方式证明是自我挫败的。负面情感,在人类心智的高级认知领域内,似乎有一种适得其反的方式。
攻击性(aggression)也有许多面孔。在人类中,有自我中心、自恋的反社会者和精神病患者,他们只是掠夺性的,不在乎伤害谁。更糟糕的是,有些人积极想要伤害他人并享受这样做。我们也将在本章中探讨这种掠夺性冲动,尽管我们的大部分内容将致力于讨论愤怒(anger)能力的祖先根源。我们对这些根源的了解来自对哺乳动物大脑初级过程愤怒(RAGE)系统细节的理解。为了理解人类愤怒(anger)的根源,我们必须在相关的动物模型中非常详细地研究这一强大的情感系统。
不幸的是,近年来关于这一系统的大脑研究几乎从神经科学领域消失了。为什么RAGE研究被抛弃了?在某种程度上,答案是出于政治动机。20世纪90年代初,美国国家精神卫生研究所(National Institute of Mental Health)的一位首席行政官在组织一场关于暴力生物学根源的会议时,提出了一个不敏感且政治不正确的建议。他建议——也许没有深思熟虑这些问题——内城贫民区类似于丛林,因此动物研究可以帮助我们理解这些贫民区的文化问题。一种解读是,这隐含地将生活在内城的男性比作超攻击性的灵长类动物。这一暗示被视为既冒犯又带有种族主义色彩。会议被取消了。这场喧嚣至今仍给攻击性的神经生物学研究蒙上了阴影。
关于攻击性的研究也减少了,因为对动物愤怒的研究常常导致一只实验室动物攻击另一只,这种做法可以理解地令许多人反感。因此,斗鸡和斗狗等行为在许多州和国家已被适当地禁止,在科学研究中也常常必须采取预防措施,以防一只动物严重伤害另一只。
但肆无忌惮的愤怒并不局限于任何人类亚群,实际上也不局限于任何哺乳动物物种。我们现在掌握的知识足以自信地断言,RAGE系统存在于所有哺乳动物的大脑中。我们知道这些回路位于何处,也了解一些能够唤起或抑制攻击性易怒的化学物质。但仍有许多有待了解的内容,包括这些感受究竟如何在人类心智的高级大脑空间中发挥作用。如果实验设计具有一定程度的谨慎和敏感性,神经科学家没有理由忽视哺乳动物大脑(包括我们的)中内置的RAGE潜能。我们对这些回路的神经生物学了解得越多,就越能理解这一对生存至关重要的天然工具,它可能在家庭生活和整个社会中造成大混乱。也许我们还可以为控制这种激情的药物产生新想法——帮助消解已成为精神病学重大问题的愤怒感受。当人们变得令人反感地愤怒时,一个常见的建议是”吃片药”!这样的药物实际上并不存在,但有一些有希望的药物开发线索正被忽视,因为这不是被接受的精神病学适应症。
各种情况会无条件地唤起RAGE:身体活动受限或身体表面受到刺激都容易引发这种感受。在次级层面,当SEEKING系统的愿望受挫时,人和动物也会感到愤怒,比如预期奖励的突然撤回。在前一章中,我们提到了一个微不足道但常见的例子:当自动售货机未能提供承诺的零食时,期待如何迅速转变为愤怒。这种失望相对温和,愤怒很快就会消散。然而,如果你对梦想中的房子提出了报价,却发现它被出价更高的竞标者抢走了,也许是被你特别讨厌的人抢走了,你的挫败感会更强烈,你可能会在一段时间内保持情绪低落和怨恨。虽然这通常被称为生气,但我们认为证据表明,这种激发的感受是由RAGE回路产生的,我们将在本章中讨论。最令人担忧的是童年虐待或忽视,这可能引发持续一生的愤怒。RAGE在战争和社会动荡时期可能会剧烈爆发。但夫妻之间为琐事无休止地争吵也太过常见,年幼的孩子可能会在自己家中目睹攻击行为和相关的不公正。
体内平衡失调,比如食物匮乏引起的饥饿,也会使RAGE冲动变得敏感。在第三章中,我们注意到过度的SEEKING唤起会导致附加行为(adjunctive behaviors)的出现,这些是无用的仪式性活动。附加行为似乎也在一定程度上是由挫折引发的RAGE唤起的。正如我们在上一章讨论的,当动物非常饥饿且无法轻易满足饥饿时,实验室中会出现附加行为。相反,动物被小块食物”逗弄”,使它们保持在持续的SEEKING状态。换句话说,当人和动物过度饥饿、口渴或性挫折,并且无法轻易获得满足时,愤怒很可能会产生。尽管SEEKING系统仍处于唤起状态,尽管SEEKING唤起能产生积极的热情情感,但RAGE系统也可能因挫折而同时被唤起,两种激情可以协同作用。虽然RAGE本身不是认知性的(即它不是由信息处理创建的心理状态),但它注定会通过学习与认知影响交织在一起。
例如,像失去爱这样微妙的情况在神经科学上不容易研究,但当我们的社会欲望受挫时,愤怒情绪很容易产生。兄弟姐妹之间的竞争可能是最常见的例子之一。如果年长的孩子担心新生儿会抢走父母的爱,他们可能会开始憎恨新的兄弟姐妹。有时年长的孩子会问什么时候把婴儿送回医院,或者建议把婴儿冲进马桶可能是个好主意!这些烦躁的认知反应,为愤怒和仇恨提供了肥沃的土壤,并不仅限于幼儿。在大多数与性伴侣生活在一起的成年雄性哺乳动物中,最容易激起攻击性愤怒的方法之一就是在它们的领地中引入另一只雄性。人类成年人的嫉妒在历史上引发了暴力行为,有时甚至导致谋杀。考虑到即使是最幸福生活中不可避免的变故,很容易理解为什么某些愤怒激发的时刻是每个人生命中不可避免的特征。无论脾气多好,没有人能免于这种情感体验。这是我们祖先遗产的一部分。然而,亚里士多德式的情感智慧(phronesis)最终可以使愤怒成为一种平衡的工具——让我们能够选择对谁生气、以什么强度生气以及生气多久。从长远来看,理解和和解可能是最值得追求的选择。
Alan Siegel(2005)详细描述了攻击行为的神经解剖学。愤怒系统从杏仁核的内侧区域向下延伸,主要通过终纹的弯曲通路到达内侧下丘脑,然后到达导水管周围灰质(PAG)的特定区域(见图4.1)。在迄今为止测试过的所有动物中,通过电刺激这些大脑区域都可以引发愤怒。当电流开启时,动物会迅速攻击,通常会咬它们面前的物体。当电流水平增加时,攻击会变得更加激烈。如果在人类身上进行这类脑刺激程序,人们往往会咬紧牙关并报告强烈的愤怒感(King, 1961; Mark等, 1972; Hitchcock & Cairns, 1973)。但受试者不明白他们为什么变得愤怒——他们很难为自己的感受提供任何理性的原因,因为眼前没有现实的攻击对象。人们觉得这种体验令人不安,因为在正常情况下,人类的愤怒有一个引发的对象或事件,并自动被新皮层的三级过程中的愤怒和仇恨所阐释,始终伴随着具体的怨恨和关于该责备谁的想法。但那些外部诱发事件和想法并不总是存在。如前所述,愤怒反应也可能因某些身体变化而加剧,例如饥饿。血压升高也往往会使愤怒系统变得敏感(Mancia & Zanchetti, 1981)。同样,脑部病变,如侵犯相关回路的肿瘤,会刺激愤怒系统,使人类和动物都越来越可能表现出自发的、看似无目的的攻击行为(Blumer, 2000)。
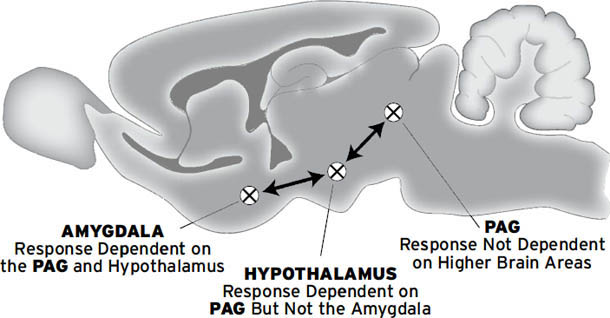
图4.1. 大脑中愤怒的层级控制。圆圈表示可以通过局部脑刺激引发愤怒的主要脑区。X表示损伤,因此对较高区域(如杏仁核)的损伤不会减少从较低区域(下丘脑和导水管周围灰质[PAG])引发的反应,而对较低区域的损伤会损害较高区域的功能。下丘脑损伤会消除来自杏仁核的反应,但不会影响PAG,而PAG的损伤会显著降低从较高脑区引发的愤怒反应(摘自Panksepp, 1998a;经牛津大学出版社许可重新发布)。
愤怒回路是分层排列的,较深的结构对于实际产生攻击行为比位于大脑较高位置的结构更为关键。从PAG引发的愤怒不会因内侧下丘脑或杏仁核的损伤而减少(DeMolina & Hunsperger, 1962)。然而,PAG或内侧下丘脑的损伤可以完全消除从杏仁核引发的愤怒。正如人们所预期的那样,对系统中部——下丘脑的损伤会阻止来自杏仁核的愤怒,但不会阻止来自PAG的愤怒。因此,可以说PAG对这种情绪至关重要,内侧下丘脑很重要但重要性较低,而内侧杏仁核对这种本能情绪反应的产生甚至更不重要。但杏仁核对于建立引发愤怒的认知联系更为相关。大部分认知联系通过内侧杏仁核从较高的脑区传递过来,这些脑区阐释恶意的反刍思维。但对于完整的情绪反应,PAG和内侧下丘脑区域仍然至关重要。
这种层级排列突出了所有基本情绪系统的一个普遍原则。每个情绪系统中较低、更古老的部分对于它们产生的连贯情绪反应更为关键,包括原始的愤怒感受,比高级脑区更重要。这种层级控制在所有情绪网络中都很明显。不幸的是,这些控制层级并未对所有情绪系统进行充分研究,尽管对寻求系统和恐惧系统都有很好的佐证研究。例如,人们早就知道,低级脑区的损伤对自我刺激的影响比高级脑区的损伤更大(Huston & Borbély, 1974; Valenstein, 1966)。
可以推测,基本的生理”刺激”如饥饿和激素/性欲挫折,通过大脑的其他部分进入愤怒系统,比如监测身体稳态(homeostasis)的内侧下丘脑。例如,饥饿的动物总是比营养充足的动物更容易打斗。饥饿如何与愤怒联系起来,迄今为止还没有得到太多神经科学的关注。这很遗憾,尤其是在花费如此多资源去寻找”正在寻找功能”的大脑机制的时候。
这并不意味着高级控制在日常愤怒中不重要。它们当然重要,尤其是对于习得性愤怒反应,从认知引发的愤怒到对某人的持续仇恨。来自最高级脑区的大量认知信息可以输入愤怒系统,为中脑导水管周围灰质中精心设计的粗略情绪编排提供微妙的改进。例如,皮层感知到的各种环境刺激,部分通过新皮层/认知输入传递到内侧杏仁核区域,该区域位于愤怒系统的最顶端。人类,以及可能一些动物,可以使用这些高级控制来掌控愤怒的感受。再次,正如亚里士多德在本章开头的题词中强调的,对愤怒获得理性的控制是智慧的一个方面。有时,心理治疗在走向成熟的过程中提供了巨大帮助,在那里人们成为情绪的主人而不是奴隶。
几位早期研究者认为,对这个系统的电刺激并没有产生任何真正的愤怒类型的感受,而只是产生了假性愤怒(rage的行为表现而没有相应的主观情感)。这似乎是合理的,因为一小部分动物即使在嘶嘶作响和咆哮时也可以被抚摸(Masserman, 1941)。然而,在该子集中受到刺激的电极位置似乎位于脑干的相当低的位置,即运动表现的实际最终共同通路发散到脊髓的地方,或者在三叉神经(也称为颅神经V)运动核的附近,该神经控制咬合的力度。这可能解释了为什么没有触发完整的情感性愤怒反应。这种情感上空洞的大脑位置很少在愤怒回路的高级区域中发现。
现在看来,中脑愤怒网络执行部分(导水管周围灰质内)及其上方的大多数电极放置确实会唤起一种与原始人类愤怒非常相似的中枢情感状态,除了人类通常是因为某种感知到的过错而对某人生气这一事实。除了这些认知成分外,另一个区别是电诱导的愤怒在电刺激结束后不会持续很长时间,可能是因为没有思想来维持这些感受,或者可能是因为突然释放了一个对立过程(opponent process)[1],比如停止刺激时的迅速缓解,以及寻求回路中平衡活动的恢复。
值得注意的是,沃尔特·赫斯在20世纪30年代中期首次在猫脑中发现愤怒系统(他因此在1949年获得诺贝尔奖),使用下丘脑的局部刺激,他是最早提出该行为是”假性愤怒”的人之一。然而,他在退休后发表的著作中承认(如第2章所述:例如《心灵的生物学》[1964]),他一直相信动物实际上经历了真正的愤怒。他承认分享了他自己并不相信的观点。为什么?他只是不想让自己的工作被当时占主导地位的行为主义者边缘化,这些人无法容忍谈论情绪体验。结果,我们仍然不太了解愤怒系统如何与大脑的其他认知和情感系统相互作用。
同时,在这种情况下,我们应该强调,下丘脑的RAGE系统部分(集中在腹外侧和邻近的基底下丘脑)与SEEKING回路(集中在背外侧区域)以及FEAR回路(集中在更偏内侧腹部的区域)非常接近。因此,某些电极位置很可能同时刺激RAGE和其他系统之一。如果是这种情况,那么来自SEEKING冲动的积极情感将抵消RAGE反应产生的消极情感。这可能解释了为什么动物有时会自我刺激那些能引发类似RAGE攻击性的位置。相比之下,同时刺激RAGE和附近的FEAR系统(将在下一章详细介绍)可能会产生比单独RAGE更强的防御性甚至更强的厌恶感。事实上,这种混合情况会变得非常混乱,特别是因为我们现在有充分的理由相信,掠食性攻击(我们将在下文讨论的话题)是由SEEKING系统的唤醒所促进的。
在使用局部脑电刺激时,记住这些问题很重要,因为许多系统在空间上紧密相邻,并且经常在行为序列的控制中相互作用。很少有情况是单独刺激一个系统。也许这些关于刺激相邻网络的困难可以通过新兴的神经科学技术来解决。例如,将产生视紫红质的分子病毒植入特定的大脑区域(这可以使神经元对光敏感)可以精确定位,从而能够用特定波长的光从许多重叠的网络中选择性地只激活一个大脑神经化学网络。因此,我们现在能够通过植入大脑正确区域的光纤更有选择性地只刺激一个大脑系统(Airan et al., 2009)。同样,用特定神经化学物质进行局部脑刺激也可以提供对特定系统的选择性刺激,其精确度是电刺激无法达到的(Ikemoto, 2010)。这些进展应该会大幅完善我们对基本情感系统功能细节的认识。
神经科学家对影响RAGE的脑化学物质了解很多(Guerra et al., 2010; Siegel, 2005)。能够促进RAGE的化学物质,通常在存在其他支持性刺激的情况下,包括睾酮、P物质、去甲肾上腺素(NE)、谷氨酸、乙酰胆碱和一氧化氮合酶。这些影响中的许多可以用药物抑制。例如,因为脑内去甲肾上腺素可以促进愤怒,普萘洛尔(一种阻断β-NE受体的药物)可以减少易怒性,但这也适用于其他类型的唤醒。其他减少RAGE的化学物质包括血清素,特别是埃托普嗪(一种血清素激动剂,有时被称为宁静剂药物,可增强血清素的作用)所强调的,但同样这种效应并非特异性的。血清素倾向于降低所有形式的情绪唤醒。RAGE抑制剂的清单还有很多。也许最突出的一种是γ-氨基丁酸(GABA),这是大脑的通用抑制性神经递质。GABA能降低RAGE活动,但它也会降低大脑其他广泛活动的神经放电率。换句话说,GABA也倾向于抑制每种情绪、抑制癫痫发作,并且在促进睡眠方面非常有效。因此,就像血清素一样,它对RAGE系统并不具有特异性。
我们列出这些化学物质只是为了强调,每个大脑系统都受到多种化学物质的控制。但正如我们将看到的,似乎有一些化学物质,如神经肽P物质,在大脑的某些高级区域确实更特异性地激活RAGE(尽管在较低的区域它们促进完全不同的大脑功能,如恶心)。其他神经肽如内源性阿片类物质(大脑的吗啡模拟物)以及催产素(另一种大脑社交安慰和增强信心的化学物质)也可以非常有效地平息RAGE。但同样,它们在大脑中也发挥许多其他作用。所有这些都表明,某些情绪的神经化学控制在单个大脑回路水平上可能相当精确,但它们在其他大脑系统中也可能有不同的效应。这是为精神病学实践设计更精确的”精神药物”一直很困难的原因之一。
部分由于每只动物都表现出特征性的神经化学优势和劣势,情绪气质必然在个体之间以及物种之间存在广泛差异。已经开发出基于情绪的人格量表来识别人类气质(Davis et al., 2003),但为动物设计这些量表会更加困难。然而,我们通常可以通过选择性育种(即通过应用”行为遗传学”技术)相当容易地培育出动物的情绪特质差异。
我们也知道,雄性和雌性在几乎所有情绪系统中都有不同的敏感性。大量的动物研究表明,总体而言,雌性在生物学上比雄性更不容易愤怒。循环性激素的差异,即使在人类中,也至少是造成这种性别差异的部分原因。睾酮明显使雄性比雌性更自信和更具攻击性。事实上,当人类女性被注入睾酮时,她们很快就会变得更具攻击性且对他人的容忍度降低(Hermans et al., 2008)。因为睾酮也促进雄性的支配倾向,它似乎积极影响几种不同形式的攻击性。
当然,睾酮/攻击性的联系仅适用于身体攻击。还有其他发怒的方式和造成伤害的方式,其中最恶劣的可能是社交排斥(social rejection)(MacDonald & Jensen-Campbell, 2011)。当人或动物被剥夺爱和接纳,当他们被拒绝并被迫进入社会等级的较低阶层,在那里他们几乎没有权利和更少的快乐,这通常会造成情感伤害。虽然社交排斥不会造成即时的身体伤害,但谁能说心理伤害从长远来看不是同样有害的呢?毕竟,压力可能是致命的,而社交排斥会引发巨大的压力。似乎一个物种的雌性,当然包括人类物种的女性,完全有能力对他人造成这些情感和社会伤害。因此,尽管身体伤害通常似乎是男性的领域,但女性往往更擅长对心理而非身体造成更微妙的伤害,其对健康的不利影响相当(Knack et al., 2011)。如果有人怀疑女孩的攻击意图,他们只需深入研究任何教室中女孩之间的社交政治即可。一个关键问题是,这种倾向是否反映了潜在的初级加工情绪系统的差异,还是更容易受到学习和文化影响的高级认知过程的差异。关于这些问题的研究很少,但我们认为这更多地与心智的三级过程有关,而非初级过程,这意味着在许多过度攻击的案例中,社会和文化干预必然比生物干预更重要。
愤怒(RAGE),就像每一种基本情绪一样,受到许多心理过程和许多大脑区域的调节。我们将总结一些引人注目的,尽管有时令人困惑的发现,主要是为了强调每个情绪系统的整体调节是多么复杂,以及还有多少有待了解。
例如,对不包括在愤怒系统中的大脑部分进行某些受限的损伤可以显著提高攻击性。腹内侧下丘脑(ventromedial hypothalamic, VMH)损伤(使动物过度进食并变得严重肥胖,并抑制雌性性行为)也可以使动物长期易怒——简直是野蛮的——如果没有防护装备几乎无法处理。这种变化是持久的,即使随后反复温和处理也几乎不会减轻。我们不能确定为什么这些损伤会加剧愤怒,但也许这种脑损伤产生的疤痕组织正在长期刺激邻近的愤怒回路。在癫痫病灶靠近内侧杏仁核愤怒回路的人类中,可以看到类似的长期易怒现象(Mirsky & Siegel, 1994)。此外,来自下丘脑最内侧基底部弓状核(arcuate nucleus)附近的神经网络,它向大脑其他部分发送身体能量充足的信号,可能直接调节攻击回路,当身体的能量资源充足时抑制它们。
另一方面,动物的愤怒刺激可以通过刺激某些高级回路(如外侧隔区(lateral septum)中的回路)来改善。这导致研究人员推测外侧隔区可以调节和抑制愤怒系统(Brayley & Albert, 1977)。这可能解释了”隔区愤怒(septal rage)“这一戏剧性现象,即对这一中线脑区的损伤可以显著提高攻击性长达数周。这些损伤的动物对触摸和许多其他刺激过度敏感。通过对杏仁核中的愤怒和可能的恐惧回路进行额外的损伤,这种易怒可以迅速减少(Jonason et al., 1973)。同样,随着时间的推移和大量的温和处理,这些隔区损伤的动物逐渐变得非常温顺和不具攻击性,最终甚至比正常情况下更具亲社会性。简单的时间流逝以及非威胁性的生活经历可以缓和隔区损伤后的过度活跃的愤怒反应。在腹内侧下丘脑损伤的动物中没有观察到类似的恢复。隔区位于许多重要的情绪和认知系统的十字路口,这表明它对于高级认知系统和低级情绪系统之间的相互作用尤为重要。事实上,它是大脑中另一个主要的情绪/认知交叉点,就像我们已经在寻求系统中看到的伏隔核(nucleus accumbens)一样,以及当我们讨论恐惧系统时将会看到的杏仁核外侧区域。因此,当隔区受损时,皮层抑制被削弱,导致至少在一段时间内出现更多的情绪表现。目前还不完全清楚为什么隔区损伤的动物最终变得比以前更加平静和具有社交性。但显然它们对社交奖励变得更敏感。
正如我们之前提到的,新皮层的移除,尤其是额叶执行区域,会增加情绪性。最早发现的这类现象之一是”去皮层狂暴”——如果外科手术移除了额叶皮层区域,狗和猫会变得非常易怒。此外,愤怒也受连接到脑干的”小脑”——小脑的控制,小脑曾经被认为只控制我们动作的平滑协调。小脑最深层和最古老的核团,即顶核(fastigial)和中间核(interpositus),在受到电刺激时可以产生攻击行为。有些人认为,也许就像新皮层抑制和调节情绪以产生更有分寸的行为和心理反应一样,小脑皮层——小脑的外缘——可能会调节攻击性行为倾向。事实上,情况可能确实如此。例如,罗伯特·希思(Robert Heath)是一位神经外科医生,他在精神外科时代(尤其是在20世纪50年代)对人类大脑的情绪做了大量研究,他认为可以通过刺激暴力患者的小脑皮层区域来抑制攻击性。据报道,这一程序确实非常有效(Heath et al., 1980)。但它从未被采纳为常规做法,这可能主要是由于对人脑进行这种直接技术操作的伦理担忧。
直到今天,我们仍然没有高效的方法来控制病理性暴力,除了可能使用产生极度镇静作用的药物。尽管对攻击性进行了大量研究,精神病学尚未开发出能够充分抑制持续性愤怒/怒气的可行药物,无论是对人还是对动物。因此,社会仍然容易受到生活在精神和情绪易怒的腐蚀性控制下的危险个体的威胁。由于P物质(Substance P)已被证明会加剧猫的愤怒(Gregg & Siegel, 2003; Siegel, 2005),我们长期以来一直主张P物质受体拮抗剂,如阿瑞匹坦(aprepitant)(现已获医学批准用于治疗恶心),可能作为抗愤怒、抗易怒药物非常有效。然而,这一提议仍有待在人类中进行评估。尽管如此,越来越多的研究显示这类药物如何持续地减少动物的愤怒型攻击(Halasz et al., 2008)。该系统内的受体变异也与人类的攻击性和自杀倾向有关(Giegling et al., 2007)。在这种情况下,重要的是要注意大脑中的大多数神经化学受体系统都有几种变体。例如,就P物质而言,有NK1、NK2和NK3受体(NK代表神经激肽(neurokinin),即P物质所属的神经肽家族)。只有NK1受体会促进攻击性。可以说,在这方面,任何释放到神经系统中的神经化学物质只有在有相应的特定类型受体可用时才会产生特定效果。虽然这是一个需要记住的重要点,但我们会避免在本书中详细阐述,因为本书旨在让非专业读者也能理解。
精神病学家需要明白,仅仅通过抑制过度活跃的愤怒系统无法实现心理健康。愤怒通常通过对社会后果的理解和积极社会关系的激发来平息。如果有朝一日针对过度愤怒的药理学”疗法”问世,这类药物应该伴随心理治疗干预,增强患者享受与朋友和家人积极联系的能力。换句话说,在寻找药理学药物时,精神病学家不应该简单地试图将愤怒作为一种不受欢迎的行为类型来根除。这是一条普遍规则:精神病学家应该意识到不同情绪原型之间的情感互动,并应寻找最大化幸福感的方法,其特征是促进快乐和社会和谐的丰富积极情感。显然,社会政策也是实现这些目标的有效工具。
我们对大脑所有内置情绪系统的认识远未完整。目前,人类研究者,即使是那些进行脑成像的研究者,还没有可视化愤怒系统和其他主要过程情绪系统工作方式的方法。部分原因是他们没有常规的实验方法来对这些古老的情绪系统进行因果研究。功能性磁共振成像(fMRI)在可视化高级认知大脑功能方面比在低级情感功能方面要好得多,因为前者的神经活动速率远大于控制我们情绪的古老大脑网络。这些局限性意味着,即使使用现代脑成像技术,我们也无法轻易可视化愤怒网络爆发时的强度。我们也无法轻易监测人脑中释放的促进愤怒的化学物质的数量。
此外,在人类脑成像技术的限制内很难激发强烈的愤怒。fMRI扫描追踪大脑中的血流,假设更多的血液会流向神经活动增加的区域。然而,在fMRI扫描中,受试者需要保持头部不动。因此,如果激发了强烈的情感,如果技术要正常工作,人类受试者需要主动抑制表达这些情感的冲动(即他们需要抑制本能行为)。因此,化学PET成像的大脑功能必然更有效地理解人类的低级愤怒回路。
尽管如此,一些优秀的研究已经对愤怒进行了成像。在最早的研究之一中,愤怒的感觉确实导致了各种中线皮层下区域的激活,但特别明显的是额叶皮层区域的激活,尤其是在大脑左侧。当人们感到焦虑时,这些大脑区域也会”点亮”。神经活动的抑制(血流减少)在介导认知的各种高级脑区更为明显,特别是在大脑右侧。当人们焦虑时,大脑的额叶皮层区域受到抑制。当人们愤怒时,大脑后部的区域,包括映射体表的顶叶区域,受到抑制(Kimbrell et al., 1999)。其他研究人员在愤怒和悲伤的面部表情期间,观察到前扣带回和后扣带回区域有相当程度的激活,杏仁核对悲伤有一些独特的反应,而眶额叶皮层对愤怒有独特反应——眶额叶皮层位于眼眶正上方,参与多种情感感受(Berlin et al., 2004; Blair et al., 1999)。这些激活中有多少可以被认为是情绪的初级过程表现,而不是感觉和内稳态情感?有多少与次级和三级调节过程有关?在这类研究中不可能知道这些答案;事实上,大多数大脑成像研究人员并不关心这些重要的区别。我们不期望这些大脑变化的列表对普通读者特别有启发性,因为对大脑成像研究人员自己来说,如何解释这些扫描结果也绝非清楚。
作为成像数据模糊性的一个很好的例子,在愤怒感受期间在成像研究中点亮的眶额叶皮层,可能倾向于抑制愤怒而不是激发愤怒,因为该区域的损伤通常会增加人类的易怒性和冲动性(Berlin et al., 2004)。此外,当大脑中稍靠后的附近内侧区域(如腹侧纹状体,它是SEEKING系统的一部分)受损时,人们很难识别他人的愤怒(Calder et al., 2004)。为什么会这样仍不清楚,但这可能再次表明SEEKING系统参与调节了多少种情绪。当然,如果一个人渴望攻击某人,SEEKING系统的某些部分必然会被激活。如果是这样,可以想象,如果该系统受损,人们可能难以感知愤怒引起的激活。但为什么在其他试图在人脑扫描中捕捉愤怒的研究中,SEEKING系统没有被激活呢?也许是因为功能性磁共振成像(fMRI)技术的局限性,其中神经元放电必须发生相当显著的变化才能被检测到。当然,在核磁共振扫描仪的限制条件下,实验环境根本不利于体验强烈的情绪。我们必须再次记住,介导情绪的大多数皮层下区域的神经元放电速度相当慢,相比之下,像丘脑和新皮层这样的高级脑区在介导感知和认知时放电速度极快。因此,微小的变化,特别是在皮层下情绪区域的变化,很难用功能性磁共振成像检测到。需要采用能够进行更长时间曝光的不同技术,如正电子发射断层扫描(PET)成像,以及更精细的实验方法,以充分成像情绪过程。
事实上,通过使用这类替代的大脑成像技术(如PET扫描),一些研究人员在非常低的脑区观察到了强烈的血流变化(表明神经激活)。在愤怒期间,Damasio等人(2000)发现在大脑内侧脑干深处的血流强烈增加,那里是PAG(导水管周围灰质)——情绪性的中心——所在的位置,以及一些邻近的脑区,如控制整体大脑唤醒的蓝斑核。这项杰出的PET研究强调,当愤怒和大多数其他初级过程情绪(恐惧、快乐和悲伤)被激发时,高级皮层区域往往会关闭。这表明强烈的情绪感受可以损害或缩小认知处理,这一现象长期以来一直被心智学者所认识。当情绪被强烈体验时,新皮层的许多区域会关闭,这一事实再次凸显了我们在大脑中最强烈地感受情绪的位置,即我们与其他动物共有的古老的皮层下情绪网络。
重要的是要强调,关于哺乳动物大脑中RAGE网络位置的大部分知识都是从动物研究中收集的,只有偶尔从人类那里获得相关数据。因此,现在就断言人类大脑在日常愤怒和怨恨期间究竟发生了什么还为时过早。然而,我们预期,如果人类大脑中没有RAGE系统,他们将很难产生充满仇恨的激怒。
人们倾向于认为人类的愤怒有助于激发战争动机,但这将是一种过度概括。即使在激烈的战斗中,战术上有效的士兵通常也不会处于暴怒状态,尽管在肉搏战中这种激情肯定会出现。显然,大量的社会学、政治和历史因素在战争中起着重要作用。而且,正如贪婪和统治等高级情感冲动所反映的,寻求系统(SEEKING system)在人类战争中可能比原始过程的愤怒回路(RAGE circuits)更具影响力。此外,如果我们的愤怒能力导致了所有战争,那么我们可能会看到其他物种进行更多的集体战斗;然而很少有其他动物表现出这种群体攻击性。黑猩猩等近缘物种偶尔会与其他群体发生集体冲突(Goodall, 1986),但与青少年或年长流氓团伙的类比可能比战争冲突更合适。无论如何,目前我们完全无法说清楚愤怲冲动(RAGE impulses)相对于掠夺性冲动在各种形式的群体攻击中贡献了多少。也许某些类型的愤怒只有在动物真正投入到激烈的攻击状态时才会爆发,此时原始情感可能会迅速燃烧和转换。
因此,我们在这里所说的很少能够阐明人类物种战争的原因。当然,人类的战争倾向最终伴随着许多仇恨情绪,包括贪婪、怨恨和得意,更不用说强奸和掠夺等行为,但据我们有限的知识,这些复杂的情感,就像我们的嫉妒、怨恨和仇恨一样,并不是哺乳动物大脑古老情感部分的本能原始过程潜能。它们可能通过发育和社会学习从高级脑区产生。其他动物不具备我们所拥有的新皮层复杂性。因此,大多数其他动物根本无法像我们这样对这些事情产生复杂的想法和感受。但这并不是说它们无法产生更简单的原始怨恨(proto-resentments)、原始嫉妒(proto-jealousies)和原始仇恨(proto-hatreds)。尽管如此,像恐惧(FEAR)和愤怒(RAGE)这样的基本情感肯定会在每个战场上爆发,这些情感源于我们与所有其他哺乳动物共享的情感系统。
在前面的章节中,我们强调总体而言新皮层会抑制那些唤起新皮层的情感系统。我们还指出,当被唤起时,新皮层,特别是支持工作记忆(思考策略能力)的背外侧区域,可以触发和维持情感(更深入的讨论见第6章)。愤怒(RAGE)特别清楚地展示了这些原则。很容易看出新皮层如何引发和维持愤怒。在他”不加掩饰”的自传中,著名的具有神经科学背景的精神分析学家约翰·盖多(John Gedo, 1997)描述了他如何回应导师告诉他,他在芝加哥精神分析研究所的新课程未获批准,因为课程委员会认为他”作为讲师还不够成熟,无法获得这样的特权”。基于观察和推理(所有这些都是认知/新皮层功能),盖多确信导师”亲自策划了这一结果”,因为他们曾共同教授的一门课程产生了过往的不满。这些想法引发了盖多的愤怒,他爆发了。正如他所说——“不再需要取悦他,我用足以让整个研究所都能听到的声音告诉他,他可以把他那该死的课程塞到屁股里去!我一生中很少如此愤怒”。即使是人类心智方面的专家偶尔也需要发泄他们的动物本能。但宣泄对你有好处吗?毫无疑问,这取决于它是否给你带来了你想要的东西,而从长远来看,你应该想要的最重要的东西是正念(mindfulness)和智慧。
显然,即使在极其聪明的人身上,相当轻微的认知触发也可以引发愤怒攻击,有时是出于相当琐碎的原因。也许盖多在这次爆发中消耗了他的愤怒;然而,人们可以想象他可能在一段时间内仍然怨恨不已,思考报复的机会,并可能制定真正的计划来破坏那位被憎恨的导师。这样,他的新皮层(他的思想)就会让他的愤怒保持活跃,并维持他愤怒系统的唤起。然而,如果情况不同,盖多的新皮层可能会促使他控制住自己的愤怒。有办法让自己远离你不想拥有的感受,恢复镇定的两个主要方法是深呼吸几次和反思你想成为什么样的人。新皮层总是关注关于什么可能增加奖励和生活满意度以及惩罚将如何降低幸福感的想法。盖多发泄了他的愤怒,因为他已经受到了惩罚,没有什么可失去的了。然而,假设他在芝加哥研究所的一些资深同事批准了他的课程并推翻了导师的反对。盖多可能仍然怨恨他的导师,但他可能会推理任何愤怒的表达都可能疏远其他是他盟友的同事。因此,如果他愿意的话,他可能会保持沉默,他的新皮层可以抑制他的愤怒系统。
这些新皮质计算也影响着原始RAGE系统在其他动物真实互动中的表现方式。例如,如果电刺激猴子等更复杂、认知更成熟的生物的RAGE系统,被激怒的动物通常倾向于将愤怒发泄到更顺从的动物身上,并避免与更具统治地位的动物对抗。然而,新皮质或许能够考虑到这样一个事实:从长远来看,愤怒的动物往往会提升社会地位。事实上,在猴群中,如果持续长时间重复刺激某只特定猴子的RAGE系统,该动物可能在现有的统治等级中地位上升(Delgado, 1969; Alexander & Perachio, 1973)。或许持续的易怒情绪可以帮助动物克服既定的统治关系。然而在自然界中,通常是雌性选择哪些炫耀实力的雄性被允许登上权力的巅峰。如果这样的雄性失去了大多数雌性的青睐,他很快就会被众多在一旁等待的热切追求者击败。
我们知道RAGE是一种不愉快的情感,不仅因为人们这样说,还因为动物和人类都会试图避免对这个系统的电刺激。当刺激无法避免时,动物会表现出逃避行为,表明它们希望终止这种情感体验。尽管如此,确实有些人似乎对RAGE有一种渴望,似乎享受愤怒的感觉。如果RAGE不可避免地导致人际交往中的成功(胜利),动物和人类可能有时会享受RAGE。可以很容易想象,拳击场上的拳击手可能遭受许多激起他愤怒的打击,然后他可能更加享受击倒对手的过程。换句话说,愤怒的表现可以带来许多次要好处。同样,如果人们知道自己身处电影院的安全环境中,或者在某种狂欢节装置上摇摆,身体以其他方式会引发强烈负面情感的方式被抛来抛去,他们可能会享受FEAR的体验。
如果愤怒被用作防御,它也能提供解脱。所有防御都能提供一些快乐,或者至少能减轻痛苦和困扰。例如,憎恨一个抛弃自己的恋人,比无助地忍受被拒绝的爱情之痛感觉更好。但在大多数情况下,纯粹的RAGE感觉很糟糕,这是心理治疗师和咨询师需要记住的重要考量。我们经常看到患有持续愤怒的人,乍一看似乎享受愤怒的感觉。但这可能是因为愤怒产生了一种强烈的态度,可能被误认为是热情(实际上,在叙述愤怒事件时可能会唤起SEEKING状态)。人们可能看起来享受愤怒,只是因为他们主动寻找麻烦,以非理性和不公正的方式挑起争论,可能获得只有他们自己才能理解的次要好处(例如权力感)。也许他们享受愤怒胜利的时刻,但没有人或动物享受持续RAGE的体验,因为这种情感感受根本不愉快。在绝大多数情况下,长期愤怒的人无法轻易控制自己的愤怒;有些人似乎情不自禁地寻找争斗,也许是因为在他们生命中的某个阶段,某些事情或某个人让他们感到无助的愤怒。还必须记住,在某些非常不寻常的医学情况下,例如当人们患有刺激RAGE回路的脑肿瘤时,人们可能会变得长期易怒,尽管他们没有合理的外部愤怒理由。
治疗师应该知道愤怒从根本上是一种不愉快的情绪。长期愤怒的人是困扰和不快乐的。他们可能一生都在愤怒,从未体验过真正解决分歧后的内心平静。他们在愤怒的争吵中反复循环,通常以不令人满意的情感僵局告终。如果我们知道愤怒感觉很悲惨,并且我们向患者传达这一认识,这本身就能提供解脱,因为愤怒的人通常甚至不考虑他们愤怒是有原因的这种可能性。通常他们只是认为自己天生愤怒,因此是坏人。
多年前,August Aichhorn(1925)写道,年轻的违法患者应该始终知道治疗师站在他或她这一边。经典的中立立场对这些年轻人不适用(如果它真的适用的话——但那是另一个讨论!)。神经科学可以提供建立这种治疗关系的关键,因为它告诉我们RAGE唤起感觉很糟糕。如果患者患有慢性愤怒,治疗师可以诚实地告诉他,虽然有时满足愤怒可能感觉良好,但总的来说这是一种悲惨的感觉方式,没有人选择感到愤怒。所以如果患者大部分时间都感到愤怒,那一定是有原因的。某些事情或某个人引发了这种愤怒。他并不是因为自己是坏人而简单地选择愤怒。这是神经科学见解可以帮助建立诚实治疗同盟的一种方式。它可以提供对情感生活本质的理解。这种理解可以作为共情但诚实地探索患者感受和心理状态的基础。
同时必须强调的是,攻击行为并不仅仅是 RAGE 系统在起作用。特别重要的是要关注这样一个事实:一种攻击形式,即所谓的掠食性攻击(predatory aggression),主要源于 SEEKING 系统,人们很容易混淆攻击和愤怒。事实上,神经科学家很难接受动物的”安静撕咬”掠食性攻击,就像我们人类的狩猎冲动一样,更多地来自 SEEKING 系统的心理能量,而非 RAGE 系统。从某种意义上说,SEEKING 系统总是在寻找令人满意的终点,无论是捕食者追逐一顿活动的美餐,还是人类渴望赢得一场激烈的争论。攻击有多种形式,将在下一节中讨论。
动物有两种主要的攻击行为类型,它们不是 RAGE 系统的纯粹表现。第一种是掠食性攻击,发生在动物捕猎食物时。食物以其他动物的形式出现,捕食者会杀死它们,我们通常认为杀戮是一种攻击行为。然而,目前的神经科学证据表明,掠食性攻击是 SEEKING 冲动的表现。当掠食性动物跟踪并杀死猎物时,它们似乎体验到的是期待的愉悦,而不是 RAGE 的尖锐刺痛。当然,如果猎物激烈反抗或碰巧逃脱,那么动物会合理地感到沮丧和烦躁,但这是因为 SEEKING 系统受到了挫折,没有获得体内平衡的满足,即一顿美餐。
现代社会很少提供人类掠食性攻击的例子,因为食物供应充足,至少在发达国家是这样。所以我们没有必要狩猎。在超市里觅食通常就足够了。然而,掠食性性攻击在许多现代社会中仍然猖獗。例如,某些形式的强奸在多大程度上是由 SEEKING 而非 RAGE 能量驱动的,这必然会引起争议,但在我们目前的知识状态下,这无法通过科学确定。
大多数食肉动物确实会捕猎食物,神经科学家对猫和老鼠进行了大量研究,这些研究证明了 RAGE 和掠食性攻击之间的决定性差异。几乎所有的猫都会进行安静撕咬的掠食性攻击,这是一种相对受控的,如果不是平静的,跟踪、杀戮和有条不紊地撕咬猎物的行为(Bandler, 1988; Flynn, 1976; Siegel, 2005)。跟踪和安静撕咬都可以通过电刺激外侧下丘脑区的内侧前脑束来产生,该区域位于 SEEKING 系统的核心。另一方面,猫的 RAGE 系统的唤起会产生截然不同的行为。愤怒的猫会咆哮和发出嘶嘶声。它们的毛发竖立,并表现出自主神经唤起(如心跳加快、血压升高、肌肉血流增加和体温升高)。这不是猫在跟踪和捕获猎物时的行为方式。这些关于猫的数据表明,掠食性攻击是由 SEEKING 冲动而非 RAGE 支配的。
在实验室大鼠身上可以看到进一步的证据,大多数实验室大鼠不像野生大鼠那样表现出强烈的掠食倾向。也许这些倾向已经从实验室种群中被培育出去了。然而,相当比例的实验室大鼠明显具有掠食性(它们很容易攻击较小的动物),而有些几乎具有掠食性(它们对小鼠等潜在猎物表现出极大的兴趣,但未能撕咬它们)。神经科学家发现,这些几乎完全掠食性的动物可以通过刺激它们的 SEEKING 系统而转变为安静撕咬的攻击模式,这再次表明掠食性攻击显然是被唤起的 SEEKING 系统的反映,而不是 RAGE。事实上,这些动物会自我刺激它们的 SEEKING 系统,直到它们对小鼠表现出完全的掠食性攻击。换句话说,动物放大了自己的 SEEKING 冲动,达到了激励它们成为掠食性杀鼠者的程度。如果没有额外的自我施加的 SEEKING 系统人工唤起,就不会观察到这种完全完成的行为模式(Panksepp, 1971)。
大鼠表现出另一种区分 SEEKING 和 RAGE 的行为差异。当它们表现出由 SEEKING 唤起产生的安静撕咬掠食性攻击时,它们会撕咬活的和死的小鼠。然而,当它们的 RAGE 系统被唤起时,大鼠只会攻击活的动物。它们会简单地从死小鼠身上走过(Panksepp, 1971)。显然,当动物愤怒时,它们需要一个活的目标来发泄愤怒。愤怒的动物也会攻击同种动物(同一物种的其他成员),但它们不会将其视为猎物(即,同种动物通常不是掠食活动的适当目标)。
这可能是家长和治疗师需要牢记的一个有趣观点。当孩子生气时,有时会被鼓励将愤怒发泄在无生命物体上,如枕头或拳击袋。然而,这可能是一种无效的疗法,因为愤怒(RAGE)似乎只针对有生命的目标;让孩子对无生命物体进行报复甚至可能增加其挫败感。也许如果孩子努力想象枕头是,比如说,一个讨厌的兄弟姐妹,这可能为攻击性提供真实的表达。然而,这种方法并不可取。我们已经说过,所有情绪系统如果被过度唤起都可能被敏化(sensitize)。如果使用这样的策略人为地唤起愤怒系统,结果可能不会起到宣泄(cathartic)作用。它更可能敏化一个已经岌岌可危的过度唤起系统。这些事实对暴力电视节目和电脑游戏也有影响。尽管如此,在治疗环境中真诚地表达愤怒可以帮助建立相关的治疗对话,而且短时间模拟表演愤怒冲动,如模拟掐枕头,在治疗指导下可以有效地用于促进被压抑情绪冲动的情感解决。
除了行为差异外,愤怒(RAGE)在各种解剖学和药理学方面也与捕食性攻击不同。通过刺激大脑的不同区域,人们可以选择性地调节捕食性攻击或情感性攻击(Siegel, 2005)。小剂量镇定剂减少愤怒(RAGE)并增加安静咬攻击的机会。另一方面,安非他明(精神兴奋剂)可以增加愤怒(RAGE),而对捕食性攻击几乎没有影响。如前所述,P物质促进愤怒(RAGE),中等剂量的阿片类药物抑制愤怒(RAGE),而低剂量的阿片类药物可以促进寻求(SEEKING)(其他几种神经肽也可以,例如神经降压素、催产素和食欲素)。还有其他非常一般的兴奋性和抑制性控制,如谷氨酸和GABA,促进和抑制这两个系统,以及所有其他原始过程情绪系统。神经化学相互作用对各种类型攻击的影响是如此复杂,以至于需要大量篇幅来描述已经发现的大量内容(Miczek, 1987; Siegel, 2005)。
最重要的是,动物渴望自我刺激(例如,按压杠杆),以实现对诱导安静咬攻击的寻求(SEEKING)脑部位点的电刺激。这表明动物喜欢由促进捕食的寻求(SEEKING)唤起产生的情感感受。但如果刺激诱导纯粹愤怒(RAGE)的大脑位点,动物将总是表现出逃避行为。因此,愤怒(RAGE)产生不愉快的情感,而寻求(SEEKING)让人感觉良好。所以,捕食性动物享受扑杀猎物。但它们不享受由愤怒(RAGE)系统过度唤起带来的感受。当然,在所有这些中我们必须记住,拥有更多认知意向性心智的人类也可能比他们希望的更冲动地行动,例如,拿起手枪或其他武器,这太常见地导致他们后来后悔的行为。
总之,所有这些实验发现表明,愤怒(RAGE)和捕食性攻击产生不同的生理反应、行为和情感。重要的是再次强调,关于行为、神经解剖学、脑化学、精神药理学和情感体验差异的大量证据表明,捕食性攻击是寻求(SEEKING)系统的功能,而不是愤怒(RAGE)的表达(Panksepp, 1971)。然而,人类的捕食冲动往往可以以最反社会的方式表达。例如,我们已经提到,假设一些应受谴责的行为,如性跟踪,部分是由认知方向不当的寻求(SEEKING)冲动驱动的,这并不太牵强,但我们不会在此展开这些有争议的想法(但参见Panksepp & Zellner, 2004)。
在攻击性文献中有大量观察结果很难根据参与的情绪类型进行分类。一个特别引人入胜的发现是杀婴行为的案例,这在自然界中非常普遍,在我们人类中也并非罕见。在几乎所有在野外研究过的物种中,尽管不一定在人类中,雄性往往比雌性更多地杀婴。这种行为通常具有繁殖优势:哺乳期雌性往往不排卵,杀死它们的幼崽会迅速恢复性接受能力。
当新的雄狮成为狮群中的统治雄性时,它们的首要行为之一就是”杀死”前任雄性的幼崽;这会使雌性更快地重新进入发情期,有助于确保新雄性自己的血统延续。正如我们将在欲望(LUST)章节中指出的,交配行为本身往往会使具有杀婴倾向的雄性大鼠不太可能杀死幼鼠(Mennella & Moltz, 1988)。随着雄鼠自己幼崽出生时间的临近,这种杀戮冲动会系统性地减弱。这是一个在良好控制的实验室条件下研究得出的真正显著的事实,即使雄鼠不再与交配过的雌鼠在一起,这种情况也会发生。我们推测这种日益增长的和平倾向可能是由某种长期的、依赖经验的表观遗传效应介导的,也许是大脑中催产素传递的促进,尽管目前还没有确定的答案。然而,在这种情况下,我们的观点是,雄性的杀婴行为似乎也是寻求(SEEKING)系统的一种表达。参与杀婴的雄性这样做是为了与雌性交配,顺便提一下,这也是雄性对其他雄性进行攻击的原因之一。
我们不确定这是否与人类行为有关。可能有关。在家庭中,亲生父亲虐待和杀害亲生子女的可能性远低于继父(Daly & Wilson, 2001)。也许这是因为在缺乏稳定社会纽带的情况下,陌生男性更容易对新伴侣的前任子女感到厌烦,这种愤怒的增加导致了令人遗憾的行为。我们只是不知道。无论如何,动物的杀婴行为,就像导致支配等级的雄性间攻击一样,似乎是寻求(SEEKING)系统的表达,而不是愤怒(RAGE)系统的表达。也有可能大脑中存在一个独特的支配(DOMINANCE)系统,但同样可能的是,支配是在其他原始情绪系统如寻求(SEEKING)、愤怒(RAGE)、恐惧(FEAR)和游戏(PLAY)的支持下通过学习产生的。
让我们考虑一下这个在大多数物种中普遍存在但理解不足的心理行为过程。除了捕食性攻击和杀婴行为外,还有另一种攻击性/自信行为不是纯粹愤怒(RAGE)的表现。这就是社会支配的冲动。这种冲动最常见的表达出现在雄性之间,特别是当它们建立领地权并为性优势而相互斗争时。一些人认为支配的冲动是特定类型大脑攻击回路的表达,而愤怒(RAGE)是我们知道存在的主要回路。尽管愤怒(RAGE)通常被用于服务于一般的社会支配和特别是雄性间的攻击,但不应假设支配冲动是愤怒(RAGE)系统的直接表达。尽管在争夺”财产”权的雄性间攻击”竞赛”中——无论是对可消费资源的实际获取、领地还是与雌性的性接触——愤怒(RAGE)肯定会被激起,但有证据表明雄性间攻击和支配冲动与愤怒(RAGE)截然不同。
一些调节雄性间攻击的大脑区域也是传递愤怒(RAGE)冲动的区域(例如,内侧杏仁核和中脑的导水管周围灰质),但对其他区域的损伤(包括下丘脑前区的视前区、外侧隔、伏隔核和中缝核)可以减少雄性间的攻击性,但会增强愤怒(RAGE)。雄性间攻击和愤怒(RAGE)也可以在化学基础上区分。大多数支持雄性间攻击的大脑区域都有高水平的睾酮受体,没有睾酮的雄性表现出更低的支配冲动。我们已经注意到愤怒(RAGE)是一种不愉快的情感,但最近的证据表明,在人类中,睾酮使男性感觉比安慰剂好,但同时男性更不信任他人且更多疑(van Honk et al., 2010)。因此,看来愤怒(RAGE)感觉很糟糕,而睾酮驱动的雄性间攻击冲动感觉良好。因此,虽然愤怒(RAGE)和雄性间攻击可能高度互动,但睾酮在激发愤怒(RAGE)系统方面似乎不是至关重要的(尽管它在某种程度上确实会促进它)。
其他人断言,如此多的动物存在支配倾向表明哺乳动物大脑中必须存在进化提供的支配(DOMINANCE)系统(Ellis & Toronchuk, 2005)。但我们不接受仅凭概念分析的奢侈做法。我们没有足够的证据得出社会支配的冲动来自单一情绪系统的结论。社会支配可能反映了当各种基本情绪系统被激发时发生的学习。换句话说,它在很大程度上是一个次级情绪过程,具有一些初级过程的生物倾向因素。促成因素包括寻求(SEEKING)和愤怒(RAGE),以及恐惧(FEAR),当然早期与粗暴游戏(PLAY)系统的经历也肯定参与其中。
例如,在粗野打闹游戏(rough-and-tumble play)中,幼年大鼠表现出所有在成年大鼠社会支配遭遇中可能看到的行为模式(见第11章)。但当动物玩耍时,这些活动至少最初是在积极情感的背景下进行的。当人类儿童进行粗野打闹游戏时,也能看到类似的支配倾向(想想”山大王”游戏)。实际上,在锦标赛物种中,如鹿,成年雄鹿互相接近的方式就像小鹿渴望玩耍时的样子。当然,雄鹿进行格斗是为了确立雄性优势地位。但考虑到相似的行为,人们不禁会想,成年格斗是否可能是PLAY(玩耍)的成年变体——类似于人类”职业摔跤”和其他目前作为大众娱乐的武术中看到的行为类型。
不幸的是,我们对这类成年行为模式的神经学知之甚少。特别是,我们缺乏必要的数据来证明由局部脑刺激引发的连贯情绪模式,包括奖赏和惩罚特性,而这是我们判断基本过程情绪存在的黄金标准。
尽管如此,为了分享一些关于这种系统的提示性证据,让我们探讨一下替代论点。促进雄性间攻击性增强的一条化学路径涉及从睾酮开始的分子级联反应,睾酮诱导基因表达,激活产生加压素(vasopressin)的基因,加压素是一种促进雄性攻击性和性行为的神经肽(Pedersen, 2004; Veenema & Neumann, 2008)。加压素水平只有正常量一半的去势雄性大鼠,其攻击性和性活跃度远低于加压素水平正常的大鼠。向下丘脑视前区注射睾酮可以恢复大鼠的正常攻击性和性行为水平。这些注射也具有奖赏性,因为动物会对睾酮表现出位置偏好。
有句老话说”有的人总能得到更多”,这似乎适用于睾酮系统:胜利的经历(无论是赢得摔跤或网球比赛,还是从法学院毕业)会产生睾酮分泌增加,从而提高雄性的自信和性行为(Gleason et al., 2009; Strüber et al., 2008)。因此,毫无疑问,大脑具有基于神经的支配型攻击冲动;我们认为这似乎源于几个基本过程脑情绪系统的相互作用,而不是来自我们在本书中讨论的那种情绪”原动力”。尽管如此,睾酮的作用,特别是在年轻青春期男性中的作用,是明确且显著的(Lumia & McGinnis, 2010; van Honk et al., 2010)。值得注意的是,女性激素(雌激素和孕酮,以及催产素)通常会抑制攻击性,因此人们很容易认为女性在气质上通常更平和,而男性更好斗。然而,正如我们上面提到的,女性可以表现愤怒而不进行身体攻击,例如,通过对竞争对手施加社会排斥。可以相信,女性可以使用更多社会手段而非身体手段来施展她们的社会支配地位。然而,在睾酮的作用下,女性人格往往会转向更典型的男性特征(攻击性增强、多疑和性欲增强)。实际上,正如我们将在第7章中看到的,女性的性渴望和性愉悦具有很强的睾酮介导成分(Tuiten et al., 2000),这些”男性”激素通常是由肾上腺睾酮产生提供的。
我们必须暂时得出这样的结论:我们不知道有任何明确的大脑机制能够证实在女性中促进支配行为的基本过程力量,尽管睾酮显然可以做到这一点。在自然界中,这在某些物种中非常明显。考虑雌性斑鬣狗,它们的睾酮水平异常高(在第7章中进一步讨论)。雌性鬣狗比雄性更具攻击性,它们使用增大的、类似阴茎的阴蒂主要用于社会性交流,特别是支配展示。雌性中的高睾酮水平似乎是其攻击性和支配行为的原因。也许高水平的睾酮也会促进新生鬣狗的类似攻击性/支配行为,它们通常以双胞胎的形式出生。它们出生时就带有战斗情绪,两只中的一只通常在进入以友好打闹游戏为特征的温和青年期之前死亡。这种攻击行为也可能是RAGE(愤怒)系统的表现,但我们必须保留这样的可能性,即它反映了支配欲望的早期表达。显然,关于各种形式的攻击性如何在神经系统中展开,我们还有很多需要学习。
不难想象 RAGE(愤怒)、捕食性攻击和社会支配是如何在人类心理的三级过程层面交织在一起的。以职业网球运动员为例,他们一年中大部分时间从一场比赛转战到另一场比赛。他们彼此非常了解。有些成为朋友,有些则不那么亲密——但迟早他们会在球场上成为对手。当朋友之间相互比赛时(甚至是姐妹之间,如塞雷娜·威廉姆斯和维纳斯·威廉姆斯),他们是在争夺一种社会支配地位。也许对手也可能扮演猎物的角色,这突显了 SEEKING(寻求)在这类对抗中的作用。当然,其他动物热衷于捕杀猎物以享用美餐,而运动员则热衷于击败对手以享受社会胜利。然而,在这两种情况下,SEEKING 系统无疑都被激活了。
也许在一场艰苦战斗的激烈时刻,即使对手是朋友或姐妹,人们也可能会对对手感到瞬间的 RAGE。人们经常听到运动员谈论获胜所必需的”杀手本能”。在某种程度上,杀手本能是 RAGE 的一种表达,特别是在挫折和即将胜利的时刻。然而,杀手本能可能也源于捕食性攻击、习得的社会支配欲望,以及认知介导的成为顶尖选手的愿望。因此,很容易看出这三种生物学促成的攻击形式——RAGE、捕食性攻击和社会支配——是如何在三级过程层面的高级心智中融合的。这可能就是为什么我们难以将所有这些感受理解为独立的、基本的情感概念。只有当这些概念得到大量可靠神经科学证据的证实时,依赖初级过程情感概念才具有科学有效性。尽管目前的研究表明,RAGE、捕食性攻击和社会支配欲望在神经生物学上在某种程度上是不同的,但其中只有第一种似乎是一个专门用于初级过程攻击形式的独立情感系统。
我们从解剖学和化学的角度描述了 RAGE 系统。我们还描述了 RAGEful 行为在人和动物身上是如何表现的。特别重要的一点是,尽管击败对手可能会带来即时的快感,但 RAGE 系统会产生不愉快的情感。这是每个心理健康专业人士都应该牢记的一点。尽管男性往往比女性更具攻击性,但这仅适用于身体攻击。而对于女性大量参与的心理社会攻击的微妙方面,我们了解得要少得多。
我们将 RAGE 与捕食性攻击和杀婴行为区分开来,后两者似乎都是 SEEKING 系统的表现形式。我们还讨论了社会支配欲望,其神经基础尚不完全清楚。然而,我们不认为社会支配欲望反映了单一初级过程系统的存在。支配行为可能是多个情感系统被激活时发生的学习结果。当然,支配行为在儿童玩耍时出现,在我们祖先的环境中,当狩猎采集者由大家庭组成时,年轻人之间的这类活动很容易形成持续到成年的自然支配等级。还有一个考虑因素是,雄性之间的支配似乎受到睾酮和加压素的推动,这将其与 LUST(性欲)系统联系起来。
有一点非常清楚。暴力犯罪是一个规模巨大的社会问题,这突显了对能够抑制 RAGE 的药物的需求(目前需要在人类身上评估 P 物质拮抗剂如阿瑞匹坦)。然而,心理治疗师和精神科医生也应该记住情感系统的相互作用,并理解当人们(特别是儿童)遭受虐待和忽视时,RAGE 会被强化。从病理性 RAGE 中恢复的关键是建立或重新建立一个人形成和维持温暖信任关系的能力。持续友好和积极的互动对愤怒的灵魂可以产生绝佳的安抚效果(想想那些患有隔膜损伤的动物,随着时间的推移和亲社会经验变得温顺)。同样,治疗环境中的积极情感体验可能有助于减弱许多种令人烦恼的记忆的锋芒。心理治疗可以帮助患者摆脱那些否则会演变为消极和令人恼怒的反刍思维的问题。威廉·布莱克(1793)在他深刻热情和人文主义的诗歌中注意到了这一点;例如,在《毒树》中他写道:
我对朋友生气。
我说出愤怒,愤怒就消失了。
我对敌人生气:
我没说出来,愤怒就增长了。
这在今天仍然是真实的——似乎是一种普遍的人类体验——反映了我们的高级心智如何与我们初级过程的 RAGE 潜能相互作用。
在他短暂的洞穴生活中,他从未遇到过任何值得害怕的东西。然而恐惧就在他心中。它通过一千次生命从遥远的祖先传给他。这是他直接接受的遗产……通过之前所有世代的狼传承下来。恐惧!——那是野性的遗产,没有任何动物能逃脱……所以灰色幼崽知道恐惧,尽管他不知道恐惧是由什么构成的。
——杰克·伦敦,《白牙》(第52页)
在本章中,我们将讨论恐惧的本质——富兰克林·D·罗斯福总统在1933年3月4日的著名演讲中强调的原始恐怖,当时他告诫国家:“我们唯一需要恐惧的就是恐惧本身——无名的、非理性的、毫无根据的恐怖,它会麻痹将撤退转化为前进所需的努力。”这些预言性的话语是在美国开始从大萧条中爬出来的那一年说出的……也是阿道夫·希特勒在德国上台的那一年(这给数百万人带来了深刻的恐惧和苦难)。
如果我们已经经历过可怕的经历,我们就会学会恐惧恐惧本身。在历史上的所有战争中,年轻的战士都感受到了周围死亡的恐惧。这种恐惧持续的时间越长,它就越深地刻在他们大脑的突触(synapses)中,有时会使他们毫无意义地、痛苦地、永久地害怕一切——从某种意义上说,又什么都不怕。慢性焦虑的无对象恐惧直接来自他们过度活跃的初级过程恐惧系统,而不是来自他们当前处境的实际现实。对我们大多数人来说,很难想象这种”无对象”的恐惧,但这就是我们的恐惧系统可以产生的那种自由漂浮的慢性焦虑。这个系统,像所有情绪系统一样,表现得就像我们身体的肌腱和肌肉。使用得越多,它们就越强;使用得越少,它们就越弱。二十世纪大战中的许多士兵都经历过”炮弹休克”,现在被称为创伤后应激障碍(PTSD)——恐惧感作为灵魂中永远存在的刺激而逐渐渗透,许多可怕的图像被刻在心灵的记忆表面。所有哺乳动物都可能患上PTSD,因为我们都有非常相似的古老恐惧系统,在我们核心情感意识的认知黑暗中,这些系统可能会变得敏感和充满恐惧。
当我们受到足够严重或足够长时间的惊吓时,恐惧系统可能会变得过度敏感。从出生起,这种自由漂浮的恐惧能力就内置在我们的大脑中;最初它只能被少数非条件刺激激活,但经验可以创造出恐惧记忆,从此可以被世界上以前中性的事件触发。恐惧,像其他所有情绪系统一样,本质上是”无对象”的诞生,并且像脑心(BrainMind)的所有其他情绪系统一样,它通过学习与现实世界建立联系。显然,老鼠仅仅能够感到害怕是不够的。它必须学会害怕各种特定的对象和情况。我们也是如此。进化在大脑中创造了恐惧的能力,但它没有(也不能)告诉我们所有我们可能需要恐惧和避免的事情。实际上所有这些都必须学习。而且因为我们如此聪明,我们人类可以学会害怕比小老鼠更多的事物,无论过去还是未来(见下一章的题词,罗伯特·彭斯深情地描绘了老鼠和人类恐惧的差异)。从多种意义上说,我们人类是地球上最恐惧的生物。我们可以为自己创造超出任何其他物种想象的恐惧。由于我们的新皮质能力,我们甚至开始害怕心灵中虚无缥缈的幻影。但我们并不太清楚这种内在学习是如何发生的。正如我们将在下一章中讨论的,然而,我们确实对恐惧学习的最简单形式如何在大脑中发生了解很多。
我们不必深入思考就能在生活中找到被恐惧感吞噬的时刻,尤其是在我们年轻的时候。即使我们较高的认知思维可以轻易地引导我们认识到我们没有面临真正的威胁,我们也经常忍受这样的状态。我们甚至可能因为变得焦虑而焦虑。我们不知道其他动物是否能够产生这种二阶的、自我产生的预期焦虑。但它们的恐惧系统——像我们的一样——被设计用来预测未来的坏事,如果它们反复受到创伤,它们肯定会在各种令人生畏的情况下变得敏感和过度活跃。换句话说,我们知道大脑的恐惧网络在所有哺乳动物物种中都可能反应过度,就像我们大脑的所有其他基本情绪过程一样。
想象一下,你独自一人,在夜色笼罩下的树林中迷了路(见[图5.1])。你在远足途中不小心迷路,对自己找到出路的能力信心不足。月光透过疾驰的云层洒下,伴随着凛冽的寒风。头顶的树枝不祥地摇摆着。你的想象力开始狂奔,浮现出各种恐怖景象,从猛兽到梦中的妖魔。这些幻象就像你童年时熄灯后想象中的怪物一样可怕,即使安全地躺在床上……但往往是孤身一人(因为与母亲同睡这种所有其他灵长类动物都会做的事,在我们人类中已经过时了)。突然,一根树枝在你身后断裂掉落。如果你在自家后院听到这个声音,可能只会略感惊讶地转过身去。但因为你已经处于恐惧状态,你经历了一次剧烈的惊吓。你屏住呼吸,僵在原地一动不动,脑海中充满恐惧。你所有的感官都集中在声音的来源处,快速分析可能的来源。是美洲狮要扑过来吗?是蝙蝠在头顶盘旋吗?在恐惧的谵妄中,你甚至可能想象出神话中的狼人。如果你感到迫在眉睫的危险,可能会猛然爆发出剧烈的逃跑动作,跑得比你以为自己的双腿所能承受的速度还要快。如果你足够幸运找到了一个安全的地方(也许是树林中一座废弃的小屋),你会躲起来,心跳加速,浑身颤抖(不仅仅是因为体力消耗;恐惧总是伴随着自主神经系统的唤醒)。你可能在途中尿了裤子,或者更糟。你会在冷汗中长时间保持警觉,警惕地评估每一个新的声音,每一个可能表示危险的阴影。

图5.1. 典型恐惧序列的漫画:在黑夜的树林中迷路,任何突然的声音都会让人僵住,想象中的妖怪会产生许多可怕的可能性,导致逃跑。即使找到废弃小屋的避难所,仍处于焦虑唤醒状态,这种状态可能会在随后许多个夜晚的梦中重现(这幅漫画最初由Sandra Paulsen为本书绘制,并由Lonnie Rosenberg精修)。
幸运的是,在黎明时分你找到了出路。以后你会更加小心,不再迷路。你可能会连续几个晚上梦到这段经历。如果你真的遇到了美洲狮或狼獾,你的恐惧行为可能是适应性的。如果你尖叫着像个疯子一样四处奔跑,特别是来回挥舞双手,你可能会吓跑那只动物。在这次磨难之后,纯粹的恐惧和各种相关的想法可能会在你心理的神经基质中酝酿——也许会持续多年。你甚至可能发展出轻度的创伤后应激障碍(PTSD)。当这样的情绪系统变得过度敏感时,即使你在认知上知道自己是安全的,你也可能经历”恐惧本身”的使人衰弱的痛苦。
恐惧在所有形式上都是痛苦的。突然被恐怖袭击是可怕的。被持续的焦虑感不断吞噬,摧毁你对世界的安全感,也同样可怕。这些感觉是由一个协调运作的原始大脑系统产生的,从导水管周围灰质(PAG)到杏仁核(amygdala),再回到导水管周围灰质。当这个系统被突然唤醒时,会产生恐怖感;当受到较温和、更持续的唤醒时,会促进慢性焦虑。当恐惧刺激距离较远时,大脑的高级认知部分,如内侧前额叶皮质和杏仁核,也会被唤醒;你可能会躲藏并保持静止。但当可怕的捕食者在你身后紧追不舍时,恐惧回路的较低区域,特别是中脑导水管周围灰质,会接管控制(Mobbs等,2007)。那些无条件的恐惧回路绝对会迫使你逃跑。
许多人仍然认为恐惧能力是习得的,人和动物都是通过预期危险来学会恐惧的。如果这是真的,我们出生时就不会害怕任何东西。只有在某种方式受到伤害后,我们才会知道恐惧意味着什么。但动物即使从未经历过疼痛或危险,也表现出与生俱来的恐惧能力。我们知道这一点,是因为如下一节所述,对大脑特定部位的电刺激可以在完全安全环境中饲养的动物身上产生全方位的恐惧反应。电刺激并不携带关于环境中危险或疼痛的身体刺痛的任何信息。直接刺激只是唤醒了恐惧系统的内在情感潜能——它唤醒了恐惧本身。
这一点本应在半个多世纪前的1950年代和1960年代就被研究人员更好地认识到。当时他们最初发现了大脑中不仅有动物会自愿自我刺激的区域([第3章]),还有其他邻近区域,刺激这些区域会让动物像躲避心理瘟疫一样逃离——这些区域甚至会激发习得性逃避行为(Delgado et al., 1954)。在研究过程中,这些研究人员偶然发现了原始的恐惧系统,该系统贯穿许多动物自我刺激的大脑区域附近。当这些恐惧结构被无意中刺激时,动物表现出各种恐惧行为,在低电流水平下冻结,当电流增加时则仓皇逃窜。因此,早在恐惧系统被正式概念化(Panksepp, 1982)之前,就可以推测实验室大鼠具有先天的恐惧系统。这一点很明显,因为大鼠害怕某些类型的大脑刺激,而这与对外周厌恶性事件(如常用的足部电击)的学习无关。所有在这些区域受到刺激的哺乳动物都表现出恐惧行为。显然,这个系统是由进化而非动物的生活经验所建立的。
很久以前,在早期脊椎动物大脑进化过程中,对某些威胁性外部刺激的识别被编码进我们祖先的大脑构建DNA中,产生了对某些持续造成疼痛或预示危险的刺激的先天恐惧。例如,大鼠天生害怕某些捕食者的气味,如猫、雪貂和狐狸。它们最初并不害怕这些捕食者的外观,只害怕它们气味的某些方面。如果将这些生物的毛发放入在受控实验室环境的完全安全中长大的大鼠或小鼠笼中——这些动物一生中从未遇到过任何捕食者——它们仍然会表现出恐惧反应。许多动物会简单地冻结;其他动物则表现出普遍的警惕(研究人员称之为”风险评估”,即此类情绪的认知担忧类型方面)。即使在这些可怕的气味被移除后,大鼠和小鼠也会长时间保持胆怯,这是由于它们大脑内释放的一系列恐惧神经化学物质。大鼠的社交活动会被抑制相当长一段时间,它们在玩耍、进食、梳理、性行为和其他积极行为方面的参与会减少(相关数据描述见下一章的[图6.1])。如果动物长时间受到这些压力源的影响,它们会开始表现出抑郁症状。这种天生害怕捕食者气味的能力促进了生存,因为遗传的恐惧系统激励动物在捕食者附近时冻结和躲藏,如果捕食者靠得太近就逃跑。从进化的角度来看,与捕食者的一次无所畏惧的相遇就已经太多了。
对来自捕食者的气味的恐惧帮助动物避开捕食者居住的地点,而不是捕食者就在附近的信号。我们可以得出这个结论,因为捕食性气味是通过它们的犁鼻器进入啮齿动物大脑的,该器官检测大的非挥发性分子,而不是通过监测相对远处”随风飘来”气味的主要嗅球(Panksepp & Crepeau, 1990)。小鼠也是如此。这种令人反感的气味的分子组成最近在猫唾液中被识别出来;结果证明是属于主要尿蛋白家族的单一分子,称为Feld4(Papes, et al., 2010),该分子此前已被同一组研究人员确认可以加剧小鼠雄性之间的打斗。据推测,小鼠很容易与陌生者打斗,部分原因是它们携带的气味使动物彼此警惕。
除了害怕疼痛和捕食者的气味外,大鼠天生害怕光线充足的开放空间、突然的移动和巨大的噪音。所有这些刺激都表明可能存在危险,它们作为进化记忆(即硬连接到恐惧系统的感觉输入)被传承下来,因为大鼠天生害怕它们是具有适应性的。在所有哺乳动物物种中都存在一些引起先天恐惧的刺激。疼痛是普遍的诱因。大多数动物在听到巨大噪音时也会变得害怕。人类婴儿在没有被安全抱住时会变得焦虑,随着年龄增长,许多婴儿在黑暗中独处时往往会哭泣。这些负面感受可能与社交恐慌/悲伤系统([第9章])同样来自恐惧。事实上,如果没有大脑研究,可能很难区分这些”焦虑”系统中的哪一个更活跃。它们有可能同时活跃,但这些问题尚未被神经科学家研究。在本章中,我们将只关注恐惧系统。像所有其他初级过程情绪系统一样,它生来相对”无对象”,但哺乳动物可以迅速学会对许多预示恐惧诱发条件的刺激做出反应。
所有幼年动物最初只对少数几种内在输入产生恐惧反应,其中疼痛是最被理解的。疼痛刺激直接进入导水管周围灰质(PAG),因此那里也存在疼痛抑制机制。在PAG的适当位置给予刺激,可以缓解人类相当严重的疼痛(Mayer et al., 1971; Richardson & Akil, 1977),这是因为内源性阿片类物质的释放(Hosobuchi et al., 1979; Herman & Panksepp, 1981)。然而,如果刺激到PAG中的恐惧位点,动物就会表现出完整的恐惧反应。这种恐惧状态可能促进学习,这可能是动物快速发展出对捕食者相关的视觉和听觉刺激的习得性恐惧反应的方式(见下一章,专门讨论恐惧学习和记忆)。通过这种方式,恐惧系统在简单学习(二级过程)和更复杂的认知(三级过程)层面上都受到大量生活事件的控制。
人类和其他动物表现出自由浮动焦虑的事实表明,他们具有体验恐惧的内在能力。换句话说,变得焦虑的能力是大脑进化情绪工具箱的一部分。正如我们已经提到的,证明这一点的简单事实是,仅仅通过电刺激或化学刺激特定的大脑区域,就可以轻易引发一整套行为和生理恐惧反应。这种反应在所有被研究的哺乳动物中都很明显。根据几乎所有测量指标,动物都不喜欢这种感觉——它们试图逃离刺激,避开发生过这种刺激的地方,等等。当然,学习可以为恐惧系统增添很多内容(见下一章),但我们的关键点是,学习本身并不能解释恐惧能力。这种基本能力是由大脑中固有的情绪系统提供的。
只有当恐惧系统本身以某种方式被破坏,或者感觉输入进入该系统的通路以某种方式被阻断时,恐惧能力才能被消除或减弱。这可能通过损伤或疾病发生。例如,有些寄生虫(如弓形虫,一种常见于猫身上的寄生虫)可以攻击啮齿动物的恐惧系统,使老鼠对猫的恐惧减少(Vyas et al., 2007)。这促进了猫科动物的捕食。猫吃掉更多老鼠,因为它们的猎物不像平常那样容易躲藏和逃跑。所以受感染的啮齿动物进入猫的胃里。而猫的身体是原生动物完成其繁殖周期的完美环境。
恐惧系统存在的最直接证据来自使用大脑直接电刺激的实验(最近的综述,见Panksepp et al., 2011)。当电刺激应用于哺乳动物大脑的特定部位,即所有哺乳动物共有的深层皮层下区域时,即使环境中没有令人恐惧的刺激,动物也会表现出先天的恐惧反应。不同强度的电刺激产生不同程度的恐惧。轻度电刺激产生警惕、抑制的活动,偶尔出现冻结不动的现象,类似于老鼠笼子里放置猫毛时表现出的抑制行为——这些行为在捕食者可能还很远时很常见。如果在同一大脑位点进一步加大电流,动物会逃跑,就像猫靠得太近准备攻击时老鼠的反应一样(Panksepp, 1991)。随着这些进展,神经唤醒的级联从较轻的恐惧形式(冻结、担忧)转向更强烈的形式(逃跑、恐怖)。
在人类中,当恐惧系统被内部物理刺激唤醒时,例如边缘系统中该情绪网络所在部位的癫痫活动,已观察到恐惧感增加。癫痫是大脑中的电风暴。当这种风暴侵入恐惧系统时,人(或动物)表现出强烈的内在恐惧,可能在某种程度上感觉类似于创伤后应激障碍(PTSD)(Adamec, 2001; Pincus, 1981, 2001)。因此,癫痫发作产生的电流可以以非常类似于实验室中直接电刺激恐惧回路的方式起作用。
电刺激实验揭示,先天恐惧系统是一条双向通路,从杏仁核的中央区域延伸到下丘脑的前部和内侧部分,围绕第三脑室,然后到达中脑导水管周围灰质(PAG)内的特定(背侧位置)区域(见图5.2)。恐惧系统包括特定的自主神经和行为输出,控制恐惧的生理症状(如手心出汗、心跳加速、冻结或逃跑)。药理学和手术抑制沿该系统的活动可以使动物和人类变得温顺。简而言之,许多无条件的——本能的——恐惧表达直接来自这个神经系统。
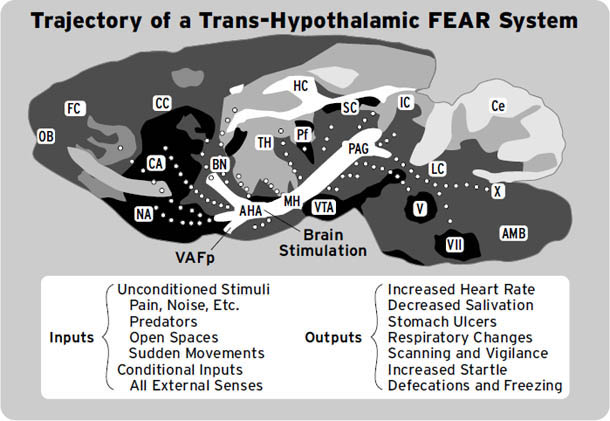
图5.2. 恐惧系统轨迹及刺激该情绪系统诱发的各种症状的示意图(改编自Panksepp, 1990b; 解剖学标记见原文)。大脑区域的深浅程度近似表示大脑中乙酰胆碱的水平。
许多实验表明,动物非常厌恶这种大脑唤醒。动物会试图逃避它。如果有机会,它们会迅速学会通过按压杠杆或简单地移动到从不发生刺激的位置来关闭这种刺激(Panksepp, 1991)。它们还表现出条件性位置回避(conditioned place avoidance):它们会避开曾经接受过刺激的地方(Roberts & Cox, 1987)。如果动物暴露在发生过这种大脑刺激的环境中,当有机会选择时,它们会避免回到那些位置。但即使它们在附近的安全区域,它们看起来仍然会紧张,经常比正常情况下更多地僵住和排便——因此,仍然表现出恐惧的行为和自主神经症状(Panksepp, 1991)。
因此,恐惧情感状态的客观关联物是这种刺激产生的视觉上明显的僵住和逃跑。再次强调,大脑刺激引发的明显行为是一个客观等价物;它是我们无法轻易观察到的心理状态的外部指标。然而,根据情感神经科学策略,类似的大脑刺激应该在人类身上引发被唤起的情绪状态的自发语言指标。事实上,这已经被一致观察到。
在这些大脑区域受到刺激的人类持续报告突然出现的恐惧和焦虑。例如,当对导水管周围灰质(PAG)的刺激开启时,最初的一位受试者说:“我吓死了”(Nashold et al., 1969)。在另一项观察人类脑室周围灰质电刺激后心理变化的研究中(Amano et al., 1979),患者报告”突然感到不确定,就像进入一个又长又黑的隧道”,一种在海边”海浪从四面八方涌来”的感觉,以及”有人正在追我。我正试图逃离他。“恐惧系统的唤醒迅速在皮层触发了充满焦虑的场景,也许来自真实的过去经历,或者只是来自记忆中的故事。这种互动的速度使人们很容易理解为什么人们会认为感觉是在皮层中产生的——认为想象这些场景的能力对于产生感觉是必要的。但请记住,最初的恐惧感来自对更深层情绪系统的刺激,而这个系统为所有哺乳动物所共有,无论它们的认知天赋如何。
疼痛总是在某种程度上唤醒恐惧系统,但反之则不然。恐惧实际上可以减弱对疼痛的感知(Miczek, 1991)。当人类大脑中的恐惧系统受到电刺激时,人们报告恐惧但不报告疼痛。当这个系统在动物身上受到电刺激时,它们表现出恐惧,但很少像真正受伤时那样尖叫或吠叫。然而,强烈的恐惧常常可以抑制疼痛的体验,因为在恐惧发作期间,大脑会分泌镇痛的大脑化学物质,如大脑自身的阿片类物质(opioids),暂时减轻疼痛感觉(Miczek, 1991)。这是一种适应性机制,允许受伤的动物忽略疼痛,增加它们可能逃离捕食者的可能性。然而,它也可能导致伴随创伤后应激障碍(PTSD)的麻木。有一些证据表明,阻断阿片受体实际上可以减少这种麻木和心理解离,帮助边缘型人格障碍患者对心理治疗做出更积极的反应(Bohus et al., 1999)。这同样适用于创伤后应激障碍(Pitman et al., 1990)。
虽然突然的疼痛是一种通常可以唤醒恐惧系统的刺激,但我们刚刚看到,该系统也可以很容易地被不会引起身体疼痛的刺激唤醒。捕食者的气味不会给老鼠造成身体疼痛。光线充足的开放空间不会造成身体疼痛。同样,如果人类婴儿在身体上没有得到良好的支撑,它最终可能会摔倒并受伤,但在经历任何身体疼痛之前,表面上的支撑缺失就会唤醒恐惧。巨大的噪音可能令人不快,但它们很少是疼痛的。然而,婴儿和大多数动物都害怕雷鸣般的或刺耳的声音,因为这些”惊吓”刺激在大多数哺乳动物物种的进化历史中经常预示着危险事件。事实上,如果动物已经焦虑,惊吓反应会被放大。人们早就知道,焦虑的气质特征可以通过使用行为遗传选择程序很容易地培育到动物身上。研究人员开始详细说明这种遗传气质产生的大脑变化(Harro, 2010; Harro et al., 2011; Kanarik et al., 2010; Singewald, 2007)。
身体疼痛经常用于恐惧条件反射实验,因为它很容易施加给实验室动物,最常见的是通过施加电击。在恐惧条件反射中,当呈现与总是唤醒动物恐惧系统的非条件刺激(如电击)配对时,动物学会害怕条件刺激(以前的中性刺激),如听觉音调或光线,这在人类身上也是如此。很快,即使没有电击伴随,动物也会学会害怕音调或光线。换句话说,预测疼痛事件的线索总是开始在几乎所有被研究过的动物身上产生恐惧反应。对条件刺激的恐惧反应的快速发展是成功恐惧条件反射的标志(见下一章)。
很少有从事此类工作的行为神经科学家愿意承认,甚至谈论他们的实验动物是否经历了任何可怕的事情。他们声称,有时显然是出于机会主义,这种内在感受无法直接观察到,因此应该被排除在科学讨论之外。然而,这似乎是短视的。事实上,如果厌恶感受是动物大脑学会在行为上产生恐惧的关键原因,这些科学家就永远无法理解他们如此感兴趣的恐惧条件反射究竟是如何运作的。科学家应该遵循”证据的权重”,但在这个领域,科学家的这一标准价值似乎被忽视了。这就是为什么我们强调了一种双面一元论(dual-aspect monism)策略(Panksepp, 2005b, 2007a),它有能力将动物的情绪感受转化为人类的具体心理预测。
除了对条件刺激产生恐惧之外,大鼠很容易对在条件反射体验期间恰好存在的各种背景(外部)刺激产生恐惧。例如,大鼠很容易学会害怕与电击配对的音调,但它们也会害怕条件反射箱的墙壁,也许还会害怕那些测试笼子中使用的木屑的独特气味。大鼠还可能对把它们放入测试箱的实验者的外观、声音和气味产生恐惧。这些都是在系统性恐惧条件反射实验中也被纳入条件反射范围的背景刺激。
研究人员热衷于了解恐惧系统的化学机制,主要是出于精神病学的原因。例如,许多患有创伤后应激障碍(PTSD)的人可以通过缓解其强烈恐惧感的药物得到帮助。四种一般类型的实验被用来研究恐惧系统的化学机制。每一种都涉及诱导恐惧的方法和测量对特定药物的恐惧行为减少的方法。假设任何减少动物恐惧反应的潜伏期(持续时间)或强度的药物也可能减少人类的恐惧情感。这项实验工作揭示,在大多数情况下,苯二氮卓类药物(benzodiazepines)[1](BZs)能够平息大多数类型的恐惧。
我们不会在这里详细研究为研究动物恐惧而设计的各种实验程序。如果读者感兴趣,可以参考之前对这些问题的详细介绍(见Panksepp, 1998a, pp. 209-212),本简化版本就是基于此。理解这些方法对于该领域的工作者比对普通读者更重要。然而,一些读者可能希望了解各种药物在涉及条件性情绪反应(conditioned emotional responses, CER)、增强惊吓反应(potentiated startle responses)以及诸如高架迷宫(elevated mazes)等本质上令人恐惧的环境的实验中改善恐惧的详细效果,所有这些都利用了动物的先天防御行为。例如,你可以在大鼠笼子里放置一个带电击的探针。迟早,在探索过程中,大鼠会触碰到探针,通常是用鼻子,并会受到不受欢迎的电击。通常,大鼠会在笼子里堆积木屑或其他垫料,试图覆盖探针。(这是大鼠的本能防御行为还是基于先前学习的行为尚不清楚。)如果由于药物作用,大鼠需要更长时间才能在冒犯性探针上建立屏障,则该药物被视为减少了大鼠的恐惧。同样,抗焦虑药物如苯二氮卓类药物在减少这些和许多其他防御行为方面是有效的。然而,正如我们将看到的,它们在减少分离恐慌反应方面并不特别有效,这只是表明它是一个独特的负面情绪系统的众多证据之一,它引发了一种不同类型的”焦虑”(见第9章)。
我们不会对与药物相关的实验研究进行全面的检查,但我们想详细讨论两个主题。第一个是恐惧系统影响惊吓反应的方式。所有动物都对大的噪音表现出惊吓反应。然而,这种反应的强度可能会有所不同。如果恐惧系统已经被唤起,惊吓反射将会强烈得多。例如,如果你让一只实验上未经训练的大鼠暴露于大的噪音,该大鼠可能会对那个非条件刺激表现出适度的惊吓。然而,假设你之前已经训练大鼠将光线与足部电击联系起来。那只动物已经学会害怕光线。如果你让条件反射后的大鼠暴露于光线下,从而唤起恐惧系统的背景水平的唤醒,然后不久之后让大鼠暴露于同样的大噪音,它的惊吓反应将远远大于如果它没有被训练害怕光线的情况。顺便说一下,这就是为什么当你偷偷接近正在看恐怖电影的人时,通常可以引发”增强惊吓”或极端反应。因为她已经处于恐惧状态,所以这个人的惊吓反应比她看喜剧时更加剧烈。
这种”增强”的神经学细节已经被阐明。基本上,惊吓反射本身在神经系统中组织得非常低层,作为一种快速反应的反射,远低于恐惧回路。然而,恐惧回路的输出确实会下降到那个层次,如果恐惧被唤起,它会促进脑干内这种古老反射的强度。这是观察恐惧系统如何增强特定反射的绝佳方式。类似的程序已用于人类,通过对眼睛施加一小股气流引发恐惧增强的眨眼反应(Davis & Lang, 2003)。
第二个我们将简要探讨的话题涉及与这些药物实验相关的一些概念和方法学问题。这些问题对某些药物的疗效产生了一些混淆。认为任何能够减少明显恐惧行为的化学物质就必然减少恐惧情感,这是一个错误。假设动物笼子里的一个杠杆会传递疼痛的电击。在第一次或第二次电击后,动物会避免触碰杠杆。如果你给动物一种诱发遗忘的药物,它将不再避免触碰杠杆,因为它会忘记杠杆是痛苦的来源。所以这种药物会增加受惩罚的行为,但它不会通过减少动物每次电击后的情感痛苦来做到这一点。这是动物愿意参与受惩罚行为并不反映恐惧减少的一个例子。其他药物可能只是解除动物的抑制,使它们更活跃,愿意做更多事情。这类药物也可能增加随机按压杠杆的行为。当测试一种增加动物愿意参与受惩罚行为的特定药物时,研究人员总是需要考虑该药物可能影响除焦虑减少之外的大脑过程。这些限制适用于所有其他情绪系统的研究。这就是将情感感受排除在科学话语之外的危险。
在血清素(serotonin)研究中,有一个领域将一般性的去抑制误认为恐惧的减弱。在1950年代到1970年代之间,一些科学家确信大脑中血清素活动的增加是焦虑的原因。得出这一结论是因为血清素受体拮抗剂(antagonists)减少了大脑中的血清素活动,导致动物参与更多受惩罚的行为,比如按压传递食物的杠杆,即使在呈现预测即将到来的足部电击的CER刺激时也是如此。因此这些研究人员认为血清素的减少肯定减少了焦虑,他们得出结论认为高水平的血清素会导致人类和动物感到焦虑。
然而,现在已经清楚的是,大脑中血清素的减少会使动物总体上变得更加躁动和冲动。血清素在大脑的大部分区域全局性地发挥作用。血清素被减少的动物往往在广泛的情况下被解除抑制,它们在引发焦虑的情况下倾向于过度反应,这是因为它们的冲动性,而不是因为焦虑的真正减少。事实上,血清素耗竭的动物往往比正常动物更容易焦虑,并且在所有领域通常都是超情绪化的;例如,它们往往表现出比正常情况多得多的攻击性,并且经常表现出性欲亢进。因此,在面对惩罚时行为的增加可能只是反映了主动行为倾向的普遍释放或去抑制,而不是焦虑感受的减少。
尽管血清素调节焦虑的强度,但它调节其他负面情绪的程度并不比调节焦虑更大。血清素调节所有情绪的强度。大脑血清素活动的升高通常抑制情绪,包括恐惧,而较少的血清素则唤起情绪,包括恐惧。因此,当血清素缺乏的动物表现出受惩罚行为的增加时,是因为它们所有的情绪都被唤起,而且它们过度活跃。目前几乎没有经验性理由相信大脑中血清素活动的全局性升高在促进焦虑或恐惧体验方面起主要作用。目前,大多数可用数据更符合替代性结论,即神经元之间突触中血清素活动的整体增加会减少焦虑并产生放松的感觉——血清素可以抑制大脑中的每一种情绪和动机冲动。这就是为什么选择性血清素再摄取抑制剂(SSRI)抗抑郁药,增加突触中血清素的可用性,在放松压力过大的人群和减少他们的易怒性方面相当有效(Knutson, Wolkowitz et al., 1998)。
然而,在过去几十年中,已经确定了大量血清素受体类型(目前已知有15种),看起来其中一两种受体实际上可能会促进某种负面感受。尽管如此,不同血清素受体如何精确地参与放松的产生以及促进负面情感,目前还远未得到充分理解。例如,一种相对较新的抗焦虑药物丁螺环酮(buspirone)(其品牌名称是BuSpar),已知作用于血清素受体,这些受体可以增加或减少大脑血清素活动,这取决于受体相对于突触的位置。最初,丁螺环酮被认为通过减少突触前末梢的血清素释放来减少焦虑,但现在看来更可能是通过增加一种突触后受体的血清素活动来实现的(详见Panksepp, 2004, p. 501)。因此,尽管很明显血清素活动的全局性促进会减少焦虑,但关于血清素在个别受体上的作用仍有很多需要学习的地方。
总之,有许多方法可以监测动物的恐惧感——从在称为”开放场地”的大型测试区域中的胆怯行为,到”社交互动测试”、“高架十字迷宫”以及”情境性僵立”。这些环境和行为如何与恐惧系统联系起来仍不确定。不过总体而言,目前关于恐惧学习的研究工作量巨大(如下一章所述),而关于大脑进化提供的恐惧回路的研究工作却很少。因此,虽然神经科学家对允许条件刺激进入恐惧系统的神经化学机制(例如谷氨酸突触)了解很多,但他们对恐惧系统本身的工作方式了解相对较少。尽管如此,在过去几十年中,巴西的几个研究小组一直在深入研究中脑导水管周围灰质中调节防御行为的各种神经化学机制,用我们的术语来说就是恐惧系统(例如,Brandão等人,2003年,2008年;Del-Ben & Graeff,2009年)。其中一些细节难以简洁地总结,但读者应该相信,对这个系统的神经化学理解将为药物开发提供许多可能性,包括简单的操作,如减少介导某种焦虑形式的回路中的炎症级联反应,这种焦虑可能类似于社会分离痛苦的感受(见第9章),在成瘾后吗啡戒断时会引发这种感受(例如,Hao等人,2010年)。
并非每种形式的焦虑都来自恐惧系统。我们在多种情境中使用”焦虑”这个词,但我们现在知道”分离焦虑”在大脑中是一种与本章迄今所描述的各种情绪恐惧非常不同的过程。心理治疗师和科学精神病学专家认识到大脑中存在几个不同的负面情绪系统,并且在任何给定时间可能不止一个系统被激活,这一点很重要。这些系统如何相互作用仍然未知。然而,成功的治疗很可能依赖于理解在每个患者中哪个系统占主导地位。例如,第9章详述的恐慌/悲伤系统,对于人们在经历”恐慌发作”时所产生的强烈社交不安全感和失落感,可能比对可怕的非社交事件产生的预期焦虑更为重要。
我们将用整整一章来讨论恐慌/悲伤系统,但为了扩展之前的评论,我们注意到有两个很好的理由来区分恐慌/悲伤系统和恐惧系统。首先,它们由不同的大脑结构支持,因此在解剖学上是不同的。其次,恐惧和恐慌/悲伤系统在某种程度上由不同的大脑化学物质控制,并且对药物有不同的反应。正如我们所看到的,苯二氮卓类药物(BZs)通常能有效平息恐惧,但在消除幼年动物与父母分离时发出的痛苦叫声方面几乎没有效果。最初的苯二氮卓类药物(利眠宁和安定)在平息人类恐慌发作方面也几乎没有效果,尽管一些现代高效苯二氮卓类药物,如阿普唑仑,相当有效。另一方面,最初的三环类抗抑郁药丙咪嗪可以在低剂量下改善恐慌障碍。事实上,丙咪嗪是第一个被发现对人类具有明确抗恐慌效果的药物,也能减少动物的分离叫声(Klein & Rabkin,1981年;J. Scott,1974年)。
人们还可以在临床基础上区分恐慌/悲伤和恐惧,因为它们动员不同的自主神经反应。自主神经系统有两个主要分支。交感神经分支使动物为主动反应做好准备。例如,交感神经系统可能提高心率和呼吸频率,从而为燃烧升高的血糖水平提供氧气,这对于逃跑是必要的。它同样可能扩大瞳孔以提高警觉性。另一方面,副交感神经分支在动物处于更被动状态时接管。在副交感神经系统的影响下,心率减慢,呼吸规律,瞳孔保持未扩张状态。副交感神经系统对情绪变化也很敏感,它促进眼泪、唾液分泌和性唤起。
预期焦虑(条件性恐惧)的特征是普遍的忧虑紧张,倾向于产生源于自主神经系统交感神经臂的各种症状。因此,快速心跳、出汗、胃肠不适和肌肉紧张增加等症状是恐惧的特征。然而,恐慌/悲伤的表现伴随着虚弱感和抑郁性倦怠,具有更多副交感神经性质的自主神经症状,如强烈的哭泣冲动,通常伴随胸闷和喉咙有肿块的感觉。虽然恐惧促使人们逃离加剧焦虑的情境,但恐慌/悲伤会引发关于失去情感对象的想法,并促使人们寻求所爱之人的陪伴。
尽管存在不同的情绪系统,每个系统都有特定的情感和行为特征,但它们经常以复杂的方式相互作用。关于依恋障碍(attachment disorders)有大量的心理治疗文献,依恋障碍是恐慌/悲伤系统的表现。患有严重依恋障碍的儿童无法信任他人,会拒绝自己的依赖感,无法与他人产生共情。他们往往贫乏、贪婪且提出不当要求,在青春期和成年期经常转向药物,特别是鸦片类药物和酒精。
患有依恋障碍的人也经常遭受持续的恐惧,这源于童年时期被忽视或虐待的经历。在审查被监禁的青少年的病史时,人们经常看到这种复杂的情况。这些孩子长大后会变得极具攻击性,并且常常表现出反社会行为。同时,他们经常对自己感到绝望。显然,他们在几个基本的情绪系统上存在问题。他们情绪需求和局限性的复杂性使得他们难以康复。充分理解涉及这些行为的大脑情绪系统,对于开发治疗技术和有效药物来治疗这些不幸年轻人的持续恐惧和依恋障碍至关重要。
创伤后应激障碍(PTSD)是另一种涉及多个不同情绪系统的复杂状况。除了长期过度活跃的恐惧系统和惊恐/悲伤系统(PANIC/GRIEF)的表现外,PTSD是一种恐怖状态,通常伴随着愤怒,正如我们在开篇小故事中提到的,这可能是反复创伤(如士兵在战时经历的)的后果。PTSD的一个方面,与单纯的惊恐/悲伤或恐惧不同,在于PTSD可以通过抗癫痫药物如卡马西平来减轻,而这种药物在控制惊恐发作或预期性焦虑方面并不总是有效(Berlin, 2007)。这表明存在一种额外的癫痫类型过程,可以将几种负面情绪演化为全面的PTSD状态(Agrawal et al., 2006)。尽管PTSD尚未明确与焦虑类型的情绪系统(如恐惧系统或惊恐/悲伤系统)联系起来,但它似乎是大脑可能受到创伤的另一种方式,可能涉及多个情绪系统的参与,如恐惧系统、惊恐/悲伤系统以及愤怒系统(RAGE)。
事实上,生活的变迁无常,我们每个人都会受到各种情绪挑战的冲击,因此几乎不可能证明任何情绪障碍仅仅是由单一情绪系统引起的,更不用说单一的化学物质失衡了。大多数人会反映出几种情绪失衡,这解释了为什么”共病”概念在精神病学中如此普遍。这本质上意味着不止一种精神疾病综合征同时发生。以抑郁症为例,它通常伴随着过度的心理痛苦、焦虑、愤怒易怒,以及寻求和追求其他生活兴趣的冲动减少。事实上,“抑郁症”这个术语非常模糊,既暗示着普遍的不适,也暗示着疾病。更准确的描述需要涉及相关的情绪系统,以及它们的过度或不足唤醒如何导致临床症状,包括越来越多的可能性——表征许多类型疾病的炎症级联反应在抑郁个体中过度活跃(Dantzer et al., 2008)。我们怀疑,在未来的某个时候,科学精神病学家可能不太需要目前使用的诊断类别,因为我们开始根据对失衡的大脑情绪系统的更好描述以及对可能导致情感困扰的许多神经化学变化的理解来理解情绪问题。
我们才刚刚开始理解潜在神经解剖学和神经化学的巨大复杂性。未来能够与更具体的情感心理治疗干预良好配合的生物精神病学,可能会基于更容易与患者实际情绪体验联系起来的知识。这种情况没有像应该的那样快速发生的一个原因是,许多研究者仍然认为心理学是一门软科学,最好将精神病学诊断类别直接与大脑事实的变化联系起来,而不需要中间的情绪分析。大脑中独特情绪系统的存在可能会促进一种比目前存在的更全面的心理生物学方法(Panksepp, 2004, 2006a, 2009a, 2009b)。
直到上世纪中叶,可用于治疗恐惧的唯一药物是阿片类药物、酒精、巴比妥类药物和甲丙氨酯(meprobamate)(后者被称为Miltown,曾经非常流行,但由于服用者容易自杀,已完全退出治疗实践)。这些早期药物有许多缺点,最严重的是安全范围差,通常导致意外过量或自杀。
由于焦虑通常伴随着自主神经唤醒,包括心率和血压增加,一种有用的策略是使用能够减少被称为内源性儿茶酚胺的大脑和身体唤醒化学物质作用的药物——特别是肾上腺素和去甲肾上腺素。这些大脑化学物质属于一组称为生物胺(biogenic amines)的物质,它们激活交感神经系统,在强烈的情绪唤醒期间进入高速运转状态——这种”战斗或逃跑”反应混合了大脑的愤怒系统和恐惧系统机制。无论如何,阻断它们的活动会产生镇静效果。β受体阻滞剂(beta blockers)(抑制一种类型的去甲肾上腺素受体)有助于症状性控制焦虑,如心悸和出汗。事实上,像普萘洛尔(propranolol)这样长期获批的药物有时被用来抑制公开演讲或表演期间的焦虑。艺术表演者和公众演说者服用这种药物来最小化可能妨碍巅峰表现的”紧张”并不罕见。
氯氮䓬(chlordiazepoxide, CDP)的偶然发现彻底改变了焦虑症的治疗方式。CDP的疗效是在1960年研究的最后阶段被发现的,当时霍夫曼-罗氏实验室关于苯二氮䓬类药物(BZs)的研究项目收效甚微,即将终止。几乎是作为最后的尝试,研究人员发现其中一种苯二氮䓬分子CDP在驯服当地动物园的野生动物方面非常有效。CDP很快以利眠宁(Librium)的商品名上市,在控制多种焦虑症方面取得了巨大成功。它能在远低于致死剂量百分之一的剂量下减轻焦虑。不久,更多更强效的苯二氮䓬类药物如地西泮(安定,Valium)问世,随后还有更多同类药物。这些药物几十年来一直是畅销药品。
苯二氮䓬类药物治疗初期常见的轻度镇静作用往往会迅速消退,而抗焦虑效果在长期使用中得以维持。最初,这些药物偶尔使用时似乎不会产生明显的身体依赖。然而,焦虑患者长期服用越来越高剂量的苯二氮䓬类药物很快成为普遍做法。对这些药物产生依赖的患者,停药可能会出现类似震颤性谵妄(DTs)的症状,即常伴随酒精戒断的意识混乱、躁动、幻觉状态。由于这些原因,某些苯二氮䓬类药物在医学界失宠,但同时它们也成为帮助戒酒者的非常有用的治疗方法。
很长一段时间里,神经科学家和精神病学家并不知道苯二氮䓬类药物为何能有效治疗焦虑。直到1979年发现苯二氮䓬受体,这项研究才得以开展。通常当像苯二氮䓬类这样的外源药物对大脑产生作用时,人们期望能找到大脑自然分泌的类似内源性脑化学物质。例如,恐慌/悲伤(PANIC/GRIEF)系统可以通过给予阿片类药物来平息,而大脑会以内源性阿片肽的形式产生类似化学物质。神经科学家假设大脑会产生一种类似苯二氮䓬的内源性化学物质,与苯二氮䓬受体结合,产生镇静效果。但情况似乎并非如此简单。当研究人员给予苯二氮䓬受体拮抗剂时,他们预期焦虑会增加,但这并没有发生。苯二氮䓬拮抗剂对增强或减少焦虑都没有作用。它们在心理上基本上是中性的。
研究人员随后发现,苯二氮䓬类药物并不独立减轻焦虑。它们不是单独作用来平息恐惧(FEAR)系统,而是通过增强γ-氨基丁酸(gamma-aminobutyric acid, GABA)的作用——GABA是一种抑制神经元活动、降低其放电速率的神经递质。苯二氮䓬类药物在GABA的A型受体上有自己的结合位点。苯二氮䓬类药物与GABA结合,会减缓恐惧系统(以及其他各种情感系统)的活动。这种增强的GABA传递作用使人和动物保持苯二氮䓬类药物治疗特有的平静安详状态。研究人员随后发现,其他较早的抗焦虑药物,包括酒精和巴比妥类药物,也是通过促进大脑中GABA介导的抑制来平息焦虑。实际上,GABA受体可以设想为一把有多个钥匙孔的锁,不同的钥匙可以同时插入。每增加一把钥匙都会增强主钥匙GABA的作用。许多人在GABA受体处酒精与其他药物的复合抑制作用中找到了缓解,不幸的是,他们也因此付出了生命的代价。这种组合会降低许多身体功能。
苯二氮䓬受体集中分布在恐惧系统从中央杏仁核到导水管周围灰质(PAG)的轨迹上,甚至更远到尾侧脑桥网状核(nucleus reticularis pontis caudalis),恐惧在那里调节惊吓反射(M. Davis, 1992)。苯二氮䓬受体也存在于大脑皮层的许多区域,这可能是它们能有效减少令人不安的思维的原因。因此,苯二氮䓬类药物能有效降低所有层面的恐惧性焦虑的活动,从惊吓反应到令人痛苦的想法。
尽管神经科学家发现了苯二氮䓬类药物如何与GABA分泌协同工作,但寻找内源性苯二氮䓬分子的过程并不顺利。研究人员至今还无法明确识别出一种执行同样功能的脑化学物质,但过去和现在都有许多候选物质。此外,大多数研究人员认为,如果存在一种与苯二氮䓬受体结合的内源性化学物质,它可能不执行与苯二氮䓬类药物相同的作用——它不增强GABA的抑制作用。大多数研究人员认为,这种内源性化学物质在苯二氮䓬结合位点上充当反向激动剂(inverse agonist),作用于GABA受体的方式是降低而非增强GABA的抑制作用。降低GABA抑制的结果将包括恐惧系统活动增强,使动物更加焦虑。担任苯二氮䓬受体内源性反向激动剂这一角色的主要候选物质是地西泮结合抑制剂(diazepam binding inhibitor, DBI),这是一种神经肽,当它与GABA受体的苯二氮䓬结合位点结合时,似乎会促进焦虑。但尽管经过多年研究,仍然没有确凿证据表明DBI实际上是大脑中主要的焦虑生成递质(Möhler, 2011)。其他神经肽如促肾上腺皮质激素释放因子(corticotropin releasing factor, CRF)有更多证据表明它们是大脑中强大的焦虑和压力促进系统。
目前,各种神经肽是特定药理学控制焦虑亚型的有前途的靶点。当在脑内施用时,许多神经肽会唤起恐惧系统。例如,CRF引起激动性唤醒,同时减少各种积极动机行为:进食、性行为、梳理、玩耍等。动物也倾向于在先前接受CRF的环境中冻结,这表明这些环境包含许多动物已学会恐惧的情境刺激。相反,由足部电击引起的冻结会因CRF受体拮抗剂而减弱。然而,使用CRF拮抗剂治疗病理性恐惧可能不可行,因为通常CRF也是一种有用的激素,通过血流传播,使大脑和身体有效地应对压力和危险。例如,这类药物可能会削弱免疫防御,加重肠易激综合征等身体疾病。无论如何,目前CRF拮抗剂主要被作为抑郁症的潜在治疗手段,这是一种现有抗抑郁药无法统一控制的疾病。尽管CRF拮抗剂在临床上有效,但已观察到有问题的副作用,如肝毒性。
除了CRF和儿茶酚胺外,许多其他神经肽也能激活恐惧系统。神经肽α-MSH在许多鱼类和爬行动物中促进伪装型色素变化。当这些动物受到惊吓时,它们的皮肤往往会变黑。虽然这种肽在高等脊椎动物中不控制皮肤色素沉着,但在小鸡脑内施用这种肽可以引发强烈的冻结/躲藏模式。促肾上腺皮质激素(ACTH)来自产生α-MSH的同一基因的同一片段,具有类似的效果。注射ACTH可以引发剧烈的逃跑,以及大鼠和其他动物的冻结。一个特别深入研究的肽是胆囊收缩素(CCK),它可以引发来自恐惧和恐慌/悲伤系统的广泛焦虑症状。神经肽Y(NPY)似乎也能够平息恐惧系统,因为NPY拮抗剂可以在动物模型中引发焦虑。如果这些发现得到进一步研究的支持,可能会产生一类特别有用的药物。
大脑含有许多其他激活恐惧系统的化学物质。兴奋性神经递质谷氨酸是传递无条件恐惧信号的关键,例如大鼠对猫气味的先天情绪厌恶。谷氨酸也控制无条件恐惧反应。如果在恐惧回路集中的内侧脑干区域施用谷氨酸激动剂,动物开始表现出自发的逃跑(通常处于半蹲姿势),伴随着明显的心理痛苦。视觉导向的动物如鸟类表现出快速的头部扫描、持续的发声和凸出的眼睛,暗示着深刻的恐惧。这些发作可以被谷氨酸受体拮抗剂抑制。然而,由于谷氨酸受体在大脑中广泛分布,控制着学习和我们大部分的高级认知思维,直接的药理学操作不太可能产生有用的抗焦虑药物。尽管如此,通过谷氨酸受体上的甘氨酸受体”侧旋钮”进行较温和的刺激可能是一种非常安全和有用的焦虑和抑郁治疗方法。这类药物目前正在开发和临床测试中。
自1939年以来,已知颞叶的广泛损伤会产生显著的恐惧缺陷,称为Kluver-Bucy综合征。随后许多缺陷被定位到杏仁核,它位于颞极的中心(图1.1)。因此,人们普遍认为杏仁核位于恐惧系统的中枢。杏仁核由大约十几个核团或部分组成,其中几个被称为基底外侧杏仁核(BLA)复合体,参与恐惧条件反射。这一点通过以下事实得到了加强:在几乎每一项与焦虑或负面情绪(偶尔也包括积极情绪)有关的脑成像研究中都可以看到杏仁核的唤醒。正如我们将在下一章(专门讨论学习和记忆)中更详细讨论的那样,BLA充当将恐惧线索传递到杏仁核中央核的通道。中央核位于内在的、原始的恐惧系统的最顶端,但BLA中的核团不是该初级过程情绪系统的一部分。因此,尽管杏仁核中的BLA核团在经典条件反射中起着关键作用,但它们的重要作用似乎在于将信息传导到恐惧系统的能力,而不是自身产生恐惧的能力。因此,虽然杏仁核的中央核是无条件(本能)恐惧系统的一部分,但其他核团则不是。
那么杏仁核中央核是否就是恐惧系统的核心?鉴于恐惧系统还包含许多在杏仁核进化之前就已存在的更深层结构,中央核不太可能是该系统最重要的部分。事实上,患有Urbach-Wiethe病的人类患者,其大脑两侧的部分杏仁核(特别是基底外侧复合体)会逐渐完全退化,但他们仍然有丰富的内心焦虑和丰富的情感生活。尽管这些杏仁核受损的人通常被报道在识别脑成像实验中常用的静态恐惧面孔方面存在缺陷,但随着研究人员更仔细地研究恐惧缺陷,结果并不像早期研究所显示的那样明确(例如,Talmi等人,2010;Wiest等人,2006)。同样,对创伤后应激障碍(PTSD)患者进行的脑成像研究(这是一种习得性恐惧)通常发现杏仁核的激活强度高于正常水平,但有时并非如此(Lanius等人,2005)。这也表明恐惧可以源自杏仁核以外的大脑区域。还有一个事实是,通过手术剥夺了杏仁核内所有神经元的幼年动物,在保持穿过该区域的神经纤维通路完整的情况下,仍然能够表现出恐惧和焦虑气质,这可能是由恐惧系统中更深层的结构所产生的(Amaral等人,1992;Kalin等人,2001)。因此,在发育过程中,上脑许多区域的恐惧能力很可能是由下脑区域编程的。
关于高级脑区恐惧的编程,研究人员需要考虑,在生命早期,甚至杏仁核和相关的颞叶结构也可能都需要由恐惧系统中更深层的结构(如导水管周围灰质PAG和下丘脑)进行编程,然后才能让高级脑系统更好地评估恐惧刺激和情境。同样,成年动物的习得性焦虑可能严重依赖于下层结构的影响,例如杏仁核影响额叶和扣带皮层。然而,目前这些可能性在很大程度上还只是推测性的。
无论如何,与许多媒体报道(在研究恐惧学习的科学家的鼓励下)相反,杏仁核对于产生焦虑感并非绝对必需。相比之下,导水管周围灰质和下丘脑肯定是必需的。这是因为恐惧系统,就像愤怒系统(图4.1)一样,是分层组织的,其中源自中央杏仁核等高级情绪功能,完全依赖于下脑功能(例如下丘脑,其情绪功能依赖于完整的导水管周围灰质)。
如果恐惧可以由比杏仁核更深层的结构产生,如果杏仁核对于幼年时期本能性恐惧的产生并非必需,那么即使是杏仁核也可能因为被恐惧系统中的下层结构编程(教导)而获得其大部分恐惧产生能力。这种由下层脑结构对高级脑区(如杏仁核)的编程,在其他情绪系统中得到了越来越充分的证实,特别是寻求系统。我们将在下一章重点关注这个主要问题,因为它被恐惧条件反射研究者过度忽视了。这种自下而上的学习控制可能也适用于皮层中的情绪学习,尽管在这些问题上没有大量数据。无论如何,这里的论点是,情绪脑的深层部分教导皮层结构执行与情绪调节相关的各种认知策略。
这种编程在认知-知觉领域的一个著名例子是由诺贝尔奖获得者David Hubel和Torsten Wiesel(1979)阐明的,他们首次证明视觉皮层中的神经元是由视网膜编程的,以区分特定类型的视觉信息,如线条和边缘的方向,以及它们在特定方向上的运动。这些高度调谐的敏感性被认为构成了视觉的基本神经元语法,必须通过新皮层的发育编程和学习来实现。事实上,现在已知视觉皮层不是由基因内在编程的,而是由丘脑视觉通路向成为视觉皮层的高级脑区的典型投射所编程的。如果胎儿小鼠出生前该皮层区域被破坏,它们会在通常用于处理触觉的邻近新皮层区域发育出良好的视觉皮层(Sur & Rubinstein, 2005)。事实上,如果正常成年人一周不被允许看东西,并被教导阅读盲文,他们的视觉系统就会开始重新专门化以进行触觉的精细辨别(Elbert & Rockstroh, 2004)。这意味着许多皮层功能可以在一生中保持灵活性,当它们通常介导的功能不再需要时,可以适应其他技能过程。
因此,新皮层的所有区域往往通过早期条件反射获得其功能,这再次突显了教育的重要性,也表明高级脑区中情绪的三级过程参与主要是通过学习来发展的。人们可能会认为,在儿童身上越早建立良好的情绪习惯,他们的心智就会发展得越好,尽管这仍然是一个颇具争议的概念,因为关于这个主题的优质实证研究相当有限。然而,越来越多的研究表明脑心智(BrainMind)在整个生命周期中具有可塑性,这无疑给我们带来了越来越多的希望,即早期学习只在特定情况下才是最终定型的。在生活的许多方面,通过学习和适应过程实现健康成熟在任何年龄都是可行的,尤其是在儿童时期,他们在面对逆境时可以表现出惊人的韧性。
然而,某些大脑系统确实会迅速失去其早期充分承担我们通常期望在人类身上看到的功能的能力。同样,如果我们以视觉系统作为研究最充分的例子,视觉皮层的编程存在一个机会窗口期。这种编程必须在生命早期发生。如果视觉皮层在这个窗口期关闭之前没有被编程,视觉皮层将永远无法正常运作,动物将在余生中保持视觉受损,甚至失明。生物学上的机会窗口期在高级神经情绪过程的发展中也并不罕见。我们已经学会操纵其中一些过程。
例如,产后的群居动物(如绵羊)与幼崽建立联结有一个短暂的机会窗口期。如果母羊在分娩后2-4小时内无法接触到她的羔羊,她就会拒绝它。通常,在联结窗口期内,母亲会学习识别自己羔羊的气味,并将其挑选出来优先照顾,而不是照顾其他羔羊。然而,如果错过了联结机会,可以通过操纵大脑化学物质再次短暂打开这个窗口,要么通过直接施用母性神经化学物质(即向大脑输注催产素(oxytocin)),要么通过能够达到相同预期化学和情绪结果的物理和/或社会干预(这些主题将在[第8章]讨论照料系统(CARE system)时更详细地阐述)。在情绪学习、适应和成熟领域的一个重要问题,我们也将在后面专门讨论基本社会过程(性欲LUST、照料CARE、恐慌/悲伤PANIC/GRIEF和玩耍PLAY)的章节中探讨,即我们能在多大程度上发展促进人类大脑高级区域亲社会网络发展的社会结构。可以想象,这些积极的社会力量将能够非常显著地抵消恐惧(FEAR)的影响。
在现实生活中,我们经常看到初级情感过程和三级认知过程如何在表面上看似无缝的影响和反影响层面中融合。就恐惧(FEAR)而言,其表现可以是微妙而广泛的。要解读正在发生什么是非常困难的,尤其是当儿童试图整合引导他们发展的诸多情感力量时。
值得注意的是,在临床情境中,人们会遇到关于恐惧表达的难题,特别是当父母报告说他们的孩子似乎什么都不怕,并且不假思索地将自己置于危险境地时。有时父母认为他们的孩子真的无法体验恐惧;然而,对于大脑完整、具有完整的导水管周围灰质(PAG)、下丘脑和杏仁核回路的儿童来说,这是不太可能的。当前的大脑证据(其中一些在前一节中已介绍)表明,只有大脑恐惧系统低级核心的损伤才能使一个人真正”无所畏惧”。因此,看起来无所畏惧的儿童实际上可能在他们的低级脑区相当恐惧,但他们正在成熟的高级脑区尚未整合这些信息,而且新皮层可以对低级脑功能施加抑制性控制。在童年时期,心智的顶部和底部——即三级和初级脑心智过程——并非注定会很好地协同工作。
事实上,表面上无所畏惧的儿童往往专注于内心的焦虑以及避免这种负面感受的方法。一个这样的无所畏惧的6岁儿童,最近看了电影《大白鲨》(Jaws),他谈论鲨鱼可以重新长出牙齿,以及海星可以重新长出”手臂”。他大声笑着说人类也可以做到这一点,然后他拿起一把画笔,像挥剑一样向一个看不见的敌人挥舞。在这场想象中的决斗过程中,他有一两次说:“我不怕你!”似乎很可能他在大脑的某个层面实际上非常害怕,但他故意表现出的攻击性让他感觉好了一些。
当然,儿童有时会让自己置于危险境地,比如爬高梯或不看就跑过繁忙的街道。这通常是因为他们还没有真正理解那种特定的危险。然而,有些儿童故意让自己陷入危险,以此来吓唬和惩罚父母。在这些情况下,并不是恐惧系统(FEAR)不起作用。相反,是另一个情绪系统,也许是愤怒系统(RAGE),在发挥主导作用。与此相反,当遭受忽视的儿童让自己陷入危险以赢得他们无法通过其他方式获得的爱和关注时,可能是恐慌/悲伤系统(PANIC/GRIEF)在起作用。更糟糕的是,当儿童遭受虐待时,他们有时会从事危险活动,因为他们已经得出结论,认为自己很淘气,应该受到惩罚。从某种意义上说,他们正在用父母可能惩罚他们的方式惩罚自己。这些看似无畏的行为实际上是在努力整合并可能接受虐待父母的要求,从而赢得他们的爱。这也可能是恐慌/悲伤系统的一种扭曲表达。
在人类心理学中,愤怒系统(RAGE)和恐惧系统(FEAR)之间也存在密切关系。这两个系统紧密交织在一起,这解释了战斗和逃跑反应的紧密配合。这两个系统在解剖学和化学上相互交织但又各自独立,因此它们经常协同工作。一个系统相对于另一个系统的主导地位取决于环境中危险的类型。例如,如果有可能避免危险,恐惧可能占主导地位,动物会静止不动,希望被忽视。否则,如果危险太近、太迫在眉睫,动物会拼命逃跑。然而,如果捕食者不那么强大(可以成功攻击)或者无法逃脱,愤怒系统就会占据主导地位。然后,预期的猎物会攻击攻击者,希望造成伤害或制造转移注意力的机会,使其得以逃脱。
在临床环境中,这两个系统通常很难区分,特别是在与幼儿打交道时,因为幼儿看待世界和与世界互动的方式与成年人截然不同。当一个孩子发脾气时,她可能是非常愤怒。或者她可能是非常害怕。如果你回想一下遇到一个巨大而危险的捕食者的例子,你恐惧的尖叫和四处奔跑可能已经把它吓跑了。恐惧行为通常与愤怒行为并没有太大不同。当你愤怒时,你会大喊大叫、挥舞拳头,也许还会来回踱步。如果你感到恐惧,你肯定会表现出略有不同但同样充满活力的外显行为。成年人很少感到恐惧。但儿童的生活在情感上并不那么平静。通常很难判断一个孩子是非常生气还是非常害怕。只有当孩子平静下来,足以谈论他或她的感受时,才能发现真相,但这需要耐心和理解的沟通。
以一个4岁女孩的案例为例,她不同寻常地不愿意进入治疗室。一进入房间,她就开始四处扔玩具,无缘无故地向治疗师喊出抗议和辱骂的话语。她似乎在表达极大的愤怒,但原因不明。最终,她的治疗师能够从她那里套出一个解释。碰巧前一天晚上,女孩十几岁的哥哥做保姆,允许她玩一个恐怖的电子游戏,游戏中的反派戴着深色环绕式墨镜。然后,就在治疗会话之前,治疗师在停车场遇到了他的小病人和她的母亲。那天阳光明媚,治疗师戴着墨镜,碰巧与昨晚反派戴的墨镜相似。小女孩又害怕起来,当她的恐惧失控时,她发脾气了。她的脾气可能同时表达了愤怒和恐惧。她对她的治疗师很生气,因为他吓到了她,而他曾经承诺要帮助她。然而,她的主要情感是恐惧,这在表面上只是表现为愤怒。
他们说进攻是最好的防御。然而,愤怒的表达,当得到适度调节时,有时能产生积极效果,帮助儿童克服恐惧。一个2岁的小女孩曾被一只喧闹的小狗吓到,此后她害怕所有的狗,尤其是当她晚上试图入睡时听到狗叫声。她的父亲通过使用玩耍来抵消负面情绪,帮助她应对恐惧。一天晚上他和她坐在一起,当狗叫时,他轻蔑地挥挥手,说”小狗很愚蠢”,并大声补充道:“安静点,你们这些愚蠢的小狗!我们不喜欢你们!”然后他会心地笑了,告诉她小狗很傻,因为它们听不懂,还一直叫个不停。“我们在乎那些小狗吗?”他问道,摇着头。她也摇摇头。他说:“你知道我对小狗怎么想吗?”他的小女儿摇摇头。他用嘴发出”噗噗”的吐舌声,这让他女儿爆发出阵阵笑声。过了一会儿,小女孩加入了游戏,说小狗很愚蠢,它们应该”安静点”,她自己也发出了”噗噗”的吐舌声。在和爸爸玩了几天后,她开始在婴儿床上独自玩这个游戏。当她听到狗叫声时,她大喊:“安静点,愚蠢的小狗!”然后发出强烈的吐舌声。当她和父亲一起玩这个游戏时,看起来是一个轻松愉快的活动,引人发笑。但当她晚上独自大喊时,她似乎真的很生气。起初这是每晚的仪式,但几周后变成了间歇性的,最后消失了。通过这种方式,她用一种克制的愤怒表达克服了恐惧。最终,她对狗的恐惧消失了,她能够抚摸邻居的狗而没有任何痛苦的迹象。
当然,以这种方式使用神经科学数据完全依赖于临床直觉。除了通过准确解读儿童的本能表现,没有办法真正知道他们大脑中哪些系统是活跃的。这表明人们只能估计儿童初级加工情绪系统的运作。随着成年人,理解基本情绪系统变得越来越困难。
大量证据表明,大脑原始部分的回路产生恐惧状态——这些状态在我们更复杂的认知能力之前很久就已进化形成。尽管我们已经了解到大量关于恐惧条件反射(fear-conditioning)——即恐惧与世界事件的习得性关联——是如何产生的(见下一章),但对恐惧系统本身的研究在英美研究传统中相对被忽视,但在世界其他实验室中并非如此,特别是在巴西圣保罗的实验室(例如,Brandão等,2008)。因此,在我们对这个系统以及所有其他原始情绪系统有完整了解之前,还有大量详细的情感神经科学工作需要完成。
尽管如此,历代许多有思想的观察者都承认这种原始恐惧状态的存在。这就是为什么我们选择杰克·伦敦的《白牙》作为本章题词。这只年幼的狼从未”遇到过任何值得害怕的东西。然而恐惧就在它体内。它通过一千次生命从遥远的祖先那里传下来。这是它直接接收到的遗产……通过之前所有世代的狼传下来”(第52页)。这个虚构的描写对人类而言也包含了不少真实成分。
一旦我们更彻底地从科学角度理解这种”恐惧本身”,我们将能够逆转人类精神的许多内在和习得性困扰,从慢性焦虑障碍到创伤后应激障碍(PTSD)。因为我们共享这种祖先情绪,动物大脑研究最终可以帮助阐明我们自身焦虑的深层本质,以及我们如何在与世界的互动中体验恐惧。下一章将深入探讨大脑创建的情绪记忆的神经科学,从简单的皮层下学习开始,进而到更高级皮层的参与。有许多方法可以将恐惧系统塑造成它所能变成的动态灵活的恐怖。正如我们所见,也有许多方法可以驯服它,从对系统初级加工情感能量的药理学影响,到可以通过明智的亲社会和心理治疗干预最大化的三级加工认知调节。正如罗斯福总统如此深刻地指出的:“我们唯一需要恐惧的就是恐惧本身。”因此,当我们最终从科学角度理解恐惧系统时,我们将更准确地知道他在说什么。
我真诚地抱歉,人类的统治
打破了自然的社会联盟
并证实了那不良的观点
使你惊慌失措
看到我,你可怜的、地球所生的伙伴
和同样终有一死的生灵!
然而与我相比,你仍是幸运的!
只有当下触及你
但哦!我向后投去目光
看到阴郁的前景
而向前,虽然我看不见
我猜测并恐惧
——罗伯特·彭斯,《致老鼠》(1785)
在他的八句诗中(上面分享的是第二节和最后一节),苏格兰诗人罗伯特·彭斯强调了老鼠和人类之间恐惧的连续性。虽然老鼠通常在当下时刻体验恐惧,以应对明确的环境挑战,但我们在脑海中向前和向后看的能力可以创造出想象的幻影([图5.1])。通过我们的自传体记忆(autobiographical memories),我们人类以及可能一些其他动物具有在充满个人意义的情感丰富的回忆质地中进行主观时间旅行的能力。正如诺贝尔奖得主埃里克·坎德尔(Eric Kandel)所说,他研究了海蜗牛中恐惧类型学习(疼痛的经典条件反射)的神经学,“对我们所有人来说,外显记忆(explicit memory)使我们能够跨越空间和时间,召唤已经消失在过去的事件和情绪状态,但它们以某种方式继续存在于我们的脑海中”(2007年,第281页)。但记忆并不总是外显的。有些是内隐的(implicit),认知上无意识的,但仍然在情感上能够影响行为。
人类的许多情绪记忆确实在不知道其原因的情况下产生,但这并不意味着它们伴随的情感没有被体验到。事实上,尽管改变感受的认知原因可能通常是无意识的(也许可以通过精神分析来提取),但感受本身并非如此。由于情感是现象意识(phenomenal consciousness)的一种形式,体验到的感受不应该被认为是无意识的,尽管它们的原因可能在认知上是不可穿透的。这只是情感记忆具有重要精神病学意义的一个原因。我们想知道为什么我们会有这样的感受,而且通常最好在心理健康专业人士的帮助下确定来源。因为情感是我们生活中主要的心理”力量”,精神病学家可以确信,我们也可以确信,我们的高级心理装置在很大程度上是由我们的情感体验与世界的许多挑战和变迁相互作用而塑造的(Davidson et al., 2003)。
在本章中,我们将重点关注我们对情绪记忆——我们的次级加工情绪(secondary-process emotions)——如何形成的日益增长的理解。令我们惊讶的是,这个主要源于恐惧条件反射研究的大型研究领域,很少考虑大脑的初级加工恐惧回路(primary-process FEAR circuits)——大脑中产生原始情感的非条件性情绪反应系统——在产生恐惧记忆中的关键重要性。大多数研究者只是将这种初级加工情绪整合系统视为学习过程的单纯”输出”。这是行为科学家避免动物情绪体验给我们更全面理解哺乳动物,特别是人类的脑心(BrainMind)功能带来难题的一种方式。
在探讨情绪记忆的本质时,我们将主要关注动物恐惧学习研究中丰富且精心培育的传统。我们在这里谨慎地使用这个大写术语,因为情感变化在很大程度上没有被正在研究恐惧学习和记忆细节的当代科学家所涉及。然而,由于史蒂夫·马伦(Steve Maren)(见下文)强调的交流鸿沟,我们也将尝试填补那些对学习比情绪更感兴趣的科学家所忽视的各种主要知识空白。因此,当我们介绍初级加工情绪问题——该系统的非条件性反应——时,我们将使用”FEAR”(大写),而当我们描述恐惧条件反射者的工作时,我们将使用小写形式。
最常见的模型是大鼠和小鼠的经典恐惧条件反射,类似于埃里克·坎德尔在海蜗牛中使用的巴甫洛夫程序。在啮齿动物模型中,音调和光线(条件线索)紧接着是对动物脚部的电击。经过几次配对后,动物仅对音调和光线就表现出强烈的恐惧。如前所述,很少有以这种方式研究动物的人明确承认动物感受到疼痛和恐惧。有些人说,动物思维的这种主观方面无法通过经验研究(LeDoux, 1996)。但我们认为这是错误的,因为我们知道通过脑刺激直接唤起恐惧和其他原始情绪系统,可以在各种学习任务中作为惩罚(Panksepp, 1991)。这是得出某些类型的大脑活动实际上被动物体验到的结论的金标准。由于现有证据表明,情绪感受源于大脑内非条件性(本能的)厌恶产生的恐惧网络,我们也许应该明智地考虑这种情感状态的神经学如何促进学习。有充分的理由相信,唤起焦虑的记忆,对动物和人类都是厌恶的,具有向下访问恐惧回路的能力。这并不意味着动物在认知上沉思引起这种感受的事件——当然,与人类相比,实验室大鼠和小鼠对恐惧的三级加工(tertiary processing)是有限的。相反,正如本章开头的诗歌所强调的,人类的慢性创伤性恐惧思想可能源于脑心的内心动力学(intrapsychic dynamics),它不仅沉思于对记忆中过去的焦虑,还担心对预期未来的忧虑,这些往往源于敏化的恐惧回路,这些回路有自己的初级加工情感思维。
在恐惧(FEAR)的情况下,创伤事件的记忆可能会促使我们遭受慢性焦虑和紧张,通常伴随着强迫性反刍思维,其中大部分可能发生在我们额叶的内侧区域(Northoff等人,2010)。相比之下,以更快乐情绪为中心的记忆可能会促进持续的愉快情绪,这往往会导致积极想法、希望和愿望的涌现。对忠诚和充满爱的父母的记忆可以留下终生的积极印记——这是应对未来逆境风暴的宝贵心理资源。这种早期安全的情感纽带是终生的情感馈赠(见第9章)。
当然,记忆是预测和处理未来事件的有用工具,以过去的成功作为未来行动的指南针。由于原始情感(affects)是动物也能体验到的祖先记忆,我们可以用类似的方式理解它们的功能。通过预测生存问题,内在的情感状态为行为提供即时指导。这些感受通过学习与世界事件相连。如果是这样,我们必须得出结论,动物在记忆实验中体验到它们被重新激活的情绪的许多方面。因此,通过融合我们本能的情绪能力和与外部生活事件相关的记忆,我们开始以情感的方式体验世界的许多中性方面。考虑到学习和记忆的巨大复杂性,这种情况有很多种发生方式,其中一些尚未得到实证评估。在这里,我们不仅将关注该领域最可靠的学习研究发现,还将关注那些很少考虑动物情感本质的研究者所忽视的、由情感引导的学习的潜在路径,这些情感源自动物的无条件情绪反应回路。
我们在恐惧本能章节之后暂停一下,来反思学习/记忆研究者的卓越成就。通过研究简单生物如海蜗牛(Kandel等人的工作)取得的重大神经科学突破,现在正在大鼠研究中得到效仿(Davis等人,2010; LeDoux,2000; Fanselow & Poulos,2005; Maren & Quirk,2004)。目前揭示的许多一般原则很可能会推广到其他形式的情绪学习,尽管细节上会有大量变化。令人遗憾的是,目前大多数研究恐惧学习的人并没有明确承认他们的动物正在体验恐惧。为什么大多数这样的研究者如此犹豫不决,不愿意考虑实验动物有任何恐惧体验,甚至有一个连贯的初级过程恐惧(FEAR)系统?
在一篇关于条件性焦虑的优秀综述文章发表后(Maren & Quirk,2004),Panksepp通过电子邮件向他欣赏的密歇根大学的年轻同事Steve Maren提出了以下问题:“为什么恐惧学习研究者从不提及或承认在中央杏仁核和导水管周围灰质(PAG)之间运行的恐惧(FEAR)系统的存在?”他友好地给出了一个非常清晰的回答:
我认为该领域普遍认识到,厌恶刺激可以在PAG和杏仁核的层面上募集无条件恐惧反应(尽管在每个层面募集反应的非条件刺激(US)的具体特性可能不同)——所以从这个意义上说,这个系统并没有被忽视。尽管如此,恐惧条件反射领域由主要对记忆如何工作感兴趣的人(包括我自己)主导,他们使用恐惧条件反射作为模型系统,而不是主要关注情绪如何工作的人(我假设你会归属于这个阵营)。所以回应你的问题,我认为存在两种文化,一种围绕记忆和条件反射构建,另一种围绕动机和情绪系统,这些文化以某种不同的方式处理相同的问题。当然,这些方法不应该独立运作,更具整合性的工作似乎正在出现,特别是现在人类神经影像实验已经被纳入其中。
这清楚地解释了该领域的分歧,即动物的情绪被赋予了次要地位。这也解释了一些奇怪的说法,比如”不久之前,它(即杏仁核)还是大脑中一个默默无闻的区域,相对很少引起科学界的关注”(LeDoux, 2007, p. R868)。事实上,现代以情绪为焦点的大脑研究的根源可以追溯到大脑的这一区域,即所谓的”克吕弗-布西综合征(Klüver-Bucy syndrome)“(1939)的发现,该发现强调了野生动物在颞极损伤后变得温顺,这些损伤包括杏仁核、部分海马体和周围的颞叶皮层(见图1.1)。半个世纪前,人们早已清楚”主要局限于杏仁核的损伤会使野生动物变得温顺。这一结果已在猴子、家猫甚至非常野性的山猫身上观察到。例如,野猫凶猛到不用网和防护手套就无法处理,但在对杏仁核适当部位进行双侧切除后就可以安全地抚摸”(McCleary & Moore, 1965, p. 121)。此外,著名的研究表明”当猴子以这种方式变得温顺,然后被放回与正常笼友在一起时,群体内的社会关系会发生变化。实验动物在社会等级中降到较低水平”(同上, p. 121,描述Rosvold等人1954年的工作)。关于杏仁核作用的一部重要合集在40年前出版(Eleftheriou, 1972),此后不久就已清楚,杏仁核的中央核对恐惧反应本身至关重要。最近发生的变化是,随着神经科学方法的改进,分析现在聚焦于比20年前所能达到的更精细的回路层面。近期工作确实为那些对学习感兴趣的人提供了对恐惧条件反射细节更精确的理解,但代价是将真正的情绪——这是非常广泛的大脑现象——排除在他们的等式之外。这产生了对大脑在情绪唤起时所做之事的相当狭隘的观点。本章不仅基于对恐惧条件反射神经机制所收集的精确数据的认识,也基于对那种狭隘的、行为主义的、非情感性的恐惧条件反射观点可能造成的错误的认识。
恐惧条件反射研究者收集的事实令人印象深刻,但一个关键问题继续被忽视——大脑的初级过程神经情感过程可能是恐惧学习如何发生的关键中介。虽然这不是我们的主要研究领域,但我们愿意预测,大脑的情感性、无条件的恐惧基质在建立杏仁核的恐惧条件反射过程中发挥着关键作用。事实上,这可以推广到所有各种”基底神经节”脑区(如杏仁核、伏隔核、终纹床核等),大多数次级过程情绪学习都在这些区域发生。这可能如何发生将在稍后详细讨论。但首先,让我们考虑故事更令人愉快的一面。恐惧记忆可以通过巧妙利用巩固过程本身的”治疗性”手段来消除或覆盖。“巩固(consolidation)”是指将短暂的体验首先转化为短期记忆,并在体验重复几次后转化为长期记忆的复杂大脑过程的名称。然而,当痛苦的记忆被提取时,它们可以被”再处理”,然后以不那么令人困扰的方式”再巩固”。
不久之前,记忆研究者认为持久的情绪记忆是大脑中永久的实体。一旦形成,它们被认为是不可改变的。一个比喻是它们像山一样坚固。这种观点不再站得住脚。情绪记忆永远是可塑的,会受到未来事件的影响——通过一种称为再巩固(reconsolidation)的现象(Nader & Hardt, 2009)。这一认识对有效的心理治疗尤其重要。如果我们可以通过在不同的情感背景下提取情绪痛苦记忆来减轻其刺痛——可以说,在心灵之眼中以不同方式旋转它们——那么就有可能在治疗上利用这样一个简单的事实:积极情感可以抵消消极情感。通过理解旧的和痛苦的记忆并非像山一样不可改变,不使用药物的治疗性改变就成为可能(尽管某些药物可以加速这种改变)。正如我们将看到的,这已成为新兴的现代心理治疗科学的一个主要主题(见第12章)。
通过更好的技术,治疗师应该能够更有效地引导来访者远离痛苦生活经历的记忆,转向积极的心理框架。许多困扰记忆中的伤害性方面可以通过新的积极视角的半影区域进行再巩固(reconsolidation),这些视角不那么折磨人。事实上,也许有一天,不良的情感记忆可能会通过药物得到缓和,比任何现有药物都更具体、更有效。例如,这种未来疗法可能通过使用所谓的β受体阻滞剂(beta blockers)(如普萘洛尔(propranolol),一种去甲肾上腺素(NE)受体拮抗剂)来抑制大脑中促进焦虑的去甲肾上腺素影响,从而减少伤害性经历的巩固(McGaugh & Roozendaal, 2009)。如前一章所述,这种特定药物通常用于减少”表现焦虑”中常见的身体唤醒,这种焦虑可能会破坏人们最佳展示其技能或知识的能力。它也可能有效地用于再巩固。目前,还有另一种药物,d-环丝氨酸(d-cycloserine),它能温和地促进谷氨酸传递,可用于在心理治疗期间通过直接促进再巩固过程,以更积极的方式治疗性地再巩固那些令人困扰的厌恶记忆。这个想法已经获得专利(Amaral & Roesler, 2008)。
简而言之,现在人们普遍接受记忆可以通过治疗进行重塑。在未来,它们甚至可能被清除(Schiller, et al., 2010)。然而,关于大脑如何记忆和提取过去事件的大量知识,现在需要通过更好地理解情绪唤醒(即无条件刺激引发的无条件反应)如何在大脑中建立学习过程来补充。因此,让我们梳理一下恐惧学习/记忆研究中的一些概念问题。
所有基本情绪系统都在大脑中促进大量的学习和记忆,在本章中,我们将描述这一过程发生的一些方式——学习和记忆(次级过程)如何扩展和阐述我们先天的初级过程情绪能力。尽管我们主要关注恐惧学习,但我们认为这些知识的大部分也将适用于其他情绪系统。然而,除了寻求系统(SEEKING)之外,其他情绪网络的细节并没有像恐惧系统那样得到充分研究。由于我们特别关注这项工作的临床相关性,我们还将详述许多仍需澄清的模糊之处。重申一下,我们相信我们体验”恐惧本身”的无条件大脑机制极大地影响了恐惧的学习方式(Panksepp et al., 2011)。很少有恐惧条件反射研究人员明确考虑过这种可能性(即,更好地理解恐惧无条件反应(UCRs)对于真正理解大脑中恐惧学习如何发生至关重要)。对于感觉情感,也许大脑的无条件刺激(UCS)过程更重要,但在考虑情绪学习时,我们需要记住,情绪感受与大脑的情绪行动系统(恐惧无条件反应)密不可分。因此,足部电击的疼痛直接进入导水管周围灰质(PAG),并在那里帮助产生冻结和逃跑的无条件恐惧反应。
换句话说,尽管该领域的开创性研究通常侧重于传统的学习概念(例如,在经典条件反射和操作性条件反射中使用预测线索来让动物预期事件),但我们认为数据证明需要在这些方案中更明确地考虑初级过程情绪系统——无条件反应的性质。过去没有这样做的原因是,神经科学家将无条件反应视为大脑的单纯”输出”,而不是整合的情绪系统。因此,他们很少努力去理解大脑的内在情感冲动,而这些冲动是他们必须用来获得条件反射的。相反,他们似乎满足于相信学习可以被充分设想为简单的”观念”联结(学习中的经典观点)——即,你只需要理解条件反应如何从外部条件刺激(如预测性的灯光和音调等CS,以及厌恶性的无条件刺激如足部电击等UCS)的配对中产生。
但是,一旦我们将恐惧条件反射中的UCRs设想为整合的无条件情绪反应系统,它们产生本能的恐惧行为以及伴随的惩罚性负面感受,整体图景就会发生巨大变化。从我们的进化视角来看,这些基本的大脑情感机制必然在情绪学习的发生过程中起到”工具性”作用。换句话说,对环境事件的无条件情绪反应就是大脑内部感受到的”奖励”和”惩罚”。如果是这样,恐惧本身可能在恐惧条件反射中具有最重要的意义。相比之下,传统的行为主义学习观点在很大程度上将讨论限制在情感中性的”强化”过程上。通过关注这些想象的机制,人们可能会对一个明显的事实视而不见:奖励和惩罚是被体验到的,如果是这样,那么”情感”的神经表征对”强化”行为变化的强度有重大贡献(我们将其称为”情感神经科学模型”)。这是对更普遍接受的行为主义观点的根本背离。行为主义方法从第一性原则(即因为主观体验不算数)就排除了对脑心(BrainMind)的各种积极和消极情感过程如何促进学习的任何明确考虑。
在我们研究传统恐惧条件反射如何被有效用于研究学习和记忆的神经机制的神经科学细节之前,让我们先尝试澄清大量学习和记忆研究领域中的其他一些概念问题。这需要一些时间,但我们最终会回到上面提出的关于恐惧条件反射本质的新观点,即情感很重要的观点,以及情感不重要的传统恐惧条件反射观点。为了做好铺垫,让我们首先关注关于学习和记忆的三个常见误解,然后我们将进行对多种类型和复杂记忆过程的扩展总结,之后再回到情感神经科学模型。
首先,非科学家通常认为学习和记忆是有意识的过程。当然,当人类学习他们想要掌握的东西时,无论是在课堂上还是在更广阔的生活舞台上,都会发生有意识的学习。在学术环境中,学习和记忆显然涉及精心设计的努力。作为一个孩子,你必须通过反复努力有意识地应用自己,以便学习和记住乘法表和其他事实性记忆(统称为语义记忆(semantic memories))。事实上,这可能是迫使皮层学习通常是枯燥材料的唯一方法。据我们所知,大多数其他动物的学习发生在有强烈感受参与的时候。事实上,如果让我们自己做主,我们就像动物一样。当我们的兴趣——我们的寻求(SEEKING)——被唤起时,我们学得最好。所有其他情绪唤醒也促进它们自己的学习形式!
人和动物通常自动学习和记住最重要的事情。例如,我们那只害怕的老鼠别无选择,只能学会害怕我们那只象征性的猫脖子上戴着的铃铛。同样,大多数人记得肯尼迪总统被枪杀时他们在哪里,或者9/11暴行改变世界时他们在哪里。他们没有努力去学习或记住这些事实。他们情不自禁地这样做了。情绪”闪光灯”记忆会自动在我们的大脑网络中得到巩固,我们认为,这是因为伴随情感唤醒的神经化学物质的力量。事实上,对于人类来说,我们还必须考虑到,在强烈情绪事件的心理混乱期间,我们可能会学到在很大程度上是在我们想象中构建的东西。
其次,我们倾向于认为学习和记忆总是涉及在有意识的”觉察”中体验到的认知功能。我们认为,当我们学习时,我们有意识地弄清楚了一些东西,这就是让我们能够记住的原因。情况很少如此。例如,人和动物通过程序性学习(procedural learning)获得和保留身体技能,如骑自行车——这是一种只涉及练习而非认知洞察的记忆形式。尽管程序性学习通常涉及某种程度的指导,这确实涉及体验到的认知,但当我们”记住”如何执行一项技能时,我们不会反刍那些指导。主动意象可以促进和完善表现,但正是程序序列的反复执行使其成为我们日益完善的运动习惯装置的一部分。我们通常在不考虑它们的情况下完善新技能的表现。事实上,思考自己在做什么实际上会扰乱表现。人们甚至以这种方式养成情绪习惯,作为程序性学习的运动序列,这些序列表现在他们的语调、手势、姿势和整体情感人格中。
高级认知体验在导致情绪学习和记忆的大脑条件反射机制的成功中也不起本质作用。我们将讨论的大多数学习都是在没有新皮层帮助的情况下进行的。情绪学习涉及对先前中性体验获得情绪反应。情绪记忆是这种反应随时间的保持。引发习得性情绪变化的线索可能经常未被察觉——它们可能完全是无意识的。然而,我们认为这不适用于引发最显著情绪学习形式的非条件刺激——例如,将音调或光线与足部电击配对。即使是没有新皮层的动物,足部电击的疼痛和由此产生的恐惧肯定也会被体验到。这些动物表现出所有疼痛的指标,事实上更加强烈。这并不是说学习总是需要情感体验。它不需要,特别是在严格认知形式的”陈述性学习”(declarative learning)中,这种学习通常需要死记硬背(例如,7乘以7等于49)。但它通常涉及现实生活记忆(“情节性”记忆)。无论如何,对于我们在这里将考虑的情绪学习类型(有很多很多类型),新反应的获得是自动和非自愿的——所有必要的回路都位于新皮层下方。在这种情况下,重要的是要记住,各种初级过程情绪的原始体验也是皮层下产生的——它们是大脑非条件反应的各个方面。
第三,许多人认为只有一种类型的学习和记忆。这不是真的。科学家对这种复杂性进行了许多许多子概念化的方式(产生的想法并不总是相互独立的)。例如,我们上面提到的程序性记忆(procedural memories)和语义/陈述性事实记忆(semantic/declarative factual memories)(从中构建更个人的、情感丰富的、情节性的和自传体记忆)。这里不是讨论这些复杂性的地方。我们只想强调,最好的神经科学工作是用最简单的情绪记忆类型完成的,即可以通过使用巴甫洛夫经典条件反射原理(至少从外部)研究的事实记忆。如图6.1所示,这种学习的一种稍微复杂的类型是情境性恐惧条件反射(contextual fear-conditioning),动物学会害怕它们遇到捕食者气味的区域。我们通过这些研究学到的重要经验不一定适用于许多其他具有重大个人和临床意义的学习类型。
总之,尽管我们经常倾向于根据意识意图和高阶认知来定义学习和记忆,但学习和记忆的许多方面既不是有意识的,也不一定是认知的。这些记忆可能发生在我们拥有情节性-自传体记忆能力成熟之前很久——在我们能够回忆起生活中的各种事件之前,其中许多具有深刻的情感意义。这就是为什么早期童年创伤往往如此难以治疗的原因。尽管人们对成年生活中发生的许多事情感受强烈,但他们往往无法知道早年巩固的情感原因,那时他们还没有长期明确自传体记忆的能力。这些是用”谈话疗法”最难管理的情绪记忆。在本书倒数第二章中,Panksepp将讨论可能解决这些早期情绪记忆的替代疗法。
图6.1. 在四个基线玩耍日之后,猫的气味在单个测试日被引入玩耍室(即,在标准的5分钟观察期间)。尽管在所有随后的日子里房间都是干净的,但玩耍邀请(即背部接触)在3天内显著减少,而压制行为在所有5个随后的测试日中都有所减少。对照组(实线)没有暴露于任何猫毛。数据为平均值和±标准误差(数据来自Panksepp等人,1994年;改编自Panksepp,1998a,并经牛津大学出版社许可再版)。
在我们能够科学地处理这些微妙问题之前,还有大量的神经科学工作要做。到目前为止,大多数实验工作是通过经典条件反射研究非常简单的、最近获得的情绪-行为记忆来完成的,而不是来自许多过去生活经验的看似自由浮动的情感残留。尽管如此,不能过分强调的是,过去几年的一个重大发现是,每次记起某事时都会发生记忆的情绪再巩固(emotional reconsolidation)(Hardt等人,2010)。可以设计出精明的新临床干预措施来软化甚至被遗忘记忆的令人不安的情感影响。该领域充满了关于如何重新处理此类记忆的新想法(例如,参见Fosha等人,2009a和第12章),但该文献过于广泛和重要,无法在本章中充分涵盖。
学习和记忆远比表面看起来复杂,研究者已将该领域划分为许多不同的概念。在最粗略的层面上,研究者区分了外显(明确的认知体验)和内隐(非认知性的,但通常是情感性的体验)记忆。我们会偶尔使用这种分类。外显记忆有陈述性、情景性和自传性形式。最常见的内隐记忆形式是程序性记忆(procedural memory)(例如,学习一项新的运动技能)。在这种复杂性之上,有时也在其下方,我们有短期、长期和工作记忆的概念。因此,现代认知科学家和神经科学家以各种方式解析了记忆的复杂性。我们无法达成共识,认为这种划分是在”自然接点(natural joints)“处优雅地完成的,而这些接点才能恰当地构成学习的多维复杂性。因此,现在断言不同类型的学习和记忆是完全独立的还为时过早。事实上,所有这些学习和记忆类别在大脑内共享许多重叠的神经化学过程。例如,它们都依赖神经递质谷氨酸、GABA、乙酰胆碱和去甲肾上腺素,在大脑的不同区域实现各自指定的功能。
对我们所有人来说,最有意义的记忆是那些高度修饰的、以自我为中心的生活记忆——所谓的重要事件的情景记忆(episodic memories),以及我们在其中的位置,产生丰富的、高度个人化的自传性记忆(autobiographical memories)。情景记忆本质上是一种完全形成的、具有个人意义的记忆,它整合了事件的许多方面,包括具体发生了什么、在哪里和何时发生、以及谁是主要相关人员的信息。这些区别首先由恩德尔·图尔文(Endel Tulving)(2001)强调。他强调了情景记忆,特别是亲密的自传性记忆,如何让我们在经历过的过去中向前和向后时间旅行——使我们能够想象,特别是预期和思考未来的可能性(Suddendorf & Corballis, 1997)。为了思考和反复琢磨这些可能性的重要性,人们还必须利用有限容量的、通用的”工作记忆”(working memory),这对于外显思维过程至关重要。在这里,过去的记忆可以被检索到一个心理工作空间中,从而产生新的视角。
图尔文认为其他动物没有如此深刻的思考性情景记忆,因为它们没有对自身的外显意识。然而,对包括灌丛鸦在内的几种生物的现代研究表明,这些动物可以在当前的思考中使用过去的信息来实现未来的目标(Clayton & Russell, 2009)。例如,在被另一只鸟观察时,在特定地点藏匿食物的鸟类,当再次独处时会将食物藏到别处。它们是否因此具有自我意识?在[第12章]中,我们将论证所有哺乳动物确实拥有一个核心自我(SELF),可能是深层内隐的,或者本身只是最低限度意识的,但构成了一个大脑基质,使动物能够拥有初级过程情绪感受(primary-process emotional feelings),并最终拥有对过去事件的外显情感记忆(Northoff & Panksepp, 2008; Panksepp & Northoff, 2009)。然而,我们不会在此广泛关注其他动物中这些重要的高阶问题,因为它们极难研究。要获得关于其他生物情景能力的线索需要高超的实验技巧。它们没有语言来传达过去的经历和未来的愿望。所有信息都必须从它们的行为中推断。从它们的情绪行为中得出它们有情感感受的结论,要比推断它们在认知上还可能想些什么容易得多。
让我们暂停片刻讨论陈述性记忆(declarative memories),之所以这样称呼,是因为我们通常可以将这些记忆用语言表达——我们可以”陈述”它们。这些是我们能够有意识地回忆起的事实性记忆。在日常言语中,当我们说”我的记忆”时,通常指的就是这种类型的记忆。我们这种类型的大多数记忆没有强烈的情感色彩。例如,如果你的弗雷德叔叔(Uncle Fred)打电话约你下周四共进午餐,你会将这一信息保留为陈述性记忆。但你对此不会有特别强烈的感受。你可能还会记得后院一棵树充满活力的秋色、9乘以7等于63、你的汽车刹车需要维修、以及你周四有很多文书工作要做。所有这些回忆都是陈述性记忆。然而,某些此类记忆可以轻易唤起强烈的感受,即那些在情绪唤醒过程中牢固建立的记忆。
人们普遍认为许多动物具有陈述性记忆(declarative memories),尽管在实验室研究中它们被简化为最基本的要素,即最简单的感知事件。在本章中,我们最终也会关注经典条件反射(classical-conditioning),这是最简单的陈述性记忆类型(即学习一件事跟随另一件事——通常是一个中性事件后跟一个情感唤起事件)。然而,在人类中,陈述性记忆可能要复杂和精细得多,所以让我们从这里开始。考虑一个假想场景:当你的弗雷德叔叔打电话时,你可能正在沉思,电话铃声可能吓了你一跳。你可能随后会记得电话的”叮铃铃!“声有多么刺耳。当弗雷德邀请你吃午饭时,你不仅仅记住了他在电话里说的声音。你必须理解和思考他所说内容的含义和影响。你必须查看你的日程表。你还想知道他为什么这么热切地想见你,这让你思考各种可能的原因。你担心他可能健康状况不佳。这些记忆让我们能够思考复杂的情况。
思考某事是一种认知行为,它依赖于将许多过去学习的片段保持在神经科学家称为”工作记忆(working memory)“的大脑部分,其神经机制高度集中在我们庞大的背外侧额叶皮层区域(Goldman-Rakic, 1998)。当你思考可能的人际复杂性时,你思考过程的部分内容可能随后被保留为新的陈述性记忆,特别是如果它们在你的心智大脑(MindBrain)中唤起了强烈的情感。你可能记得弗雷德声音中紧张的语气,尽管你的日程很满,他似乎急切地想要面对面见面。你肯定会记得你已经明确表示只能在周四见面。谈话结束后,你计划在记忆中固定这样一个事实:你最终安排在某个餐厅12:30见面……但是知道你必须记住多少事情,以及你有时提取存储信息的能力有多么有限,你在日程表上记下来,以帮助记忆。所有记忆都有短期和长期成分。工作记忆同时使用这两个成分,以及情景性、自传性和语义性内容。因为它使用如此复杂的记忆系统阵列,能够以各种排列方式进行处理,工作记忆的概念非常接近”思维”的核心含义。
当你在一个容量有限的工作空间中思考这一切时,你的工作记忆制定了一个行动计划:你将在周四早上6:00起床,7:00到办公室。首先,你将处理文书工作。可能到9:00就能完成,然后你的同事就会上班,能够接听你的电话。你已经预约在弗雷德建议的餐厅路上的一个修车厂保养汽车。在等待期间,你将花时间放松一下,边喝咖啡边阅读《纽约时报》。然后你将见你的叔叔。
工作记忆中的项目可以保留更长时间,以便被编码为陈述性记忆。当陈述性记忆被创建后,它就可以被工作记忆提取(供将来使用)。这意味着当你试图思考某事时,你将能够访问你过去的想法。当你思考弗雷德叔叔的午餐邀请时,你记得他是一位退休的神经科学家,想象力非常丰富。因为他对意识感兴趣,你怀疑他可能想分享一些奇怪的新理论,也许是详细阐述他关于大脑中多巴胺系统的生长如何引导人类心智和文化进化的疯狂新想法。无论如何,他紧急的电话在你过度创造性的想象深处(这不过是记忆在起作用)激起了许多情感和认知可能性。但是直到周四见面,你才会知道到底是怎么回事。
我们现在的唯一观点是,你对各种奇妙可能性的考虑源于你流畅的推理能力(reasoning capacities),这需要大量的新皮层能力,也就是神经科学家所说的工作记忆。工作记忆从我们大幅扩展的额叶获得复杂性,特别是更晚扩展的外侧延伸部分(即背外侧额叶皮层),这些区域的进化晚于更内侧的以情感自我为中心的高级脑区。我们只是附带指出,这种处理较小记忆单元的能力可能导致新想法,其中许多可能最终是妄想的。考虑到这种记忆能力,特别是在大脑内侧自我参照区域,由古老的多巴胺-寻求(SEEKING)脑网络激活,我们可以看到记忆形成在服务于个人需求时如何变得扭曲。例如,寻求唤起的模式,结合关于事件之间假定因果关系的记忆自发形成,可能导致小鼠和人类的各种以自我为中心的妄想行为。相关事件转化为因果信念,当寻求系统的较低脑区([第3章])与相关认知事件结合时,让我们能够设想我们物种的许多重大突破性想法是如何产生的,以及个体人类的许多特异性精神病妄想是如何产生的。
此外,情感性感觉通常会引导我们选择自传体记忆进行检索和讨论。继续我们之前的例子,你知道你的叔叔有几个严重的医疗问题。假设当你在餐厅遇到弗雷德时,他紧张而严肃。你问是否有什么不对劲。他说有一件严重的事情要讨论。他刚满65岁,注册了医疗保险(Medicare),第一次开始思考自己的死亡和遗产。他首先说他对你隐瞒了一个秘密很长时间。他带着一丝羞涩和羞愧开始讲述他的故事,这种情绪逐渐转变为内疚。突然间,你注意到他脸上和身体上的每一个情感细微差别。多年前,当你的父亲,也就是他的兄弟,被派驻海外时,弗雷德和你的母亲发生了一段不幸的恋情,期间你被孕育出来。弗雷德当时也已婚,当他的兄弟,你所谓的父亲回家时,他们都认为最好让你被抚养长大,就好像你是你母亲婚姻的孩子一样。但现在他的兄弟已经去世几年了,弗雷德不想在不让你知道真相的情况下死去。他才是你的父亲,而你一直称之为”爸爸”的那个人其实是你的叔叔。突然间,你的世界,无论是在认知上还是情感上,都被彻底颠覆了。
在那一刻,你巩固了一个”闪光灯”记忆(flashbulb memory),因为你的情绪被如此深刻地激发了。在最初的震惊之后,你被这次对话深深打动(被一个比你自己能想象到的任何信息都更令人惊讶的信息)。即使弗雷德只在那一次提到它,你也永远不会忘记他是你的生物学父亲。这样的情节记忆将需要你对自己如何理解生活以及你是谁进行大规模的重新定向。像这些如此强烈的记忆只有在你出现阿尔茨海默病或额颞叶痴呆(皮克病)特征性的精神退化时才会消退。显然,强烈的情绪唤醒在这些具有持久性的陈述性-情节记忆中扮演着重要角色,当它们被整合到我们庞大的个人记忆自传体仓库中时,有可能重塑我们。伴随这些记忆的高级心智功能,广泛分布在大脑中并且对精神障碍很重要,现在可以通过功能性磁共振成像(fMRI)等脑成像技术部分地可视化(Naghavi & Nyberg, 2005; Ragland et al., 2007)。
大多数学习-记忆研究的进行都没有太多关注情绪问题。自上世纪中叶以来的核心假设是加拿大心理学家唐纳德·O·赫布(Donald O. Hebb)(1904-1985)在1949年提出的一个假设。他的核心思想被著名的口号”一起放电的神经元连接在一起”所概括。换句话说,当网络中的两个神经元级联放电时,它们之间就会形成持久的突触键(Hebb, 1949)。
所有神经通路,无论是稳定的还是短暂的,都是作为神经元的串联而创建的,这些神经元在分隔它们与相邻神经元的微小突触间隙中分泌化学神经递质。突触前神经元分泌神经递质,这些神经递质与突触后神经元表面膜上的受体结合。数十年的研究表明,对于大多数记忆来说,主要的递质(但不是唯一相关的)是谷氨酸(glutamate),主要唤醒NMDA型受体(还有另外两种主要类型)。大多数神经科学家设想,在记忆的创建过程中,神经元通过突触间隙向各种突触后神经元上的受体分泌谷氨酸,通过这种强化放电的复杂链条,出现了一种被称为长时程增强(LTP)的现象(Bliss & Lomo, 1973)。LTP是由细胞内分子事件的级联介导的(这里太复杂无法讨论),它们增加了构成这些记忆的神经通路的可唤醒性。每次相同的通路被相关事件唤醒时,它们变得更加敏化,从而变得更加容易唤醒。
关于长期记忆形成的神经生理学,许多内容已经通过对海马组织切片的体外方法揭示出来(Tronson & Taylor, 2007)。尽管如此,即使这些细节被破译,在LTP的精细分子机制和我们一直在讨论的真实生活记忆的本质之间仍然存在着一道无知的鸿沟。要理解我们持久的个人记忆,我们需要更清楚地了解基本的情绪和动机系统——生物体继承的主要生存进化工具——如何参与学习。换句话说,精细的分子细节需要通过大规模神经元网络方法来补充——用混沌理论(chaos theory)的术语来说,就是非线性动态网络视角——来理解脑心(BrainMind)中学习的真实心理学。如第3章所述,我们也可以设想简化的体外模型来研究寻求系统(SEEKING system)中的预期性神经变化。这将是在记忆形成的机械分析中迈出的超越LTP的重要一步。
我们的记忆是在复杂网络的重要帮助下形成的,这些网络代表着生物体的需求和情绪,而不是简单的刺激-反应神经链。在这种现代观点中,学习可能反映了各种刺激-反应网络如何嵌入到更大规模的网络中,这些网络代表着生物体的主要生物学和心理关注点。我们相信,这种对心智大脑功能的整体图景是理解对我们最有意义的学习和记忆类型的必要组成部分,例如我们在与感受密切相关的自传体记忆背景下预测重要事件的能力。
记忆的消退与记忆的形成同样重要。关于为什么某些记忆被保留而其他记忆被遗忘,我们几乎没有可靠的知识。目前大多数神经科学家认为,遗忘是大脑消除未使用信息的主动过程,可能是一种反向学习的方式。除了发现增强记忆的化学物质外,科学家还发现了其他实际上可以删除记忆的化学物质。例如,最近通过一种称为zeta抑制肽(ZIP)的分子实现了新皮层中特定长期记忆的快速化学删除,它帮助从最初巩固记忆的突触中分散谷氨酸受体(Shema et al., 2007)。
在这种背景下,重要的是要像念咒一样再次回忆起最近的发现:记忆存储是一个持续的动态过程。记忆不仅持续受到巩固这一动态过程的影响,而且还受到”再巩固”的影响(Tronson & Tayor, 2007; Schiller, et al., 2010)。这意味着当人类和其他动物使用他们的记忆时,记忆因此恢复到主动处理模式,它们可以被重塑,然后以不同于原始记忆的形式再巩固。这种重构的记忆通常包括关于原始记忆巩固时不存在的新情绪背景的信息。因此,当在新背景中提取时,旧记忆会暂时变得不稳定,并相应地被重新处理。尽管弗洛伊德对这种大脑机制一无所知,但他似乎已经充分意识到记忆过程以这种方式运作的事实,并且他发明了Nachträglichkeit(延后性)这个词来描述以心理时间性和建构为特征的心理过程(Eickhoff, 2006; Faimberg, 2007)。这基本上意味着记忆不仅可以从过去重构到未来,还可以从想象的未来重构到过去。
我们认为,在特别熟练的心理治疗过程中,最令人情绪困扰的记忆可以通过这种方式有效地改变。这可能是某些形式的心理治疗比其他形式更有效的原因之一。从最近的证据来看,对比使用各种心理治疗方法的许多研究,似乎围绕记忆过程如此核心地展开的精神动力-精神分析方法通常会产生最持久的益处(Shedler, 2010)。这可能是由于对过去进行了更充分的情感再处理,而不是那些只关注当前认知解释的不太雄心勃勃的治疗方法——这些问题显然不应被忽视。
关于工作记忆的最初表述纯粹是心理学构念,像大多数心理学构念一样,工作记忆的概念并没有非常清楚地告诉我们潜在的大脑功能。心理学家最初关注的事实是,我们在任何给定时间只能记住有限数量的项目(七个项目加减二)。但大量证据表明,工作记忆更广泛地说是一种高阶认知功能。例如,工作记忆容量与智商成正比(Conway et al., 2003)。工作记忆的基质广泛分散在大脑的认知区域,但如前所述,它们显然主要集中在新皮层的背外侧额叶。这些神经基质帮助产生广泛的认知功能阵列,从语言识别到视觉/空间信息处理,到注意力和整体认知协调以及更高的推理过程(Baddeley and Hitch, 1974)。记忆的世界充满了复杂性,困扰着科学家被迫用来概念化我们心智认知网络的简单方式。
例如,工作记忆包含如此丰富的认知活动,更准确地说可以称为”工作学习”。它包含大脑功能,这些功能处理和协调来自外部感官的信息以及来自记忆存储的知识,更不用说所有这些发生的情绪背景,以及在此过程中可以改变记忆存储本身。这种学习可以完全在内部进行,通常在相关情绪唤醒的规则下,这一事实对于心理治疗以及人们在解释同一事件时的日常误解都非常重要。
目前,工作记忆在任何清晰的神经科学术语中都理解得很不充分。新皮层是大脑的主要认知基质。它的互连非常广泛和复杂,并同时使用许多脑区;经过一生的学习,它就像一个庞大管弦乐队的指挥。然而,重要的是要强调,皮层处理受到许多皮层下状态控制过程的控制,例如那些控制前脑乙酰胆碱、多巴胺、去甲肾上腺素、食欲素和血清素水平的过程,它们与每一个认知行为中无处不在的谷氨酸和GABA神经元一起工作。如果没有这些皮层下的全局调节系统,认知皮层将会严重缺陷。事实上,有证据表明,多巴胺这一充满热情想法(无论是理性的还是妄想的)的伟大促进者,在人类大脑中比在大多数其他哺乳动物大脑中具有更大的作用范围。从解剖学上讲,多巴胺网络在人类大脑中延伸到知觉皮层的深处,比在大鼠和大多数其他动物中延伸得更远,在这些动物中,这些网络仅限于额叶区域。事实上,在人类进化(包括文化和生物学)中多巴胺能过程的放大,包括捕食性SEEKING冲动,可能与我们皮层思维帽内信息处理组织的大规模扩张一样,能够解释人类心智的智力复杂性和妄想悲剧(Previc, 2009)。因此,各种初级过程情绪系统与高级新皮层-认知区域的相互作用,这些区域肯定会产生高阶心理后果(其中许多是文化塑造的),永远无法通过研究过于简单的动物模型来完全理解。相反,如果没有对我们仍然与所有其他哺乳动物共享的古老心智层次的清晰认识,那些更高的复杂性也无法被理解——没有这个基础,高级心智将会崩溃。动物模型总是需要用来研究任何和所有基本神经心理学机制的细节,但不适用于人类擅长的三级过程心智能力。
事实上,也许是因为动物模型不足以理解人类认知,我们可能永远无法在精细的神经层面上理解人类心智。例如,我们甚至还不理解支持简单知觉重组和巩固成复杂概念的神经科学。心理学家发现,工作记忆中可用项目的数量变化不大,但每个项目的复杂性确实会变化。我们不理解初级过程情绪唤醒如何与此类三级认知-思维过程联系起来,而要找出答案所需的详细神经工作在人类中是伦理上不可能的。但正如我们将看到的,我们确实拥有关于情绪唤醒如何与次级过程联系起来的坚实科学,次级过程是更简单的学习形式,如经典条件反射,尤其是恐惧条件反射。
现在让我们回到行为神经科学家青睐的最简单的恐惧学习模型。传统的恐惧条件反射像时钟一样精确。音调和电击,音调和电击,仅仅几次,动物就会仅对音调做出恐惧行为反应。学习的速度和精确性解释了这种令人不快的压力方法对那些有兴趣理解学习和记忆的大脑基础的人的吸引力。在研究这项研究之前,让我们简要回顾一下感觉信息是如何处理的。几乎所有感觉在到达新皮层之前都必须经过丘脑;唯一的例外是嗅觉。恐惧条件反射中最常用的两种条件刺激是视觉和听觉,它们各自由丘脑中不同的核团处理。然而,足部电击的疼痛感已经在丘脑下方的导水管周围灰质(PAG)中被感知到了。
事实上,所有感觉信息首先在皮层下处理,在某个时刻,大部分这种皮层下处理的信息最终到达丘脑,丘脑不仅作为将外部感觉信息向上发送到皮层的主要中转站,在那里被转化为精细的知觉,而且还作为一个分类、混合和再处理站。然而,传入感觉信息的情感成分通常会分流到下丘脑区域和丘脑的网状区域,这些区域不会投射到皮层。这对于恐惧条件反射可能很重要。足部电击不像普通的躯体感觉认知型刺激,比如头上戴帽子的感觉;所诱发的疼痛和恐惧具有高度情感性(首先是疼痛,然后是恐惧)。假设足部电击的厌恶性非条件刺激方面需要通过丘脑的投射核到达皮层才能转化为疼痛和惊恐,这可能是一个大错误。但这似乎是一些恐惧条件反射研究者所假设的(LeDoux, 2007, Figure 4)。
丘脑从皮层接收的”返回”信息(总是通过基底神经节间接传递)与它最初直接从感官接收的信息一样多。丘脑的不同区域神经元细胞群(核团)处理不同类型的感觉信息。例如,外侧膝状体核(lateral geniculate nucleus, LGN)处理视觉(光波)刺激,而内侧膝状体核(medial geniculate nucleus, MGN)处理听觉(声波)信息,依此类推。值得注意的是,这两种类型的核团可能都是丘脑感觉处理层面相当晚近的进化新增部分,因为它们都位于丘脑的远外侧边缘。味觉、触觉、痛觉和运动觉等感觉则更集中位于丘脑更古老的中央区域。最古老的外部感觉——嗅觉,一种原始的味觉形式(品尝空气!)——甚至不需要经过丘脑就能到达皮层的某些古老区域(梨状皮层(pyriform cortex)),尽管大量嗅觉信息确实会进入丘脑最古老的部分之一,即背内侧核(dorsomedial nucleus)。该核团对情绪处理也非常重要,尤其是与依恋相关的社交情绪,包括分离痛苦(separation distress)(见第9章)。无论如何,当前关于恐惧经典条件反射的研究主要局限于由新成员LGN和MGN处理的听觉和视觉刺激。
当LGN从外界接收视觉信息时,它会向两个方向发送信息:LGN将信息向上发送到感觉(听觉)皮层,动物在那里有高层次的、有意识的详细视觉体验。然而,LGN也将信息向下发送到杏仁核,那里是初级过程恐惧系统和愤怒系统的最高层级所在。这些已被称为条件反射的”高路”和”低路”。
对大鼠的实验研究表明,恐惧预测线索的皮层下视觉处理可以直接激活外侧杏仁核(例如,Doron & LeDoux, 1999; Shi & Davis, 2001)。在人类受试者中也观察到类似的皮层下通路(Campeau & Davis, 1995; Linke et al., 1999)。其他形式的低路感觉处理,涉及声音、触觉、味觉和嗅觉,可能也能激发各种情绪反应,尽管这可能因物种而异。然而,总的来说,动物不必有生动的皮层视觉、听觉、嗅觉、味觉或触觉体验,就能对某些刺激产生非条件(本能)情绪反应或简单的习得(经典条件反射)反应。低路处理通过将皮层下处理的感觉信息发送到丘脑来激发条件情绪反应,丘脑进而将信息向下发送到杏仁核恐惧系统的源头,即中央核。几乎按定义来说,当这种情况发生时,动物会体验到原始情绪情感(raw emotional affect)。同样,从经验上讲,情感的存在通过以下事实得到证明:仅仅对恐惧系统的电刺激就可以在学习中充当惩罚。
恐惧条件反射的皮层路径——高路(LGN到听觉皮层,然后再向下到杏仁核)——因其”干净”而受到称赞,因为它提供了高水平的刺激分辨率。例如,听觉皮层可以区分枪声和响亮的摇滚音乐。然而,高路处理信息相对较慢。低路可以更快速地处理信息。低路”捷径”——直接从LGN到外侧杏仁核——被吹捧为”快速”但”粗糙”,因为它处理时间更短(估计为千分之十二秒),并且不能为动物提供任何精细的感知区分(LeDoux, 1996)。高路慢了一半。不应过分强调这一点;这可能仅仅反映了它是一条更长的路径,有更多突触。无论如何,假设低路处理可能无法区分两种惊吓声音——例如枪声和响亮的音乐——但这种刺激辨别问题尚未得到充分研究。不过,我们可以确定的是,低路(脑干到LGN直接到杏仁核)可以更快地进行条件反射,至少在大鼠中是这样(图6.2)。
重要的是要明确,并非所有情绪的低路处理都是通过杏仁核进行的。例如,产生惊吓反射的突发巨响严格在脑干层面完成(这比到杏仁核的”低路”快两倍),而且已经确定,焦虑——持续的恐惧——会使这条惊吓通路敏化(sensitize)。例如,预测电击的视觉线索会使惊吓反射更强烈。因此,恐惧会使原始保护性反射敏化。我们还认为,恐惧回路的激活主要通过来自导水管周围灰质(PAG)的向上影响使条件反射机制敏化。从某种意义上说,恐惧系统是恐惧学习的指挥者。这不是一个流行的或甚至被广泛讨论的观点,因为我们所说的恐惧网络通常被视为是心理上空洞的、仅仅是条件反射的行为和自主神经”输出”系统(Davis, 1992; LeDoux, 1996)。
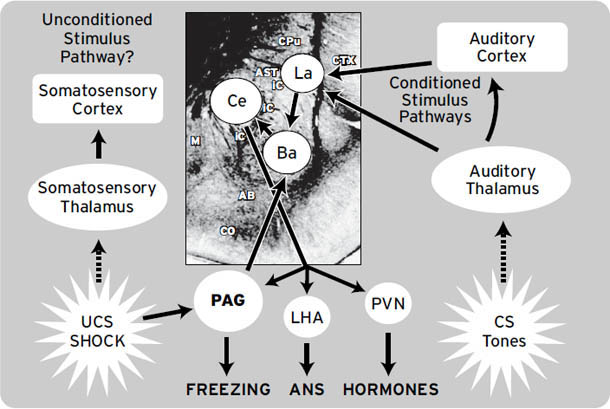
图 6.2. 恐惧反应经典条件反射的示意图概要。通常,在此类研究中,音调条件刺激(CS)后会跟随足部电击(UCS)。听觉刺激通过第八脑神经上行至脑干的耳蜗核,再投射到中脑听觉处理中继站下丘(未显示),然后投射到听觉丘脑的内侧膝状体核(MGN),最后投射到新皮层(右侧)。MGN有向下进入杏仁核的通路,首先到达外侧核(La),然后进一步向下到达基底核(Ba)。这些通路通常不会接入从中央核(Ce)开始的恐惧系统。然而,电击(UCS)在大脑中也有类似的上行通路,但它也会分流进入导水管周围灰质(PAG)的恐惧系统,后者直接激活UCR——无条件的本能恐惧反应。当CS进入杏仁核时,这里提出PAG的上行恐惧影响在导致”杏仁核门户开放”中起关键作用,从而在几次试验后发生学习之后,CS获得进入恐惧系统的通路。这是另一种情感神经科学的解释。传统观点认为,UCS像CS一样到达丘脑,声音和触觉在La中的结合是条件反射的关键联系。这两种观点仍有待直接的实证对比(改编自LeDoux, 2007,并有重大修改)。
目前有数十个实验室正在深入研究这个非常可靠的恐惧条件反射模型,使用足部电击。人们只能希望某些实验室最终会转向比足部电击更温和的恐惧”惩罚”,例如向颈后吹气,这会引起22-kHz超声波抱怨(Brudzynski & Holland, 2005),这种应激源产生的焦虑形式比即将到来的足部电击所产生的彻底恐惧要温和得多,后者会抑制这些相同的叫声(见Soltysik & Jelen, 2005)。对大鼠来说,一种不那么明显厌恶的”惩罚”是让它们暴露在猫的气味中;大鼠对这类我们几乎闻不到的刺激非常恐惧,它们会将这些经历与发生的环境背景联系起来([图 6.1])。这类更温和的模型可能与理解人类焦虑更为相关。事实上,考虑到积极情绪可以像恐惧系统引发焦虑一样快速地被条件反射,即如条件性挠痒反应产生快乐的50-kHz叫声(Panksepp & Burgdorf, 1999),人们只能希望那些单纯对条件学习机制感兴趣的研究者能转向反映积极情绪”奖励”学习的模型系统,而不是强烈的负面情感”惩罚”条件反射。Jim Olds在其生命末期进行的那种食欲条件反射(见[第3章])是一个不涉及任何厌恶刺激的快速条件反射的优秀模型。
无论如何,高路和低路的恐惧学习通常同时发生并相互补充;一般来说,大脑的高级区域调节低级区域,而低级区域唤起并敏化高级区域。因此,这些学习形式以目前未知的方式协调运作。在这种背景下,重要的是要记住,新皮层通过其许多向下的抑制性影响,可以平息情绪唤起。这当然是一种非常适应性的反应。假设一辆超速行驶的汽车在经过你身边时发生了回火。这个声音会首先通过大脑中最低的听觉”道路”进行处理,使你惊跳,这可能随后导致恐惧性唤起。你可能会僵在原地,睁大眼睛,呼吸变浅。片刻之后,各种新皮层认知会告诉你这只是一辆调试不良的汽车发出的爆炸声。你的新皮层随后会调节(抑制)恐惧系统的唤起,你会迅速平静下来。
正如我们所指出的,目前许多研究者将情感体验设想为认知活动的一个子集,从它们在完整大脑中确实强烈互动的角度来看这是合理的(例如,Pessoa, 2008)。然而,从进化的角度来看,将这些活动视为不同的控制层级是明智的:在初级过程层面(即使加上简单学习),没有理由相信几乎没有皮层的动物会对其经历的情绪唤起产生想法。尽管如此,我们不能排除这种可能性,特别是因为学会快速恐惧的啮齿动物确实需要高级内侧额叶皮层的参与才能消除恐惧。现在普遍认为,这个被称为”消退”(当奖励或惩罚终止时反应的停止)的过程不仅仅是遗忘,而是一个主动的学习过程。而这种消除学习需要比初始学习本身更多的脑力——即更多的新皮层参与(Myers & Davis, 2007; Sierra-Mercado et al., 2011)。
与所有情绪系统一样,还有很多需要学习的内容尚未被理解。一个巨大的谜团是,为什么直接刺激FEAR系统所产生的惩罚,不能像足部电击那样容易地与学习特定活动(比如响应预测性条件刺激按压杠杆以避免厌恶事件)建立联系(完整讨论请参见Panksepp, Sacks等人,1991)。相比之下,动物确实很容易对恐惧性脑刺激产生条件性场所回避(conditioned place avoidance)(Panksepp, 1998a, p. 214)。我们推测,这仅仅是因为动物很难学会如何害怕恐惧本身,如果没有疼痛的情感伴随。
但即使是通过电击介导的恐惧学习,目前也不完全清楚所有预测性刺激(听觉、嗅觉、触觉等)是否都能产生恐惧调节研究中常见的光信号或声音信号恐惧条件反射,或者有多少感觉系统能够快速、低级通路地接入FEAR系统。视觉可能不行。例如,在大鼠中,沿低级通路处理的声波会传到丘脑内侧膝状体核(MGN),比传到相邻外侧膝状体核(LGN)的视觉刺激更好地建立恐惧条件反射。然而,如果在生命早期通过手术诱导视觉系统终止于MGN,这些动物对视觉刺激的条件反射会更有效(Newton等人,2004)。显然在大鼠中,重要的不是哪个感觉系统进入丘脑,而是丘脑投射核向FEAR系统发送信息的效率如何。这表明不同物种中的不同感觉系统可能在介导快速恐惧条件反射方面有不同的”预备性”。
然而,条件反射的神经机制似乎不太可能在不同物种中有所不同,即从MGN到外侧杏仁核的传入感觉信息如何发展出通往杏仁核中央核(FEAR系统的源头)的新通路(习得性联结)。同样,从杏仁核中央核下行到导水管周围灰质(PAG)([第5章])的底层FEAR系统的运作原理,在不同物种中也不太可能有太大差异。一旦你对进化记忆(如无条件恐惧反应系统)有了良好的解决方案,以及对情绪记忆(恐惧条件反射)有了坚实的机制,为什么要抛弃它们呢?另一方面,人们当然会预期兔子比狮子拥有相对更大、反应更灵敏的FEAR系统,而且不同物种的高级脑机制会以不同方式处理情绪情境。
大部分最有价值的恐惧条件反射研究是在约瑟夫·勒杜的实验室完成的。他也是研究杏仁核条件反射的实际分子细节的先驱,效仿埃里克·坎德尔早期在海蜗牛上的类似工作。勒杜想要探究介导杏仁核亚区成功条件反射的神经变化。他的团队以及其他团队的条件反射研究非常细致,促进了许多额外的进展(Davis等人,2010;Ehrlich等人,2009)。但遗憾的是,这也导致了一个错误的观念,即杏仁核实际上是情绪产生的”总部”——情绪的中央车站(LeDoux, 1996)。事实并非如此。大脑中有许多区域对各种情绪的初级加工同样重要甚至更重要。如果大脑的任何区域值得被称为情绪的中央车站,那么显然应该是PAG而不是杏仁核。PAG以绝对关键的方式参与每一种初级过程情绪。相比之下,杏仁核主要参与FEAR(恐惧)、RAGE(愤怒)和LUST(性欲),而且它在刺激-刺激学习(CS-UCS)方面的贡献远大于对无条件(本能)恐惧反应本身的实际协调,而后者对这类学习更为全局性地重要。在这里,我们将只介绍勒杜关于恐惧条件反射的开创性研究所揭示的一些一般原则,这些研究得到了其他杰出研究者的充分补充,如迈克尔·戴维斯(Davis等人,2010)、迈克·范斯洛(Fanselow & Poulos, 2005)和史蒂夫·马伦(2005)。
很早以前就已经清楚,在条件反射建立之前,适度的声音和光线并不能直接激活恐惧系统(它们不像猫的气味那样是恐惧的非条件刺激UCS)。这就是为什么大鼠最初并不害怕经典条件反射实验中通常用作条件刺激的音调和光线。然而,关于最常用的非条件刺激——用来模拟捕食者咬伤的痛苦电击——的信息总是能够进入恐惧系统。疼痛直接影响恐惧系统的确切途径尚未被清楚地描绘出来。这可能是因为疼痛可以在多个层面进入恐惧系统——从导水管周围灰质(PAG)的底层(从而在大脑的非常低的层面激活整个恐惧系统),以及通过介导疼痛传递的各种丘脑网状核的高层输入。还有许多其他输入和促进因素,例如,促进认知联系的非特异性脑调节剂的通路,如乙酰胆碱和去甲肾上腺素。当关于电击的信息到达恐惧系统时,它可以与从丘脑内侧膝状核(MGN)到杏仁核外侧区域的低级通路传入的相关中性信息协同作用。当任何刺激被证明具有恐惧意义——持续唤起负性情感,就像电击总是那样——那么基底外侧杏仁核(BLA)中的神经细胞会通过发展与杏仁核中央核更强健的功能连接来做出反应。现在,中性刺激可以激活通往恐惧感受的”康庄大道”。
杏仁核中央核与下丘脑和中脑的许多低级脑结构一起构成恐惧系统(见[第5章])。当位于初级过程恐惧系统顶峰的中央核被条件性恐惧刺激唤起时,通常整个恐惧系统都被唤起了,大鼠会表现出全部的恐惧反应——僵住、血压升高、排便以及大量其他自主神经反应。这些不同的恐惧反应在脑干深处都有稍微不同的通路,但在完整的动物中它们通常都协同工作。动物还会表现出明显的负性情感——它们似乎以一种非常颤抖、害怕的方式感到紧张不安。重要的是要记住,足部电击产生的疼痛在神经系统中建立了这样的条件,即密切相关的刺激——预测性线索——可以控制那些预期性的条件情绪行为,这些行为本质上与非条件性(本能的)恐惧反应相同。换句话说,杏仁核中央核内恐惧系统的唤起(以及恐惧情感,这是一种神经状态)可能对学习的发生至关重要。
无论如何,正是在外侧杏仁核和中央杏仁核之间那些新的功能连接中,联想线索介导恐惧条件反射,因此这就是对恐惧学习感兴趣的行为神经科学家投入最多关注的地方,却没有清楚地承认恐惧唤起的情感性非条件反应对条件反射的发生至关重要!在低级脑区如导水管周围灰质中是否发生任何类似的条件反射仍不清楚,但如果发生了,我们预期它将是一个非常广泛的网络变化,正如”敏感化(sensitization)“所包含的那样——由重复的情绪唤起引起的情绪网络反应性的持久变化。
由于上述提到的条件反射发生所需的新的非条件反应-恐惧维度被忽视了,让我们重申条件反射的传统观点。当一只天真的大鼠最初暴露于非条件性预测线索(例如音调)时,它的恐惧不会被唤起。那么,由中性刺激和电击的快速连续引起的条件反射过程,如何赋予先前中性的线索新的进入恐惧系统的通路呢?勒杜(LeDoux)通过手术破坏了听觉皮层(高级通路)或从丘脑更直接通向外侧杏仁核的通路(低级通路),他发现高级通路受损的大鼠条件反射建立得相当快,这意味着预测性信息可以沿着低级通路有效地进行,并唤起大鼠的恐惧系统。另一方面,低级通路受损(即MGN损伤)的大鼠也建立了条件反射,但这个过程进行得非常缓慢。
MGN损伤的大鼠逐渐对与电击相关的音调产生恐惧反应。这种通过皮层的缓慢情绪条件反射在很久以前就已经被观察到了,对于预测食物的音调是如此,而皮层下区域神经元的条件反射进行得更快(Olds等,1972)。快速条件反射的优势是显而易见的,但皮层下系统为什么会比皮层系统条件反射更快却一点也不明显。
从我们的角度来看,很明显更古老的大脑系统应该在学习简单适应性反应方面具有优先权。换句话说,大脑的低级区域可能条件反射更快,因为它们在进化时间上更早获得了这种能力,因此它们在整个学习过程中具有某种优先权。如果认识到新皮层区域无法自行启动任何情绪反应,除非它们被训练这样做,这也许也是有道理的。只有经过持续的训练,某些高级脑区才能开始激发情绪。换句话说,新皮层只是缓慢地发展出向杏仁核传递明确认知信息的能力,以便激发习得的情绪性。这可能是严格的认知心理治疗方法可能不如那些同时使用熟练的初级过程情感操作的方法有效的原因之一(见[第12章])。
显然,指向情绪系统的皮层下感觉低级通路掌握着有效、快速条件反射的一些关键。但我们认为,实际上是本能的FEAR系统——震惊诱发的UCR(非条件反射)——为恐惧条件反射的发生提供了关键钥匙。然而,这并不是大多数传统学习理论家的看法。如果我们的观点被普遍接受,这种理解上的转变不仅会对所进行的实验类型产生深远影响,而且会对情绪障碍的临床治疗产生深远影响(见第12章)。无论如何,这种低级通路条件反射可能在没有意识”觉察”的情况下进行——没有任何认知理解——但它肯定充满了强烈的情感体验,即情感现象意识。
这具有重要的临床意义。认为感受只有通过新皮质中的认知-概念”库”以某种”读出”方式传递才能被体验——变得有意识——的想法,是一种信念,而非事实。相比之下,FEAR系统产生可怕体验的能力是一个事实,而非信念。无论如何,LeDoux的研究小组观察到,当大鼠被条件反射训练产生恐惧时,BLA(基底外侧杏仁核)中的细胞在发展与FEAR系统中央核的连接时会更频繁地放电。关于这种学习回路的总结,见图6.2,我们在LeDoux的工作基础上进行了修改,以突出FEAR系统可能从下方敏化杏仁核学习机制的方式(出于当前目的,我们将BLA区域内的独立神经处理合并为单一的BLA组件)。
BLA细胞利用谷氨酸,并作用于突触后谷氨酸敏感的NMDA受体(谷氨酸受体有几种类型)——据推测是将”沉默突触”转变为活跃突触(Kerchner & Nicoll, 2008)。这种联系——最初关闭的突触门的通透性增加,部分由局部抑制性神经回路介导(Ehrlich et al., 2009)——然后将来自BLA区域的条件反射信息转移到FEAR系统的中央核。这种恐惧条件反射机制类似于其他情绪系统使用的学习机制,这些机制正在以非常相似的方式被研究,但我们通常对其了解较少(例如,伏隔核中习得的食欲-SEEKING行为,通常将上行多巴胺系统的”奖励”力量纳入其条件反射视野)。到目前为止,似乎有一个普遍原则,即认知与原始过程情绪系统之间的这些接口都使用增强的谷氨酸能传递作为产生习得反应的机制。
尽管如此,情绪学习的发生似乎不言而喻,人和动物必须被非条件刺激唤起情绪激活。这表明非条件反应需要被视为条件反射过程中的一个积极部分——帮助解释如何与预测性条件刺激建立联系。如果是这样,那么神经系统的非条件情绪反应——在本例中是FEAR系统的唤起——对学习可能特别关键。在我们看来,FEAR系统介导恐惧学习的方式仍然是大脑中情绪学习如何运作的一个讨论不足的方面。这种对学习中UCR(非条件反应)的忽视可能反映出运动系统通常被设想为仅仅是”输出”回路,而没有认识到复杂的原始过程情绪行动系统实际上是脑内整合行动回路,具有心理-情感维度。
让我们提出一个具体的假设,这可能超出了一般读者愿意关注的范围:首先应该注意的是,谷氨酸受体分为两大类——AMPA和NMDA类型。到目前为止,大部分关注都集中在NMDA受体在介导条件反应中的作用上。我们认为,FEAR反应本身可能建立了必要的成分(也许是杏仁核中央核FEAR系统神经元中AMPA敏化谷氨酸受体的萌发),这可能是通过BLA复合体到达的条件刺激的许多”沉默”NMDA谷氨酸突触输入(Kerchner & Nicoll, 2008)随着学习的进行而渗透到中央核FEAR系统源头的必要条件。这是一个非常可测试的假设,值得更多的实验关注(但见Rumpel et al., 2005)。如果沿着这些思路的某些观点最终得到证实,它将再次突出大脑古老区域更古老的原始过程情绪网络在高级大脑功能如何运作中的首要地位。这是进化运作的最”合理”方式。
目前,关于最简单的恐惧条件反射模型的研究非常流行,我们预计上述假设很快就会被某人验证(也许在本书出版之前!)。我们预计这将由那些对中央核(central nucleus)亚功能非常感兴趣的人完成,例如最近关于中央杏仁核不同功能神经元群体的研究,其中外侧部分是条件反射所必需的,而实际的条件反射反应则由内侧分区的神经元驱动(Ciocchi et al., 2010),并且具有丰富的独特神经化学控制(Haubensak et al., 2010)。从直接干预的角度来看,也许最有趣的发现是,神经元的亚群受催产素(oxytocin)和加压素(vasopressin)控制,催产素通常会降低恐惧感,而加压素会增加恐惧感。由于女性大脑中的催产素神经元比男性多,而男性大脑中的加压素神经元更多(Panksepp, 1998a),这似乎表明在恐惧系统的源头就存在女性-男性在恐惧感方面的差异(Huber et al., 2005; Viviani & Stoop, 2008)。事实上,催产素增强了传统苯二氮卓类抗焦虑药物(如地西泮)的效力(Viviani et al., 2010)。这表明鼻内催产素最终可能在与焦虑相关的心理治疗干预中发挥作用,以及其他情感障碍如产后抑郁症。
总之,让我们关注一些关键的概念性问题(如 Maren & Quirk, 2004 所概述)。在基底外侧杏仁核(BLA)中看到的可塑性是真正的联想性的,因为简单的敏感化——重复电击——不会提高该核团中的神经元放电。学习仅在存在与大脑无条件反应配对的相关预测性刺激(例如,音调)时才会发生。基底外侧杏仁核中的学习可塑性完全不依赖于声音的皮质处理。条件反射在没有听觉新皮质的情况下完全进行。它也在没有内侧膝状体核(MGN)中任何先前的学习性变化的情况下进行,这些变化可能会将条件反射声音信息向下发送到恐惧系统。据我们所知,条件反射首先发生在杏仁核本身。然而,最终发生在丘脑(内侧膝状体核最终也表现出条件反射反应)的可塑性似乎确实依赖于已经发生在杏仁核中的条件反射。基底外侧杏仁核似乎”指导”它接收恐惧信息的丘脑内侧膝状体核区域。我们怀疑对于恐惧系统本身也可以这样说:以某种目前未知的方式,恐惧系统的进化记忆指导基底外侧杏仁核-中央杏仁核的连接进行条件反射。
最后,让我们简要考虑一下,当基底外侧杏仁核中的神经元被条件化时,条件性恐惧行为是否是绝对必要的后果?令人惊讶的是,事实并非如此。在训练有素的动物中,基底外侧杏仁核中的神经条件反射可以与恐惧行为分离。换句话说,人们可以测量基底外侧杏仁核内的条件性神经反应,而动物不表现出任何恐惧。这可以通过将NMDA受体阻断剂放入突触中来实现,在那里基底外侧杏仁核的信息可以被阻止进入中央核(本能恐惧系统的最上层结构)。因此,条件性神经元反应在基底外侧杏仁核中仍然明显,但动物在呈现先前的恐惧刺激时在行为上(因此我们假设在情感上)保持放松(Maren & Quirk, 2004)。我们的解释是,动物要体验学习性恐惧,条件反射信息必须首先进入恐惧系统本身。并且有许多更高级的过程可以进入这些初级过程(即,通往情绪的”皇家大道”——初级过程情感系统)。例如,除了对离散的条件刺激(如预测电击的音调)做出反应外,恐惧条件反射动物还会对许多复杂的情境刺激做出反应,这些刺激涉及它们受到伤害或只是在没有受伤的情况下受到惊吓的一般环境(例如,见图6.1)。
我们不仅害怕世界上引起恐惧的事物,我们也害怕危险的地方。因此,在现实生活中,以及在大多数实验条件反射情境中,恐惧相关刺激进入恐惧系统有几种并发的方式。通往条件性恐惧的另一条途径是通过海马体(hippocampus),它处理情境刺激,例如除了离散的听觉音调之外的可怕地方的所有其他线索。例如,LeDoux的老鼠开始害怕它们被测试的笼子,包括可能用于传递足部电击的钢条,以及地板上锯末的气味。如果笼子旁边有一扇带有明亮窗帘的窗户,老鼠可能会在看到所有窗帘时开始僵住。当它们听到实验室助理接近将动物从其家笼中取出以进行实验步骤时,它们也可能感到害怕。从我们自己的工作中,我们知道当研究人员接近时,老鼠通常会发出22-kHz的警报叫声,这些研究人员可能会将动物带到它们不喜欢的实验情境中。
虽然海马体对于许多情绪记忆的形成并非必需,但现在已经清楚,在将情境刺激(contextual stimuli)发送到始于中央杏仁核的恐惧”主干道”之前,海马体的参与是必要的。这是因为海马体除了在陈述性记忆(declarative memories)的形成中起关键作用外,还负责计算空间定位(spatial orientations),并提供关于条件反射发生情境的信息。这些情境信息也会直接从海马体传递到外侧杏仁核,以唤起恐惧系统(Ehrlich et al., 2009)。需要重申的是,当动物对如前所述的非常离散的线索进行条件反射时——一个特定的条件刺激——学习是通过不同的MGN到杏仁核通路发生的。
对情境刺激的广泛条件反射,显然增加了学习的适应价值。然而,这是通过杏仁核中略有不同的通路发生的,即通过基底核而非接收MGN输入的背侧核。但是,学习的神经生物学原理——产生线索和情境条件反射的神经元机制中的分子变化——仍然非常相似(就像其他初级过程系统中所有其他类型的基本情绪学习一样)。换句话说,谷氨酸传递效能的增加(作用于NMDA受体)对于情境恐惧条件反射和线索条件反射同样至关重要。
勒杜克斯的条件反射研究关注的是习得性情绪行为如何通过低级通路快速获得——这就是内隐情绪学习的发生方式。然而,他和大多数其他恐惧条件反射研究者并不承认,还存在一条通往原始恐惧感体验的进化”主干道”,一种非常厌恶的脑心(BrainMind)状态。正如罗斯福总统所理解的,我们唯一需要恐惧的就是恐惧本身。尽管我们钦佩这些条件反射实验的科学技巧,但当涉及到理解情绪感受的真正含义时,我们与勒杜克斯和许多其他进行此类研究的人分道扬镳。这是因为他们刻意忽视动物的感受,并且他们经常声称动物感受的存在与否是一个非科学问题(尽管在这些重大问题上有一些改变观点的迹象)。无论如何,正如我们在第2章中指出的,勒杜克斯明确支持读出理论(read-out theory)——即情感是由新皮层工作记忆功能创造的,这种功能在人类大脑中独特地扩展了。换句话说,他将情感视为高阶认知构造(也许只在人类中得到阐述),因此他将动物显著的恐惧反应设想为纯粹的生理效应,没有体验性后果(LeDoux, 1996, p. 302)。
现代对人类恐惧感的脑成像已经产生了许多关于杏仁核的有趣事实,但有一个事实突出:当研究者对那些仍然相当远离直接危险的恐惧进行成像时,杏仁核往往会亮起来。但当实验性的”捕食者”可以说就在你身后,准备咬你(即手指电击)时,恐惧系统的最低部分,即中脑导水管周围灰质(PAG),会亮起来(Mobbs et al., 2009)。这个脑区是恐惧感受和行为的中心(Brandão et al., 2008; Panksepp, 1998a)。当我们分析这里的电刺激在动物中的惩罚特性时,我们在最低水平的脑刺激下得到了可以想象的最强厌恶反应,而人类则体验到可以想象的最恐惧的心理状态(Graeff, 2004)。
这些情感体验的问题应该比现在更多地困扰恐惧条件反射研究者。我们认为,在当今时代,明智的做法是将原始情感体验的问题概念化为与动物是否对发生在它们身上的事情”自我意识”的问题截然不同,后者确实需要工作记忆并伴随对未来的巨大担忧。有力的证据表明,大脑中存在初级过程情绪网络,它们帮助在所有哺乳动物中产生现象性情感体验,也许在许多其他脊椎动物和无脊椎动物中也是如此。这些是心智脑(MindBrain)一些最重要的”非条件反应”的神经系统,对学习感兴趣的神经科学家必须经常使用它们,以便动物能够像它们那样容易地学习。我们认为产生这些体验的神经系统——脑心的各种奖励和惩罚——对于产生精神科医生和心理治疗师在日常工作中必须处理的习得性情感记忆至关重要。关于初级过程情绪的这些知识也使我们第一次能够理解情绪如何影响高级心理过程。
在总结恐惧条件反射时,让我们明确一点:只有神经科学研究才能真正阐明大脑中正在发生什么以及心理体验是如何产生的。心智是由复杂的大脑过程实例化的,这些过程在活体中运作,通常是在复杂的世界中。尽管如此,虽然行为分析在进行时几乎没有科学争议,但心理分析目前似乎只是重大争议的来源。我们认为这只是因为行为技术效果很好,而且大多数动物研究者觉得他们不必考虑心理构念,因为它们可能是我们想象的产物。情感神经科学通过识别帮助创造原始情感感受的关键大脑系统,通过各种大脑情感系统的奖励和惩罚特性,所寻求做的是提供一种理解我们情感心智基础的进化策略。大脑中基因内置的情感系统反映了祖先记忆—对生存具有普遍重要性的适应性情感功能,它们被构建到大脑中,而不是需要每一代个体重新学习。这些基因内置的记忆(本能)作为学习和高阶反思意识出现的进一步发展的坚实平台。
让我们分享一个关于恐惧所谓无意识本质的故事;这个故事广为人知,经常被那些假设上述恐惧学习在没有恐惧感受的情况下进行的神经科学家引用。“内隐情绪记忆(implicit emotional memory)”现象在1911年由一位名叫爱德华·克拉帕雷德(Edouard Claparede)的法国医生著名地证明了,他的一位女性患者大脑两侧的海马体(将短期记忆转化为长期记忆所需的区域)受损。正如现在所预期的那样,克拉帕雷德患者的大脑无法创建任何持久的陈述性-情景记忆。因此,一旦事情从她的工作记忆中消失,她就会忘记一切。每次她见到克拉帕雷德时,他都不得不向她介绍自己,就好像他们从未见过面一样。
见面时握手是惯例,因为患者能够保留程序性习惯,她能够参与这种社交仪式。有一天,克拉帕雷德在手掌里藏了一根针,在他们握手时刺穿了她的手指。疼痛让她吃了一惊,但伤口很浅,很快就愈合了。当然,她完全忘记了这件事。然而,当克拉帕雷德再次见到他的患者时,她拒绝与他握手。她无法解释为什么她如此不愿意,她做出了健忘症患者通常会做的那种借口来掩饰他们无法回忆事件的能力。(她说:“一位女士难道没有权利不把手给一位绅士吗?”)克拉帕雷德的患者不可能有意识地记得他在上次见面时伤害了她。她大脑两侧的海马体损伤使她无法生成事件的陈述性记忆。
然而,她确实从针刺中学到了一些东西。以前她不害怕与克拉帕雷德握手。握手是一种中性的,甚至可能是积极的体验。然而,在受到针刺后,患者获得了—学会了对以前中性的刺激产生负面情绪反应,即医生伸出的手,也许对医生本人也是如此,尽管她可能无法说出原因。对中性刺激获得新的情感反应构成了情绪学习。而对那种习得反应的保留就是情绪记忆。这个例子也很好地突出了情感体验在大脑中与陈述性认知体验非常不同。海马体的双侧损伤消除了后者但不消除前者。这个例子还突出了情感可以完全独立于认知。我们预期,如果克拉帕雷德或任何做过这类工作的现代科学家仔细询问他们的患者在这种恶作剧之后的感受,他们会说他们对研究者更害怕了一点,却不知道为什么(这可能只能通过理解情感可以独立于相关学习和认知而存在的临床医生的精神分析访谈来检索)。
情绪在决定我们如何行为、感知、思考和记忆方面的力量是一种非凡的特质。情绪使我们主动伸出手,通过身体和思维与世界互动。因此,我们倾向于将情绪系统设想为”吸引子景观”(attractor landscapes)(用非线性动力系统理论的术语来说),它帮助我们在思想和行为上与环境建立特定的联系。因此,我们设想初级过程情绪系统处于”有利位置”——占据上风——当涉及到学习如何控制我们大脑中记忆的形成时。这绝不是脑心智科学中的传统观点,主要是因为神经科学家通常忽视实验动物的感受。许多神经科学家尚未理解大脑中许多”非条件”过程的情感本质(UCS实例,例如疼痛性电击,以及UCR实例,例如非常可怕的恐惧唤起),他们使用这些过程来激发”条件性恐惧反应”(例如僵住和逃跑),这可以非常有效地用于研究动物的记忆。但他们与各种UCS实例配对的中性条件刺激可能之所以如此有效,是因为相关的UCR(神经系统的本能情绪反应)也”充满”了情感的神经化学物质。
事实上,存在一种内在的”记忆”过程,它完全是由于伴随初级过程UCR的重复情绪唤起而发生的——内在的情感记忆可能会对我们的思维方式造成内部破坏。如前所述,正如我们在继续学习过程中会一遍又一遍看到的那样,每个情绪系统都可以通过反复使用或长期不使用而变得更强(敏化)或更弱(脱敏)。我们内在情感心智景观的这种慢性变化使我们能够理解,重复的早期情绪体验(创伤)如何对成人思维的长期情感特质具有形成作用,我们通常称这些特质为气质或人格特质。
现代神经科学的显著之处在于,大多数研究者认为认知比我们的情感更能引导我们精神生活的方向。这在一定程度上是因为我们的新皮层非常能够发展知识结构,我们似乎在那里度过了生活中大部分的精神时刻。然而,当我们仔细观察时,许多这样的观点只是在理性和妄想之间摇摆不定的信念。完全”理性动物”的幻觉(关于精细分析,见Fogelin,2003)可能主要是由于我们是唯一拥有可以构建和传达复杂想法和妄想的语言的物种。许多人认为,没有语言我们就不可能完全有意识,但很多取决于我们对”完全”的定义。但从大卫·休谟到安东尼奥·达马西奥的几位哲学家和神经科学家认为,我们的情绪为我们的认知过程提供能量并引导它们。我们也属于其中。语言是人类大脑发展中最近的持久性发展,它的构建和引导既受文化影响,也受生物学影响。
通常阐述语言的神经机制使我们能够在认知上与他人交流(即,它们是擅长线性处理信息的习得工具)。这些机制通常集中在左大脑半球,它通常比右半球更少情绪化。相比之下,更具整体性和更具情感敏锐度的右半球维持和阐述生活的情感方面,几乎没有明确的认知意识,也许是因为它从情感和整体上看待生活。例如,它为我们的声音增添了韵律——情感旋律或情感刺耳性。
当然,否认认知活动在精神生活中的重要性是愚蠢的,但一个人以某种方式看待世界的大部分理由来自于一个人的感受。只需想想我们所拥有的信念——我们奇怪的、强烈持有的信念。如果没有来自集中在新皮层下方的情绪网络的情感和注意”能量”,许多认知观点的明显连贯性很快就会退化。
因此,同样重要的是要认识到,高级脑区可以被训练来调节情绪唤起,这是健康心智成熟的伟大成就之一(Goleman,2006)。再次考虑亚里士多德的智慧,在第4章的题词中。尽管认知和情绪在大脑的高级区域保持概念上的混合,但当这种相互渗透从下方和上方得到实质性的——温和而平静的——调节时,心理健康就会出现。这就是为什么认知行为疗法和正念疗法仍然是心理治疗领域的主流。心理治疗的一个目标,凭借其重新框定情绪困扰区域的能力,是帮助完善这些技能。另一个目标是理解我们精神生活的情感本质。心理治疗的大部分内容是一个人帮助另一个人理解我们思维的愤怒,并更加平和地对待大自然母亲和养育之父(或缺乏养育)在我们个人生活中所造就的一切。
仅仅几十年前,科学家们还相信每个人出生时大约有100,000个基因。我们现在知道每个人出生时只有大约22,000个基因。这当然足以创造我们所有人都具备的七个基本情绪系统。这些祖先的脑心(BrainMind)记忆还包括各种注意力和动机机制,我们不会重点讨论这些机制,它们由许多许多大脑化学物质控制。然而,重要的是要认识到,基因数量远远不足以解释我们心脑(MindBrain)功能的多样性和微妙性,但完全足够让婴儿动物和人类在获得成人心智的无穷特征的道路上顺利起步,其中大部分是通过学习创造的。在成年人中,个人特征的排列,更不用说个体的思维模式和专业知识领域,只能通过我们上面描述的巨大学习和记忆能力而出现。这些特征锚定在我们在本书中讨论的稳定的情绪技能和气质组合上。
然而,即使是这些情绪特质和人格维度也是通过我们在世界中的经历而巩固的。我们有许多直接从父母那里继承的优势和劣势,但也有很多是出生后从我们的养育方式中产生的基因变化——即大脑网络的表观遗传(epigenetic)塑造,导致大脑的初级过程情绪和动机网络中的各种敏化和脱敏模式。这些由环境长期塑造的基因记忆最近才进入发育思维的前沿(关于出生前这类效应的精彩近期概述,参见Paul (2010))。
表观遗传学(Epigenesis)描述了经验如何改变基因表达模式以允许创造许多个体特征的方式。表观遗传不是突变,突变是我们出生时所携带基因的物理变化。表观遗传是一种经验依赖的变化,发生在基因上,通常在我们出生后。基因可以改变的表观遗传方式之一是通过基因表达程度的变化。身体的所有细胞都有相同的基因,但在每个细胞中只有一些基因是活跃的或”表达的”。当以前休眠的基因变得活跃时,我们说发生了基因表达。基因表达导致新蛋白质的产生。当基因表达的强度在环境影响下降低或增加时,我们称之为表观遗传。
从化学角度来看,表观遗传产生于染色质的变化,染色质是围绕基因的支撑物质。乙酰化和甲基化的化学过程可以改变染色质的三维结构,这可以使转录因子(transcription factors)接触到基因(参见Szyf et al., 2008,我们将在第8章讨论他们关于母亲照料(CARE)对婴儿大脑表观遗传效应的工作)。当转录因子可以接触到休眠基因时,基因表达的速率可以提高或降低,以在新水平上产生蛋白质。大自然培育不同生命轨迹的另一种方式是通过生成称为”微RNA(micro-RNAs)“的小型调控基因控制片段,它们也可以帮助控制基因乐团在生命不同阶段的演奏方式。作为我们经历和展开的基因表达的函数而产生的蛋白质数量的变化,通常在创造新的神经通路中发挥关键作用。其中一些通路有助于编码新的技能、知识和人格特质。
当七个皮层下情绪系统中的任何一个被唤醒时,它们反过来会唤醒新皮层的部分区域。这有时涉及表观遗传和在大脑最具可塑性区域内创造新的神经通路。新皮层的表观遗传发育和专业化可能从根本上依赖于皮层下的注意力、感觉、情绪和动机功能如何帮助编织新的连接模式。在皮层下水平,其他表观遗传过程可能帮助塑造情绪系统的成熟,产生终生的情绪优势和劣势,这些决定了动物和人类一生的情感人格。
正如我们所指出的,在新皮层以下,大多数人——大多数哺乳动物——在情绪种类上非常相似,尽管在每个情绪网络的活力以及它们能影响的新皮层数量方面存在物种典型差异。我们性格的许多详细表观遗传精炼发生在新皮层中。然而,表观遗传也有助于解释我们的核心(皮层下)情绪气质是如何在发育中被精炼的——我们如何因为所处的社会和物理环境而成为我们现在这样的个体。表观遗传,连同学习/记忆,帮助我们理解为什么我们能够拥有如此复杂和多变的人格、技能和知识储备,即使我们每个人出生时只有约22,000个基因。除了传统的学习和记忆机制之外,心脑(MindBrain)不同区域基因表达的不同强度和模式,使每个人有限的基因库能够通过经验而多样化。
在本章中,我们试图传达学习和记忆的意义。在大多数情况下,学习和记忆是自动和非自主的反应(由大脑的无意识机制介导),但在几乎所有情况下,学习和记忆在其最持久的形式中,通常与情绪唤醒相关联。我们日常的工作记忆通常倾向于关注那些在情感上有意义的事物。更多时候,情节记忆和自传体记忆涉及我们生活中在情感上有意义的方面。最终,研究人员可能能够揭示一个分子和化学序列,从情绪唤醒开始,以创建持久的、高度个性化的情感性情节记忆结束,这些记忆影响我们作为独特个体的气质特征。说这些并不意味着没有一些不需要情感唤醒就能发生的学习和记忆类型,但我们不知道任何伟大的例子,除了机械记忆(我们在学校里讨厌的东西,进展特别缓慢,毫无疑问是因为无聊)和LTP型程序性记忆。
显然,情绪唤醒是创建恐惧学习记忆的必要条件——例如,对先前中性刺激的冻结反应——因为只有当训练程序唤起情感唤醒时,动物才能被条件化。情绪,尤其是寻求热情,也肯定会激励动物学习程序性技能。一般来说,这些技能需要重复练习,但我们需要被激励才能练习。然而,程序性记忆是情感唤醒对记忆检索非常重要这一规则的唯一例外,因为即使在人和动物情绪平静时,它们似乎也会持续存在。母亲可以有效地进行养育工作,而不会感到特别感动。虽然程序性记忆的检索可能不需要情绪唤醒,但它在情感调节以及他人情感调节中起着明确的作用。当孩子有礼貌时,这会让其他人喜欢他们。这对所有孩子来说都是积极的体验。当一位母亲即使在没有心情的时候也能有效地养育,这调节了她孩子的情感。
我们不了解认知心智的细胞和分子细节,但经典条件反射的神经科学(如上所述)揭示了关于大脑情绪和认知基质之间相互联系的一系列重要事实。大量实验证实了情绪对我们学习方式和思考方式的影响,部分通过我们工作记忆的运作方式(Davidson et al., 2003; Lewis et al., 2008)。这项研究表明,情绪唤醒在很大程度上决定了在工作记忆中自然处理的事物类型——当我们试图理解我们的生活时所考虑的项目类型。这是如何发生的尚不清楚,但有一些简单的可能性。例如,恐惧系统可能唤醒工作记忆,因为许多相关的神经通路(如由神经递质乙酰胆碱、谷氨酸和去甲肾上腺素驱动的通路,它们控制注意力的强度)被恐惧系统唤醒,并投射到介导工作记忆的新皮层相关区域(LeDoux, 2002)。
恐惧以外的其他情绪系统也可以以类似的方式引导一个人的思想和注意力,但关注点却截然不同。新母亲的照料系统会使她高度关注婴儿的任何痛苦迹象。在放松的时候,沉浸在养育的甜蜜中,母亲可能会思考要购买的可爱服装或琢磨温馨的照片。任何处于性欲系统唤醒状态的人都会发现他或她的思想和感知集中在性情境和各种惊人的可能性变化上。当愤怒被唤起时,我们倾向于沉思复仇的想法,并且太容易为我们的爆发找到认知借口(合理化)。当我们的恐慌/悲伤系统被唤起时,我们寻找一张友好的面孔,并怀着渴望的感觉思考快乐的团聚。我们的玩耍系统可以帮助我们成为认知小丑,用聪明的言语来取悦那些享受高级心智游戏的人。而寻求系统在正常生活的每个清醒时刻都在某种程度上被唤起,当其他更离散的情绪系统处于静止状态时,它为我们提供了学习世界的巨大能力。
考虑到情感体验和学习之间的这些关系,心理治疗师在更深层次的神经科学背景下考虑令人不安的记忆将非常重要,特别是因为再处理和再巩固可能会开辟新的和更好的机会来处理某些长期记忆的令人不安的情感方面(Panksepp, 2009a)。此外,考虑到情感负荷的记忆比非情绪性记忆更容易被自发记起,那些无休止地沉思他们烦人问题的人可能只会让事情变得更糟。这突显了他人的重要性,那些能够倾听并以敏感和积极的方式回应的人,可以帮助从更有益的情感角度重新构建记忆。事实上,现在人们可以想象全新的方法来软化创伤记忆对人们心理生活的破坏性影响(见第12章)。
以上所有知识的治疗意义是巨大的。这些事实似乎突出了关于情绪学习的一般原则,以及它们可能如何与相关的精神病理学(例如,广泛性焦虑障碍)相关联。情绪学习的额外层次甚至在大脑的更高区域中得到更广泛的呈现,可能以高度特异性的方式,这可能是使它们在治疗上如此难以管理的原因。但最终,所有这些关于恐惧条件反射的精细工作的实际价值在于,它让我们对病理性情绪困扰如何通过特定的生活经验在心智-大脑(MindBrain)中产生有了扎实的理解。理解这些机制为如何消除、重新条件化和重新情境化恐惧记忆开辟了新的途径。关于这些问题的大量工作正在进行中,总的来说,大多数研究者现在意识到”消退”——通过缺乏”强化”而导致习得性反应的丧失——也是一个主动学习过程。通过药物,如d-环丝氨酸(d-cycloserine),可以促进过去厌恶性记忆影响的消除和再巩固(通过新的学习),这些药物是学习促进剂(详细讨论见Davis等人,2006年;Myers & Davis,2007年)。这些药物是通过促进再巩固——情绪记忆通过新的情感情境和重新学习而被修改——还是通过消除旧的学习来起作用,目前尚不确定。
这些信息对于心理治疗师可能是有用和启发性的,因为它解释了情绪学习如何在人类生活中发生。所有动物,包括人类,在生命过程中都受到大量条件反射的影响,产生各种次级和三级过程的情绪现象。当条件反射回路成为记忆回路,特别是自传体回路时,情节性三级过程情绪记忆和思想的复杂性就会出现,与文化的持续流动和无尽复杂性融合在一起。反过来,当心智的皮层方面开始触发强烈且通常是特异性的情绪反应时,人们最好理解个体生活的动力学。这就是为什么治疗性对话如此有意义。当人们谈论他们的担忧时,他们会情绪激动并在人际间参与,这反过来又唤起陈述性和情节性记忆的进一步方面,为新的工作记忆提供”燃料”。思想和情绪在一连串的想法和感受中滚雪球般增长,这正是治疗性对话的特征。如果使用得当,这种对话可以成为持久情感变化的基础——一种深刻的脑-心智可塑性(BrainMind plasticity)(Doidge,2007年)——我们在精细的神经科学层面上还不太理解。但已经取得了显著的进展。
假如我们有足够的世界和时间,
小姐,这种矜持就不是罪过。
我们会坐下来思考以何种方式
漫步,度过我们漫长的爱情之日;…
用一百年来赞美
你的双眸,凝视你的额头;
用二百年来崇拜每一个乳房,
但用三万年来赞美其余;
至少用一个时代献给每一部分,
而最后的时代应该展现你的心。
——《致他羞怯的情妇》,写于1651-1652年,
由安德鲁·马维尔(Andrew Marvell)于1681年出版
从原始性欲到温柔之爱的性爱感受如何在人类大脑中产生,仍然是心智科学中最重要但最不为人理解的科学问题之一。理论很多,但共识甚少。相比之下,我们对啮齿动物大脑的性回路了解很多。通过这样的动物模型,可以解决关于原始性欲(LUST)的一些最有趣问题的细节,我们有希望阐明支撑人类性欲之爱的一般原则。动物研究发现对理解人类性行为(图7.1)的意义是巨大的。
LUST系统恰好位于我们理解基本哺乳动物身体驱力(性情感)和社会情绪之间的支点上,它们就像连体双胞胎一样紧密相连。LUST的原始冲动使社会生活变得极为有趣,因为性是所有哺乳动物生活中的主要动机,实际上是所有可能阐明人类状况的生物的主要动机。但性感受并不预示着我们自身的生存问题,而是关乎我们家族基因在未来世代的延续。性满足在经历这些体验的个体生存中不起关键作用;它们只服务于物种的生存……至少故事是这样说的。然而,令人满意的性生活很可能会促进强健的免疫系统和长寿,就像体育锻炼一样。尽管性满足可能不是生存的直接帮助,但关系良好、性生活满意的人往往比那些没有幸福关系安全感的人活得更长,无论他们的配对是男女、男男还是女女。充实的爱情关系所带来的祝福,以性满足作为坚实的组成部分,似乎是身心的良药,使许多人类能够在生育年龄之后活得比其他物种的常态长得多。但这在很大程度上也是有代价的:在共同生活中什么是重要和令人满意的问题上,女性和男性通常有不同的想法。关于这些问题的书籍不断被写出来,从20世纪70年代的《男女之间的心理战争》(Lewis, 1976)到世纪末的《男人来自火星,女人来自金星》建议(Gray, 1992),再到过去十年的《他到底在想什么?男人的大脑真正如何运作》(Gurian, 2004),强调女性应该了解许多男性相对较弱的情感生活,与许多情感丰富的女性相比。是的,男性大脑和女性思维确实有一些令人痛苦而有趣的区别特征。值得庆幸的是,我们这里的内容将主要关注大鼠,但我们将始终关注人类状况。

图7.1. Marcantonio Raimondi(1524年)根据拉斐尔才华横溢的25岁学生Giuliano Romano的原画创作的情色版画。这幅版画据说是最早的情色艺术收藏之一—“I MODI”系列中唯一幸存的图像。1524年,Pietro Aretino为Romano绘制的16种性姿势配上了十四行诗,这被认为是历史上最臭名昭著的情色艺术作品之一:Aretino的”情欲十四行诗”。这部作品与Raimondi精美的版画一起被编入印刷版。除了极少数副本外,这本书的所有副本都被教会迅速焚毁。目前已知没有完整副本幸存(来自http://www.artarchiv.net/doku/museum/Aretino.htm)。
那么作为一个有性别的男人或女人意味着什么?什么是自然的(或规范的)男性和女性性特征?当然,随着我们对人类中存在的”自然”多样性的文化意识的扩展,我们今天用来指代性身份多样性的语言和标签也扩展了。但当我们对人类性特征的了解实际上远少于对大鼠的了解时,关于词汇和标签几乎没有共识。这种不确定性有时可能是不适和分歧的来源,因为关于性别和性身份实质的文化战争仍在继续。例如,当所谓规范的男性和女性心理身份和行为倾向与身体外观混合在一起,有时几乎无法共存时,拥有双性身份意味着什么?真正成为跨性别者、变性者、同性恋者或双性恋者意味着什么?一个人的”性别角色”或”身份”是如何通过生物学、文化和个人选择的混合而创造出来的?
这些概念只是关于我们性遗产的混乱、嘈杂的辩论中的一些语义样本。这些辩论既是生物学的,也是政治化的文化—这创造了一个嘈杂的环境,在这个环境中,人们经常更相信理解不清的语言,而不是支撑我们性特征的生物学事实。在本章中,我们努力以相对直接的心理生物学和行为学方式使用性别和性身份的术语。诚然,我们的用法不一定在所有方面都符合当前文化话语中的用法(好像这样的事情甚至是可能的)。但由于我们试图尽可能紧密地讨论在动物模型中研究出来的初级过程证据,我们对性和性别术语的使用可能无法满足所有人的期望。我们请读者原谅这方面的任何明显不足,并加入我们这一章对跨物种情感神经科学探索所开启的非凡性特征视野的探讨。
在生物学层面,关于性和性别身份的本质,之所以会出现一些清晰度,是因为性别的身体外观和大脑的性组织可能与一个人假定的社会文化身份并不匹配。在生物学层面,多样性的范围可以从具有典型XX性染色体的遗传女性表现出明显的心理行为男性化,到遗传上典型的XY男性具有极其”典型”的女性身体特征,但内心感觉完全是男性化的这种模糊性。事实上,人们可以处于各种”不匹配”的身体和心智之中。当我们加上更高层次的心理自我身份时,可以肯定的是,事情变得无限复杂,在许多层面上存在概念上的激流和文化上的交叉流,从我们所说的心智脑组织的初级到三级层面。我们甚至不会触及受文化和社会影响的”性别角色”身份问题,这些问题为初级过程的神经生物学和荷尔蒙复杂性增加了如此多的三级过程复杂性层次,而这将是我们的关注焦点。人类文化问题无法在其他动物身上得到很好的模拟。然而,通过研究我们的哺乳动物同伴,性冲动和性别身份的许多生物学特征已经得到了阐明。我们认为,这种共享的生物学遗产对我们人类和其他哺乳动物在初级过程层面感受情欲的方式的影响是深远的。
本章将探讨男性和女性大脑之间的解剖学和化学差异。我们还将探索一些现代神经科学研究,这些研究确认了为什么性欲(LUST)回路应该被视为一个初级过程情绪系统,一个对女性和男性来说有些不同的系统。像所有其他情绪系统一样,性欲网络与各种稳态和感觉情感机制相联系。例如,饥饿会极大地降低性冲动,恐惧和大多数负面情绪感受也是如此,尽管轻微的疼痛有时可以增加性唤起,而不仅仅是在施虐受虐的人类中(Caggiula & Eibergen, 1969)。但主要的是,我们将深入研究一些关于性大脑和性身体表现出相对独立的胚胎发育方式的迷人研究。这种独立发育在很大程度上解释了同性恋/跨性别现象的极端情况,包括遗传(XY)男性可能更温和的心理女性化特征梯度和遗传(XX)女性的男性化心理。我们还将探讨促进跨性欲望的心智脑机制,那些似乎违反男性和女性身份简单化概念的机制。在此过程中,我们将关注一些相关的临床考虑。尽管我们对人类多样化跨性别现象背后的大脑解剖学和化学特性的具体知识有限,但我们对动物模型中的这些问题已经相当了解。至少在科学界,人们越来越接受在其他动物身上发现的生物学事实可以阐明相关的人类状况。作为这一讨论的一部分,我们还将研究一些关于催产素作为”爱的荷尔蒙”的新兴神话——这通常有几盎司的真实性,但往往也有几磅的夸张。“信心”的概念可能比”爱”的概念更能解释催产素的影响。我们将通过考虑神经科学和经典精神分析的心理性发展理论之间存在的鸿沟来总结。
有些人仍然相信男性和女性的大脑是相似的,男性和女性的性偏好完全是习得的。现代神经科学已经提供了充分的证据来消除这些观念。虽然每个性别确实都有男性和女性大脑网络,但这些回路通常强度并不相等。因此,正如男性和女性的身体在某些重要方面不同一样,他们的大脑在许多方面也是不同的,具有极其多样的心理细微差异(Hoyenga & Hoyenga, 1993)。图7.2总结了大鼠潜在性回路的示意图,可能在大多数哺乳动物中是相似的。
在雄性哺乳动物中,原始性冲动的中枢位于下丘脑前部的内侧区域,尽管不同物种涉及的确切大脑位置和术语略有不同——在大鼠中,是性双态视前区(POA),而在人类中是下丘脑前部的间质核(INAH),它们在进化上肯定是相关的。正如Phoenix等人(1959)在豚鼠中首次阐明的,并很快扩展到恒河猴,大脑性回路的组织始于胎儿期,最终导致年轻男孩和女孩在许多三级加工兴趣上存在差异。这主要由睾酮控制,睾酮在出生前后的婴儿期分泌。然后在青春期女性中,卵巢雌激素和孕酮类固醇产生的成熟预示着青春期的到来。而在青春期男性中,当睾丸开始产生大量睾酮时,就会发生强烈的性觉醒。这些决定性的女性和男性激素与大脑许多皮层下性区域的各种类固醇受体结合,特别是下丘脑的前部。这种结合产生愉悦的情感,至少对睾酮来说是这样。因此,雄性大鼠显然喜欢在大脑的POA中注射睾酮。它们会为此而努力,并会重访那些它们的性回路被睾酮注射”润滑”过的地方(King等,1999)。
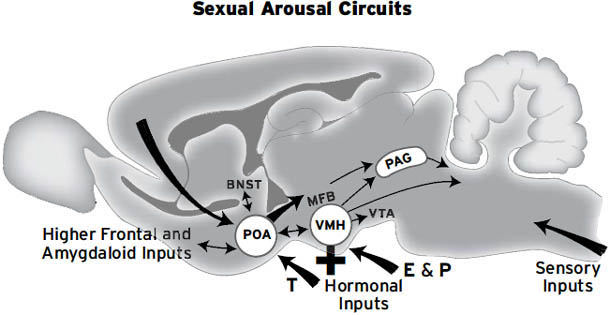
图7.2. 大鼠大脑的侧视图,总结了对雄性和雌性性行为提供差异化控制的主要皮层下网络。雄性含有更大的POA,该区域对雄性性能力至关重要。VMH(腹内侧下丘脑)在雌性性反应中显然更具影响力。这些系统部分通过敏化各种感觉输入通道来促进交配反射。这些回路在多大程度上控制性行为的情感成分仍不确定(改编自Panksepp,1998a;经牛津大学出版社许可重新出版)。
这些循环类固醇很容易进入大脑。神经化学的一个重要原理在这里发挥作用,应该牢记——激素和其他液态神经化学物质只是完成这些化学物质在整个神经系统中传递的信息的一半等式。大多数溶液中的神经化学物质,在细胞内和细胞间空间移动,只有在神经系统相关区域的细胞膜中,甚至神经元内存在相应的受体分子时,才能发挥其特征性效应。睾酮对男性大脑产生更大影响,因为男性在下丘脑前部的关键区域——特别是POA——拥有富含睾酮受体的更大神经元场。这几乎就像男孩在大脑内有一组更大的性腺,对应于更明显的外部性腺。女性性行为,正如我们将看到的,具有更复杂的基础,这些基础还没有被很好地理解。
男性大脑中神经符号性腺的概念与现实相去不远。正如在动物模型中特别深入研究的那样,睾丸或这些关键的下丘脑前部区域的损伤会产生相似的效果。例如,POA的损伤会显著削弱雄性的性冲动和能力,特别是如果损伤发生在性未成熟动物的青春期之前。如果年轻动物在达到性成熟之前失去睾丸,它们将永远不会发展出强烈的性冲动,除非将睾酮引入其大脑的适当区域。然而,此类动物的更一般的社交冲动并未受损,这表明社交需求并未完全束缚于性需求。此外,一旦成年雄性大鼠发展出性习惯,同样的POA损伤对雄性性行为的破坏不如对无经验动物那样严重。推测性动机下降得更慢,因为动物已经发展出高阶性习惯,这些习惯有了自己的生命(即,动机在二级和三级加工水平上得以维持)。通过生活经验,性动机已经转移,但只是部分地,转移到大脑的其他区域。POA损伤后,有性经验的大鼠仍然继续努力获得接受性雌性,但它们的性后续行为相当迟缓。这些经验可能也适用于人类,但相关的科学数据(特别是相关的大脑数据)很少。失去睾丸的性活跃男性往往比去势动物维持更长时间的性生活,但动机的逐渐下降是不可避免的。在其他动物中,下降通常要快得多(Meston & Frohlich,2000),大概是因为它们有更少的高级大脑机制来维持动机。
为什么睾酮在激发男性性欲方面如此强大?由睾酮激活的各种神经肽肯定发挥了作用。研究最充分的是加压素(vasopressin),在动物模型中,它促进性热情、求偶、领地标记、雄性间攻击性,以及可能的性嫉妒(Goodson & Bass, 2001; Hart & Legerstee, 2010)。睾酮在大脑中的奖赏效应是否是由于促进了加压素回路,目前尚不清楚。无论如何,男性的加压素含量是女性的两倍。睾酮还会激活大脑中的一种气体递质,一氧化氮(NO),它促进性渴望的提高以及男性典型的攻击性增强——这是一种公认的依赖睾酮的”攻击性”形式(Nelson et al., 2006)。目前的男性性功能增强药物,如伟哥,能够比过去任何催情剂更稳定地改善勃起功能,因为它们提高了大脑和阴茎中一氧化氮的产生,也许在阴蒂中也是如此。我们再一次看到,身体和心智如何在相似化学物质的影响下协同工作。
除了促进男性性反应外,睾酮在产生男性对社会支配地位的攻击性冲动方面也起着至关重要的作用。现已确定,男性性行为和对支配地位的自信冲动在大脑皮层下区域有相当程度的相互作用,而且很明显,这种相互作用在很大程度上根植于高水平的睾酮。换句话说,睾酮为促进男性性行为的回路提供动力,同时也为促进社会支配欲望的许多大脑机制提供动力。正如我们在第5章中提到的,睾酮还可以使大脑的愤怒(RAGE)回路敏感化。无论是人类还是大鼠,雌性在获得睾酮后会以类似的方式做出反应——它们变得更加自信和自信,同时对他人的怀疑也增加了(van Honk et al., 2004)。大概女性通常更信任他人(或在社交上更自信),因为她们的催产素水平升高,而睾酮对男性的影响——男性更容易沉迷于攻击性和更微妙的支配性表现——也促进了对他人动机的更大怀疑。在性行为的小鼠模型中,某些气味输入到雌性大脑往往会抑制其大脑中可能促进典型男性行为的回路(Kimchi et al. 2007)。
然而,尽管睾酮对男性性行为和男性常表现出的强化攻击性都至关重要,但社会支配的冲动不太可能简单地等同于男性性冲动。有许多理由相信,性行为和攻击性是大脑中可分离但相互作用的系统,特别是在内侧杏仁核(medial amygdala)等皮层下区域。杏仁核中的一些神经元只对性行为做出反应,另一些则对攻击性做出反应。然而,也有许多神经元对性和攻击性情况都做出反应。同样,对人类的研究表明,颞叶区域(攻击性回路集中的地方)在男性中更活跃,而前扣带皮层区域(养育和社会疼痛回路集中的地方)在女性中更活跃(Gur et al., 1995)。
当男性性欲没有令人满意的出口时,它会产生令人不安的紧张感,其中一些会将生物体带到竞争领域和各种攻击性遭遇中。从统计学上看,男性对女性的身体虐待往往多于相反的情况。这部分是因为睾酮激发了心智中对社会支配地位的攻击性冲动。事实上,如果原始的欲望冲动得不到满足——在文明社会中往往得不到满足——青春期外周性激素的激增可能会促进巨大的性挫败感。当然,这种内在力量会导致性攻击和其他社会不当的行为。
我们通过研究其他动物学到了很多关于性激情的知识。例如,这些与性别相关的气质差异部分是由于性激素影响大脑化学的方式:雌激素在雌性大脑中”滋养”催产素(oxytocin)神经系统,而睾酮则增加男性大脑中加压素的力量。正如我们将在下面讨论的,催产素对大脑产生镇静作用,这似乎促进了男性和女性积极社会联系的形成。另一方面,加压素倾向于诱发男性的竞争性,但它也可以增加他们的性联结和防御性(嫉妒?),而在女性中它通常会降低性渴望。正如我们将在后续章节中看到的,这些性别不同的神经肽控制着哺乳动物的许多非性社会行为(Goodson & Bass, 2001)。鸟类肽类,催产加压素(vasotocin),结合了催产素和加压素的功能,它与内源性阿片类物质一起,强烈地控制着鸟类的社会动机(Panksepp, 1982),特别是鸟鸣,一种长期确立的性-领地反应(Riters, 2011)。
关于雄性攻击性和这些肽的大部分证据来自对鸟类的研究,特别是关于禽类加压素(avian vasotocin)作用的研究,它在化学结构上与催产素和加压素仅相差一个氨基酸。加压素显然能平复情绪困扰并促进平和。事实上,我们发现加压素可以减少雄性的支配行为,而不改变它们明显的支配感受。正如我们在第一章中提到的,我们在成对雄性鹌鹑身上观察到的一个相当奇特的发现说明了这一点(Riters & Panksepp, 1997)。雄性鹌鹑在初次相遇时,会相当猛烈地啄对方的头部,显然是试图解决支配关系。但当给其中一只雄性鹌鹑直接向脑室注射加压素后,这只鸟几乎没有表现出相应的攻击性,似乎从容地承受每一次恶意的啄击。值得注意的是,当这些以前温和、“接受攻击”的雄性随后被给予对照溶液时,它们立即开始啄击它们假定的”主人”,情况发生了逆转。
这个结果确实相当奇怪!在正常情况下,服从啄击的动物是顺从的一方。因此我们假设加压素诱导了顺从倾向——我们预期这些倾向会持续存在。然而,加压素加上早期的啄击,并没有使鹌鹑真正变得顺从。那么为什么接受加压素处理的鹌鹑会服从如此凶狠的啄击呢?也许加压素产生了一种自信和平和的感觉,作为一种情感上的”保护”,防止之前的社会攻击固化为持续的顺从态度。通过这种基础的跨物种情绪神经科学研究,一旦我们在智识上能够考虑神经化学物质可以创造的情感变化,许多看待行为控制的新方法就会出现。
在许多动物中,雄性性行为的活力和雄性自信的必要性(即社会支配行为)在争夺交配权的复杂仪式中汇聚。例如,在许多群居动物中,只有占支配地位的雄性才有与雌性交配的特权。此外,在许多动物中,雄性具有生殖适应性的明显标志,雌性在选择配偶时会关注这些标志。突出的例子包括孔雀华丽的尾羽、山魈的面部颜色,以及狮尾狒狒的胸部斑块颜色(几乎象征着一个人的心向女性关注敞开)。雌性的接受性通过许多因素来表达,例如特别吸引人的气味或引人注目的颜色,如黑猩猩臀部的红肿。事实上,一些人类男女差异,特别是在具有精神分裂症倾向的个体中,突出了精神病学重要性的问题(Goldstein, 2006),这可能为理解分裂人格赋予新的意义。这些心理方面在人类中注定是独特的。
我们人类认为自己在性偏好方面很成熟。但男性很容易被年轻美丽的女性所吸引。女性美貌确实与各种面部和身体特征有关,进化心理学中最著名的一个是”沙漏”身材的数学比例,腰臀比为0.73,这对许多男性来说正中”要害”(Singh & Randall, 2007)。那个”要害”在哪里?也许不在视前区(POA),而在视觉系统的高阶情感感知中。也有证据表明男性和女性都存在信息素气味信号(Savic et al., 2009),但我们不会在这里详述那些仍有争议的复杂问题。人类女性,像许多其他雌性动物一样,通常被占支配地位的男性所吸引——在人类中,这些通常是那些富有、有权势(不仅仅是肌肉发达,尽管这可能有帮助)或在其他方面有成就的男人。言情小说中充斥着这样的男人,主人公并不总是最温和的角色。(我们只需考虑艾米莉·勃朗特的《呼啸山庄》中引人注目但情感上残酷的希斯克利夫。)言情小说当然是为娱乐而非启蒙而设计的逃避主义文学。但这些书一贯以占支配地位的英雄为特色,并且对许多女性来说是一个长盛不衰的吸引点,这表明它们触动了女性性爱浪漫想象中的原始和弦。女性可能被占支配地位的男性所吸引,是因为他们显示出的选择优势——既体现在传递给个体后代的遗传遗产方面,也体现在为几个后代组成的家庭提供保障的能力方面。
社会支配互动在雌性中不如雄性那么强烈或明确,因为雌性大脑不像雄性那样受到循环睾酮对脑回路影响的强烈支配。但我们会看到,人类女性的性冲动也在很大程度上受到这种”雄性激素”的影响,如果女性拥有和男性一样多的睾酮,她们可能会和男性一样具有攻击性。事实上,有一个引人注目的例子支持睾酮在社会支配中的作用规则。雌性鬣狗第二性征和成年社会行为的独特发展证明了睾酮与社会支配性攻击之间的联系。与大多数其他哺乳动物不同,雌性鬣狗具有异常高水平的循环睾酮,它们的外生殖器与雄性的惊人相似。人们不容易区分性别,因为雌性扩大的阴蒂与雄性的阴茎大小相同,而且阴蒂完全能够勃起。雌性似乎展示它们的生殖器作为一种性诱惑雄性的方式,也作为社会支配展示的一部分。
实际上,雌性在鬣狗社会中扮演着强大的角色,并且始终占据着对雄性的支配地位。雌性鬣狗的这种反常发展突显了从睾酮到支配性攻击行为的因果路径。高水平的这种激素是雌性鬣狗社会支配地位的关键。在人类女性中,或许令人惊讶的是,人们观察到单次剂量的睾酮会增加攻击性和其他男性典型的心理特征(Bos et al., 2010; van Honk et al., 2004),并有丰富的脑成像研究支持这类类固醇对大脑的影响(van Honk & Pruessner, 2010)。
女性性欲回路已在多种实验室动物中得到详细研究。这些回路与在雄性中发现的回路明显不同。雌性性接受的冲动起源于腹内侧下丘脑(VMH),这是下丘脑的不同部分,而雄性的冲动则关键依赖于POA——前下丘脑的视前区。对大脑VMH区域的损伤会严重削弱雌性的性接受能力,而这种损伤对雄性大鼠的性行为影响相对较小,只要它们没有变得过于攻击性。
如上所述,大多数雌性哺乳动物不会产生大量睾酮(尽管它们的肾上腺会制造一些)。在大多数哺乳动物中,雌性的性准备状态主要由雌激素和孕酮控制。但正如我们将看到的,肾上腺睾酮对雌性接受能力贡献很大。大多数物种中的雌性性唤起由规律的发情周期控制,而发情周期又由雌激素和孕酮释放和相互作用的严格时机控制。发情周期从下丘脑分泌激素开始,这些激素通过垂体腺——悬挂在下丘脑最底部的一个小附属物——导致卵巢准备就绪。垂体腺通过分泌促性腺激素释放激素(GnRH,也称为黄体生成素释放激素,或LH-RH)参与发情周期。这反过来诱导卵巢在卵巢”卵子”成熟时分泌雌激素,随后当成熟的卵子溢出到输卵管时会出现一个孕酮脉冲,准备受精。
雌激素和孕酮还通过促进催产素的制造,使雌性在情感上对追求者的示好更具接受性和信任感,催产素是介导雌性性准备状态的关键化学物质。雌激素激活下丘脑神经元中休眠的催产素基因,导致大脑催产素产量升高。雌激素和孕酮还促进VMH中催产素受体场的戏剧性扩张,几乎像春天草地上绽放的花朵。催产素产量增加和受体增殖的结合导致唤起信号,这些信号通过脊髓中的回路启动并支持脊柱前凸反射。这种反射引起一种性接受的身体姿势,包括下背部向上弯曲,从而暴露生殖器(Pfaff, 1999)。尽管脊柱前凸反射在下脊髓中组织,那里也有丰富的催产素受体,但它受到起源于VMH的下行神经通路的强力促进。因此,当许多哺乳动物物种的雌性性唤起时,它们会采取一种促进交配的姿势。据我们所知,在人类大脑中,这种准备状态更多地反映在心理生活的特质中,但这可能只是因为身体姿势尚未得到充分研究。
在大多数哺乳动物中,性接受能力通常仅通过雌性身体的诱人气味就能明显看出。雄性大鼠可以迅速从一百只不接受的雌性中”嗅出”一只接受的雌性。然而,如前所述,雌性性吸引力的细节在不同哺乳动物物种之间差异很大。例如,一些物种没有发情周期。这类物种是反射性排卵者,这意味着性行为本身会触发卵子从卵巢释放。人类女性确实有心理发情周期,但从表面上看,女性主要是隐蔽排卵者,缺乏许多其他物种表现出的明显性接受迹象。对于女性来说,接受能力更多的是一种心理状态,部分反映在性唤起能力的”月度”波动中。尽管如此,某种接受能力的潜力始终存在,响应各种社会变量,但不像男性那样强烈。
女性的情欲心理状态受到大脑化学的周期性变化影响。在生育高峰期,当雌激素和孕酮都很高时,女性的思绪比这些激素水平较低时更容易转向情欲幻想。这部分是由于对大脑回路的直接影响,尽管也存在多种间接影响,如对大脑催产素系统波动活动的影响。正如社会学家们准备强调的,从其他动物中收集的生物学故事对人类的适用性是有限的,尤其是女性,她们在性事务上表现出更多的选择、辨别力和微妙性(Udry, 2000)。尽管如此,许多潜在的神经化学原理在哺乳动物物种之间很好地普遍适用(Meston & Frohlich, 2000),但每个物种总是有一些独特的方面,包括人类女性性行为对睾酮的高度敏感性。
在人类中,肾上腺睾酮为女性的性欲增添了一种欲望成分,这种成分对于大多数其他物种的雌性来说并不那么重要。这几乎看起来像是在人类女性的性欲方程式中添加了一小部分男性的性冲动。由于女性性欲的这一组成部分,补充睾酮正被证明是恢复已到更年期女性性欲的有效方法(Al-Azzawi et al., 2010)。许多年长女性不需要这种补充,也许是因为她们的情欲-关爱态度是在一生中发展起来的,并且主要在习得的次级过程和心理维持的三级过程层面上运作。当然,对于满意的性生活来说,无论是男性还是女性,最重要的因素是心智的情感质量。正如我们在第1章中提到的,催产素可能会增强关爱情感,但除了催产素之外,肯定还有许多其他化学物质促进内在体验和外在表达的性欲热情。然而,催产素在哺乳动物中已经得到了特别深入的研究(Uhl-Bronner et al., 2005),以及鸟类中的祖先性物质加压素(vasotocin)(Panzica et al., 2001)。
有趣的是,催产素这种女性性欲的关键化学物质也可以唤起男性。给雄性啮齿动物大脑施用催产素,也许还有在人类男性大脑内分泌催产素,都会产生勃起。此外,在射精期间会大量分泌催产素。另一方面,加压素强烈促进男性的性欲(也许甚至是强迫性),但对女性却有相反的效果。如果将加压素注射到雌性大鼠的大脑中,她的性接受能力会被严重抑制。尽管如此,雄性和雌性大鼠都享受性。雄性会迅速对它们进行过性行为的地方产生偏好。虽然雌性大鼠也表现出这种地点偏好,但通常只有在允许她们自己控制性活动节奏的情况下才会这样(Pfaus et al., 2003)。即使是雌性大鼠也不喜欢被强加性行为。不过,如果人类研究者人为地刺激雌性大鼠的阴蒂(用振动器尖端的棉签),她会对雄性大鼠变得更具性索求,更容易怀孕,并且也会对她接受这种刺激的地方表现出偏好(Cibrian-Llanderal et al., 2010)。这种阴蒂刺激的奖赏特性似乎源于大鼠视前区(POA)的激活(Parada et al., 2010)。
尽管有许多有趣的差异,两性在初级过程层面上也有许多共同点。我们在上面提到,每种性别的大脑都包含异性典型的残余性回路。因此,雌性大脑中也发现了加压素回路,但数量较少,而雄性大脑中存在催产素回路,但数量也较少。这些回路可能如何运作?我们推测,雌性大脑中的加压素系统可能有助于激发母性行为中一些更具攻击性的方面(例如,保护幼崽免受伤害);相反,催产素系统可能维持男性行为中一些更温和的方面(例如,父亲对其后代表现出非攻击性和支持性的倾向)。无论如何,尽管不同物种在管理性欲方面存在许多差异,但在初级过程层面上,似乎所有哺乳动物都共享非常相似的欲望回路(Pfaus et al., 2003)。
重要的是要认识到,就像追求其他各种奖赏一样,寻求系统也被招募来执行寻找性伴侣的任务。这意味着,除了上面提到的性化学物质之外,性欲望和渴望还由多巴胺驱动的寻求系统促进。在人类社会中,多巴胺驱动的对陪伴的寻找以无数方式得到促进:牵线搭桥的朋友、单身酒吧、约会中介和互联网等等。事实上,无论一个人是通过互联网寻求智力知识还是肉体知识,驱动这一行动的都是寻求系统。正如我们在第3章中强调的,这种多巴胺驱动的参与充满情感(欣快的)行动,在寻找所有环境乐趣包括性方面都发挥作用。有一些证据表明,这个系统在男性中可能比在女性中更加活跃,但这一观察结果可能主要是特定情境的——更多地由特定奖赏、生存职责和动物所处的生态限制所决定。例如,当母鼠收集从巢穴中分散的幼崽时,正是催产素激发的寻求系统启动了大部分工作。雄性不是特别渴望承担这样的任务,所以就关爱而言,寻求系统在雌性中似乎更具反应性。我们知道寻求系统会被敏化——变得过度反应——面对各种生活挑战,包括压力、饥饿和滥用药物。当它被敏化时,动物更渴望追求各种奖赏——食物、性接触和药物诱导的刺激(Nocjar & Panksepp, 2002)。因此,尽管在许多情况下寻求系统在雄性中似乎比雌性中稍微更活跃,但在其他情况下情况正好相反。
虽然男性和女性之间存在许多神经化学差异,但催产素(oxytocin)和加压素(vasopressin)构成了特别重要的情感区别。很明显,这些激素作为社会性性肽具有广泛的作用,因为它们与更高级的心理过程相互作用。这意味着这些肽促进的心理特质在每个性别中的权重不同。催产素鼓励女性典型的养育态度,可以用”照料和友善”这个短语来概括(Taylor et al., 2000)。加压素使动物转向男性典型的态度,可以用”强势和竞争”这个短语来概括。根据经验法则,成年雌性大鼠的催产素活性是雄性的两倍,用以影响它们的大脑、心智和行为,而成年雄性的加压素是雌性的两倍(这是否适用于人类尚不清楚)。性别之间还存在许多其他的大脑心智差异。然而,最大的内在差异存在于情感层面,而在认知层面上明确的差异相对较少。大多数认知差异——已经记录了许多——可能主要源于后天培养而非先天本性。
性别之间还存在其他情感差异。如前所述,雄性通常具有更强的寻求(SEEKING)和愤怒(RAGE)冲动,在某些物种中可能还有更强的玩耍(PLAY)倾向。雌性哺乳动物通常具有更强的照料(CARE)和分离痛苦(恐慌/悲伤,PANIC/GRIEF)反应性(这解释了它们在婴儿养育方面的更高投入倾向)。虽然雌性似乎也更容易表现出恐惧(FEAR),但母性的荷尔蒙变化明显增加了信心。这种母性信心的提升可能部分源于大脑催产素水平升高的心理效应(Panksepp, 2009c)。
正如在引言章节末尾所指出的,催产素已经吸引了大众的想象,被称为”爱的激素”,这是对大脑运作方式的一种过度简化。确实,催产素在性行为和其他积极情绪中发挥着重要作用。它增强高潮的强度,在分娩过程中,它增强母亲完成将婴儿推向世界这一艰巨任务的能力。它还有助于缓解伴随情感和身体痛苦的疼痛。最重要的是,它在面对抚养孩子这一艰巨任务时促进女性的信心。但催产素并不是单独作为”爱的化学物质”发挥作用。它在许多其他大脑化学物质和环境输入的协助下运作。
动物研究表明,单独作用的催产素不会产生强烈的积极情感状态。一项研究确实产生了适度成功的条件性位置偏好(CPP)(Liberzon et al., 1997)。然而,在Panksepp的实验室中,只有当催产素在社交环境中给予动物,使它们能够进行友好互动时,才能获得CPP(1990年代的未发表数据)。换句话说,它们的催产素诱导的位置偏好与积极社交体验的可用性相关。因此,催产素单独可能不会直接产生强烈的积极情感体验,而只能从同时发生的社交互动中产生;因此,它可能增强其他更直接促进积极社交感受的大脑化学物质的作用,例如内源性阿片类物质(由大脑产生的类似鸦片的化学物质),当人和动物进行友好社交互动时也会释放这些物质(见第9章)。内源性阿片类物质是主要的”安慰和快乐”化学物质,当给予阿片类物质时,很容易获得CPP,无需添加社交互动(Tzschentke, 2007)。
正如我们上面指出的,催产素可以增加大脑对阿片类物质的敏感性(Kovács et al., 1998)。当动物进行友好互动时,内源性阿片类物质会在它们的大脑中释放(Keverne et al., 1989; Panksepp & Bishop, 1981),也许催产素增强了这些阿片类物质引起的愉悦感。随着更多工作的进行,以确定各种社会促进的CPP能否被阿片受体拮抗剂(如纳曲酮)有效减少,我们将更多地了解这一现象(见Panksepp et al., 1997)。
在这种情况下,应该强调的是,每当我们谈论这些社会神经肽时,我们都在谈论中枢(大脑)效应。虽然催产素和加压素也是对压力有强烈反应的外周(血液)激素,但几乎没有证据表明这些激素从垂体后叶释放后能够返回到大脑的社会性性回路中。事实上,从社会神经科学的角度来看,外周激素水平可能产生矛盾的效果。例如,如果监测处于困扰中的人类夫妇的紧张水平,血液中的催产素水平(而非加压素)与女性的压力水平正相关,但与男性无关;相反,在男性中,加压素水平(而非催产素)与社会压力高度相关(Taylor et al., 2010)。我们应该假设催产素和加压素都会在大脑中引起压力吗?不。这些激素的外周测量并不能提供关于其中枢效应的明确信息。事实上,也许催产素被压力事件释放到循环系统中,是为了抵消对身体的负面影响。
无论大脑催产素(oxytocin)是否直接诱导积极情感,大量实验都证明催产素在积极的社交互动中起着至关重要的作用。例如,当幼年动物与母亲分离时,催产素会抑制痛苦的哭泣,这表明催产素在动物独处时提供情感安慰方面发挥着关键作用(见[第9章])。这种效应不需要阿片类物质(opioid)的参与。此外,中枢和外周给药的催产素都能减少雄性大鼠杀死幼崽的倾向。性交后会分泌催产素。正如我们在鹌鹑的例子中看到的,这类分子可以唤起平和的倾向。在大鼠中,这种效应似乎在大约3周后达到峰值——这正是妊娠期。众所周知,在这个时候,当它们自己的幼崽即将出生时,父鼠会失去杀死幼鼠的冲动。因此,它们的性行为似乎已经在大脑中注册,使得父鼠极不可能杀死自己的后代。
大量研究结果表明,在动物大脑中注入额外的催产素可以促进雌性大鼠的母性情绪,即使是未生育的雌鼠也是如此(见下一章关于照料系统CARE)。通过给未生育雌鼠注射哺乳母鼠的血液也可以达到同样的效果;这一结果是否由催产素引起尚不清楚。无论如何,催产素促进了母亲与孩子之间产生的强烈社会纽带(“纯粹的爱”?)。Keith Kendrick及其同事在绵羊母亲身上对这种效应进行了最彻底的研究(Kendrick et al., 1992)。
催产素活性和催产素受体在大脑中的特定分布也促进了某些田鼠物种成年个体间的配对结合(pair-bonding),特别是研究充分的草原田鼠(prairie vole),它们倾向于形成家庭群体,成年雄性和雌性之间具有稳定的性介导配对纽带(Carter et al., 1995)。这里的”稳定性”并不意味着”专一性”;几乎所有配对结合的动物都会表现出一些”配对外交配(extra-pair copulations)“,这是一种科学的说法,意思是如果可以的话它们会乱搞,但不会与临时配偶发展持久的纽带(承诺)。研究表明,草原田鼠之所以形成配对纽带并且特别喜欢群居,是因为催产素受体在其大脑中的特定分布。另一方面,另一种田鼠——山地田鼠(montane vole)过着像隐士一样的独居生活,其催产素受体分布在大脑中相当不同但相邻的区域(Insel & Shapiro, 1992)。
但我们仍然不太确切地知道催产素到底产生什么感觉——是感觉更有爱心、更自信,甚至可能更勇敢同时攻击性更低?还是它只是帮助人更放松和悠闲?也许所有这些都是真的。我们将在下一章回到这些困难的问题。
但鸟类也可能提供线索。对鸟类的研究表明,加压素(vasotocin,以及催产素)可以促进自发的扇翅动作,这可能表明信心,特别是因为这种效应在其他鸟类存在时会显著增加(Panksepp, 1992)。事实上,在大约20年前未发表的工作中,Panksepp直接评估了这个想法。八只幼鸡组成的群体接受大脑脑室内注射催产素或安慰剂,通过观察它们在一个大型光线充足的新房间里分散的距离来测量它们的信心,在被桶限制一分钟后可以自由探索。当桶被抬起时,催产素处理的小鸡比安慰剂处理的鸟类在房间里探索得更广泛,这表明催产素处理的小鸡感到更安全——这相当于信心增强。
大多数动物研究没有充分考虑的是,如何从原始情感变化的角度来解释这些不同的发现。相反,太多的动物研究仍然只关注行为变化而绕过情感考虑。我们应该在此类实验中寻求识别原始情感过程的一个原因是,催产素能系统仅存在于哺乳动物大脑的皮层下区域——大脑中产生情感性情绪的部分。如果我们正确理解了初级过程的解释,那么我们将能够更好地理解人类催产素研究中常用的高阶情感概念,这些概念我们经常用来理解人类情绪。
人类的依恋关系,无论是性的还是其他方面的,都是微妙而复杂的,具有巨大的认知复杂性层次。然而,如果我们接受积极依恋的基本原理主要由初级过程的皮层下催产素回路介导,那么我们就可以更好地理解人类依恋的那些更高级的次级和三级过程心理方面。例如,最近的一项人类研究发现,鼻内催产素促进了一种幸灾乐祸(schadenfreude)的态度——这个德语术语可以大致翻译为”幸灾乐祸”(Shamay-Tsoory et al., 2009)。对于一个”爱”分子来说多么令人困惑。然而,也许这只是一种三级过程反应,反映了催产素产生的某种初级过程信心?
一旦我们吸收了催产素能效应对心智大脑(MindBrain)影响的初级过程经验,我们应该对催产素在精神病学中的应用有更好的想法。确定催产素和(安全的)阿片类药物如何有效对抗抑郁症将特别有趣,尤其是有时在分娩后不久发生的抑郁症类型。顺便说一句,已经知道”安全”的阿片类药物,如丁丙诺啡(buprenorphine),可以快速对抗许多未被其他药物帮助的人的抑郁症状(Bodkin et al., 1995)。
动物研究表明,催产素能减少分离焦虑,促进自信和积极的社交互动。它在人类身上是否产生类似效果?所有迹象都表明确实如此。近年来,人类催产素研究越来越多,主要是因为它可以通过鼻腔给药(鼻内给药)而不会产生任何不良副作用。过去几年关于鼻内催产素效果的许多有趣发现已被报道。催产素促进积极社交记忆和信任感的提取,同时降低社交互动中的焦虑和压力(Ishak等,2010)。最近,Ditzen及其同事(2009)给同居伴侣鼻内催产素,观察它如何调节他们在讨论可能引发冲突的各种话题时的积极和消极社交互动。鼻内催产素(40 IU)给药后,积极互动的比例上升(通过眼神接触、兴趣、情感自我表露、认可、关怀、非语言积极行为来测量),而消极互动(批评、蔑视、防御、支配、好斗、冷战、非语言消极行为、打断)减少。这种对比在男性中稍微更明显,因为男性在争论中可能比女性更好斗和强势。此外,双方的压力水平都下降了(通过血浆皮质醇测量),但女性的下降幅度大于男性。简而言之,催产素缓解了在可能出现冲突的情况下的紧张关系。
尽管我们不确切知道催产素为何具有这种平息效果,但我们可以基于以下知识进行推测:催产素能减少幼年动物的分离痛苦并促进自信。也许当关于分离的不安全感得到缓解时,人们会有更多友好的社交互动;他们不会防御或易怒。同样合理的是,当人类变得更加”自信”时,他们能够信任他人。实际上,我们甚至可以认为”自信”是描述促信任行为的最佳情感术语。这使人们能够更容易地与他人互动。最近对父亲的研究表明,在吸入几次催产素后,他们会更投入地与孩子玩耍(Naber等,2010)。在我们自己的研究中,我们发现催产素可以提高抑郁症患者通过观察他人眼睛来解读他人情绪的能力,而且他们的大脑在执行此类任务时显示出更高的唤醒(Pincus等,2010)。
总的来说,我们认为催产素在动物心智中产生的基本过程心理变化与在人类大脑中产生的变化相似。显然,人类和动物研究者应该明智地尝试找到共同的概念,使我们能够讨论所有物种的基本过程心理变化。这还没有发生。目前看来,催产素可能会加深动物和人类的积极社交参与,特别是当环境支持积极社交互动时。这可以改善社交生活质量和性感受的流动。
为此,需要对自己心智的专家观察者进行更细致的现象学研究。应该使用标准量表和自传式描述在许多情况下仔细研究不同个体,然后我们才能真正满意地掌握催产素如何影响人类情绪。简而言之,我们需要更多的神经精神分析研究,将现象学放在首位(Benedek & Rubenstein,1942;Panksepp,1999;Kaplan-Solms & Solms,2000),正如Francisco Varela(1999)等有远见者长期倡导的那样。
性欲(LUST)真的是一种情绪性情感吗?在前面的章节中,我们讨论了三方面的区分:(i)身体-稳态情感,如饥饿和口渴,(ii)感觉情感,如味觉的愉悦或良好的按摩,以及(iii)内在的基于大脑的情绪感受。我们注意到,稳态情感、感觉情感和情绪感受来自不同的大脑网络,与稳态和感觉情感不同,情绪情感总是伴随着情绪行为——在其他不像我们那样调节情绪的动物中有行动冲动和明确倾向。很容易看出,例如恐惧(FEAR)是一种情绪情感,因为它产生不可否认的强烈情绪感受,并且还产生诸如僵住和逃跑等行动(在恐惧系统的低激活和高激活下)。但性欲(LUST)不是一个如此明确的案例,因为稳态和感觉因素在性唤起中起着更大的作用。人们可能会怀疑性欲是稳态或感觉情感而不是情绪情感,或者它可能是这三者的组合。稳态解释似乎是合理的,因为身体/稳态激素释放在决定动物性准备状态方面起着重要作用。性欲直接而强烈地受到睾丸、卵巢分泌的性激素以及在较小程度上受肾上腺分泌的性激素的驱动,所有这些都受大脑底部”主腺体”垂体的控制。难道不能认为性欲产生稳态情感,更类似于饥饿和口渴,而不是来自恐惧和愤怒等大脑网络的”真正情绪”吗?
同样,感官体验,尤其是触觉和嗅觉,在性偏好和性唤起中发挥着至关重要的作用,这一点可能会促使我们认为欲望是一种感官性情感。在某些物种中,性渴望是由嗅觉信息素触发的,没有这些信息素,它们的LUST系统根本无法正常工作。在所有哺乳动物中,性行为在直接性刺激之前会通过诱发性的皮肤接触得到促进。通常,令人满意的性行为的前奏包括大量嬉戏性的求偶活动,以及通常以生殖器刺激为高潮的躯体感觉刺激。在许多物种中,特别是人类,存在大量的言语刺激、拥抱、亲吻、温柔的触摸和视觉愉悦。因此,LUST系统,就像所有其他社会情绪系统——CARE、PANIC/GRIEF和PLAY——一样,受到身体感觉所唤起的情感的强烈调节。LUST可能是与感官和稳态联系最紧密的情绪系统之一,比RAGE和FEAR更甚。
鉴于与稳态和感官考虑的如此紧密联系,为什么我们仍然希望将LUST视为一种基本的情绪?这恰恰说明了自然不尊重我们人为制定的分类。尽管如此,分类使我们能够在值得关注的复杂数据集中看到模式和关系。而LUST最自然地归属于”情绪”类别。我们坚持认为LUST是一个情绪-情感过程,因为在性活跃动物的全身求偶和交配活动中,行动准备状态是如此明显——而且因为情绪行动准备状态是LUST整体情感状态的一个基本特征。此外,外周(稳态)激素释放引起性准备状态的原因之一是它调节各种大脑化学物质,这些化学物质反过来调节大脑的欲望行动。因此,性激素不仅仅像口渴和饥饿那样具有局限的受体,它们的整体影响在于它们影响集中在动物和人类大脑皮层下区域的广泛欲望回路的方式。
对于感官方面也可以提出类似的整体性主张。嗅觉长期以来一直被认为是性准备状态和性渴望的主要信号,但其他感官也对整体情况有所贡献。例如,当雄性大鼠快速触摸雌性大鼠的臀部时,雌性大鼠以及许多其他物种的雌性会进入接受性的脊柱前凸姿势。这种触摸触发的”脊柱前凸反射”是雌性性准备状态的确切标志——这种持续的雌性身体姿势,背部弯曲、臀部上翘、尾巴偏向一侧,有利于雄性骑跨和进入。尽管关于感官体验如何唤起LUST的许多细节仍有待阐明(例如,从阴蒂到大脑更高部位的输入),但这些问题正受到越来越多的关注(Pfaff, 1999; Pfaus et al., 2003)。同样,在许多鸟类、灵长类动物,当然还有人类中,感官唤起的视觉方面也得到了充分认可。尽管如此,色情图像如此容易获得性唤起系统的途径,特别是在男性中,仍是一个谜。这是否是皮层视觉专门化直接接触原始皮层下情绪系统的一个主要例子?这仅仅是学习过程的结果,还是灵长类动物大脑固有的特性?我们只是不知道。但我们确实知道,没有皮层下的唤起,性感受会变得微弱。有大量的皮层下大脑部位可以产生性唤起和大脑刺激奖赏(Caggiula, 1970; MacLean & Ploog, 1962),突出了性愉悦的广泛位置。
然而,即使我们已经了解到性准备状态是由皮层下网络产生的,还有其他难题需要思考。这些大脑部位位于扩展的SEEKING系统的轨迹内,该系统产生各种各样的食欲行为(见[第三章])。沿着外侧下丘脑SEEKING系统轨迹的损伤会破坏性行为,实际上会破坏所有食欲行为。那么为什么我们不仅仅将求偶和交配等欲望行为视为SEEKING系统的一部分呢?性求偶与其他典型的SEEKING行为(如探索、觅食和跟踪)相比,有什么独特之处吗?
显然,专门用于性准备状态的内在LUST网络与SEEKING冲动紧密交织。但雄性交配行为与简单的SEEKING行为不同,具有专门的回路来促进骑跨、插入和射精。我们也知道有各种不同的化学物质以及共享的化学物质分别促进雌性和雄性的欲望,其中研究最充分的是催产素(oxytocin)和加压素(vasopressin)。由于性高潮在人类中是性活动中最强烈愉悦的方面,特别值得注意的是,在人类性高潮的大脑成像中,正是所有这些古老的跨下丘脑情绪系统,从腹侧中脑到爬行动物基底神经节,都被激活了(Georgiadis et al., 2006; Holstege et al., 2003)。尽管在人类性活动的食欲性触摸和爱抚阶段,许多高级大脑区域存在性别差异(Georgiadis et al., 2010),但性高潮在男性和女性中产生非常相似的图像——通常高级大脑活动减少,在介导性行为的相同大脑区域出现深刻的皮层下唤起——也许除了男性性高潮在中脑导水管周围灰质(PAG)中有更多唤起,因而更”原始”一些(Georgiadis et al., 2009)。
但在将欲望(LUST)的唤起(而非高潮性高潮)设想为一个简单的奖励诱导过程时,还存在另一个困境。施虐受虐(Sadomasochism)——通过施加痛苦来促进性唤起——长期以来一直被认为是人类性行为的一个特征。现在已经发现,即使是大鼠也会被轻微的压力源(如适度的疼痛)唤起性兴奋(Caggiula & Eibergen, 1969),这主要是由于压力唤起了外侧下丘脑奖励-寻求(SEEKING)系统(Everitt, 1990)。然而,在这里我们再次看到的只是动物应对某些促进广义寻求(SEEKING)冲动的压力源的另一种方式。轻微的足部电击可以激活大脑多巴胺释放。而轻微的压力源,如对大鼠尾部的压力,足以放大各种动机冲动(Antelman et al., 1975)。因此,这些效应可能基于与Valenstein及其同事(1970)首次描述”大脑奖励系统”内明显的动机可塑性时所暗示的相同大脑寻求(SEEKING)机制——所有这些都可以通过寻求(SEEKING)唤起能够参与各种积极和消极的情绪和动机唤起这一事实轻易解释。
在试图为情感性感受生成清晰的分类时,很难解决所有的模糊性,特别是哺乳动物性行为所涉及的那些,因为我们经常处理的是由人类产生的文化概念(三级过程,与思维相关的心智方面),而不是由进化创造的大脑功能。原始情感系统是祖先的宝藏,是让我们成为充满活力的生物的初级过程”工具”,这是本书的主要焦点。尽管如此,原始欲望(LUST)反映的是一种脑-身体状态,它位于我们选择称之为感觉性情感、稳态性情感和独特的大脑情感性情感的交汇处。此外,正如本章开头所强调的,正是在这个低层次的初级过程水平上,我们必须找到导致性别认同问题的一些生物学规则,或者也许我们应该说是议题(issues)——变性、同性恋和双性恋。既然我们已经讨论了皮层下欲望(LUST)回路的某些方面,我们现在回到那些棘手的认同(identity)议题,也许会让一些人感到惊讶的是,最可靠的知识正在从动物大脑研究中产生。
男性和女性的大脑系统在性冲动、满足感以及许多其他心理特征方面有所不同。正如我们所指出的,在男性和女性中产生这些特征的各种大脑系统是相当不同的。欲望(LUST)冲动以及两性之间的其他差异,不仅仅反映了外周的身体需求,而且决定性地反映了脑心智(BrainMind)的组织结构。哺乳动物性行为最有趣的方面之一是,性别的生理和心理表达具有不同但重叠的控制机制,这意味着身体的性别和心智的性别是相对独立发展的。此外,心智和身体相对独立的发展在子宫内就开始了——在发育中的哺乳动物胚胎中——远在生物体能够对性事有任何想法之前。
从生物学角度来说,我们将女性定义为天生具有XX染色体的个体,将男性定义为染色体为XY的个体。女性型大脑回路在大多数生物学女性中肯定更强,而男性型大脑回路在生物学男性中更强。然而,在本节中,我们将探讨性行为的这两个方面如何并不总是以我们预期的方式相匹配——当心智/身体性别/性(genders/sexes)混合时,会产生男性身体中的女性大脑或女性身体中的男性大脑。
当我们将这些复杂的生物学现象与同样复杂的个人、社会和文化现象——个体认同(identity)结合起来时——这一现象包括性取向(sexual orientation)和性别认同(gender identification)——我们可以确信事情会变得无限复杂,在从生物学到文化、从初级到三级的心智大脑(MindBrain)组织层面都存在交叉影响。正如我们之前提到的,我们不会试图讨论社会期望和性别角色的议题,因为这些确实是无法通过动物模型理解的三级过程创造物。相反,初级过程的生物学复杂性在其他动物身上比在人类身上能够得到更好的研究。我们认为这项生物学研究对理解我们的情欲感受具有深远的意义。
在开始之前,明确定义我们将使用的标签是很重要的。在这里,同性恋(homosexuality)意味着渴望以各种形式与外表上属于同一生物学性别的成员建立情欲、性关系。这并不一定意味着拥有男性典型身体的人在心智中感觉像女人,或拥有女性典型身体的人在心智中感觉像男人。同性恋在初级过程的心智大脑(MindBrain)层面可能有也可能没有(在某些或所有情况下)生物学基础。我们根本没有简单的方法来了解。我们无法像在动物模型中那样轻易地窥视人类大脑的性别分化,而在动物模型中,潜在的生物学议题可以得到非常详细的阐明。
相比之下,跨性别(transgender)一词是一个总括性术语,通常用于描述一个人在男性或女性身份方面的心理感受明显与其身体外观和/或染色体生物学特征不匹配的情况。当这种”性别认同”差异如此强烈,以至于个体考虑通过手术改变身体形态以匹配其关于自我身份的心理、情感和心理感受时,变性(transsexual)一词更为常用。缓慢积累的数据往往表明,跨性别个体的大脑在性欲组织方式上确实与大多数其他同性别身体的人不同(Gooren, 2006)。这些个体如此频繁地不得不与社会偏见作斗争才能过上满意的生活,这是一个持续的文化悲剧。
动物研究表明,出生时大脑心智中性别认同的潜在印记是不可见的,而身体的性别通常是明确的。然而,大脑的性别对决定一个人的性别认同具有决定性作用。男性身体中的女性大脑会导致一个人感觉自己是女性却处在男性身体中,而女性身体中的男性大脑会产生一个感觉自己是男性却处在女性身体中的人。这是跨性别状态的基本科学定义。显然,大脑的生物学特性对一个人的性别认同和命运有着深远的影响。我们可以确信,那些坚持说自己内心有个女孩想要出来的小男孩,以及那些感觉自己的思维更像男孩的小女孩,往往反映了一种生物学智慧,父母应该以勇气和敏感性倾听,而不是否认。
对几种非人类哺乳动物胎儿成熟过程的深入研究已经阐明,大脑和身体的性发育沿着不同的路径进行。这些研究强调了所有哺乳动物性别发育的一般原则,而不仅仅是某一特定物种的差异。现在已知这一知识与我们自己物种的同源过程高度相关,至少在少数几个地方,我们关于动物大脑中跨性别因素的知识已经在人类物种中进行了艰难的概念转化(Zhou et al., 1995)。简而言之,在产前发育期间决定大脑性别特异性组织的性激素与那些帮助指定男性和女性生殖器官外观的性激素是不同的。
如果这些胎儿大脑化学物质以非典型方式展开,发育中的有机体的大脑性别认同可能在初级过程层面发生转变。这可能由于多种原因发生,包括母亲在怀孕期间可能经历的极端心理压力,以及激素医疗治疗,甚至暴露于环境中的同系物(例如,我们污染的环境中有许多像性激素一样作用的化学物质,这有助于解释为什么一些两栖动物的生殖能力受到损害)。当这些压力源在怀孕的关键阶段占主导地位时,婴儿可能真的会出生时拥有女性身体中的男性典型大脑或男性身体中的女性典型大脑,以及”极端”之间的大量渐变。这种多样性可以用来支持关于双性恋的一种非常自由的论点:如果大脑和身体性别的渐变是常见的,那么极端情况,包括严格的异性恋、严格的同性恋或性别认同的极端,似乎更可能是例外而不是规则。我们怀疑数据支持大多数个体属于大脑和身体性别匹配的类别,但目前还没有明确的经验方法来做出这样的判断。
无论如何,胎儿大脑具有可能与其身体性器官不匹配的性别认同这一事实表明,性别认同不仅仅是在成熟过程中学习的。同时,显然所有自我认同问题都部分地受到学习和文化的塑造。但在其原始形式中,男性型和女性型大脑包含重要的先天差异。它们不一定与身体视觉上明显的男性化和女性化相匹配。
当一个孩子出生时拥有一种性别的身体和另一种性别的大脑时,社会压力只会使事情变得更糟,因为孩子的先天性别认同无法通过说服来改变。令人遗憾的是,一些自认为文明的国家尚未为跨性别人士提供平等的权利和在世界上获得幸福的机会。希望教育最终会改变那些歧视自然(这是斯宾诺莎认为上帝所是的东西)的人的心。这些神经生物学事实与许多美洲印第安部落传统上遵循的社会实践相一致:有时,自然注定女性性别认同应该在生物学男性的大脑中绽放,男性气质应该在生物学女性中蓬勃发展(Zhou et al., 1995)。我们一些祖先的智慧欣然接受了自然赋予脊椎动物的心理性别多样性——男性化和女性化的连续体——而我们文化中的许多人已经学会了蔑视这一点。但这种变异在其他动物中也很丰富,其基本原则非常相似(例如,Bagemihl, 1999; Gavrilets & Rice, 2006)。
妊娠第二孕期的发育过程似乎对建立人类大脑的性别认同至关重要(Murray et al., 2000)。这在所有物种中并不相同。在大鼠中,第三孕期(实际上在妊娠第19天达到最高峰)是确定大脑性别的关键时期。这些产前的性别印记对控制性冲动、母性冲动和其他社会倾向(如攻击性)的广泛大脑回路和神经化学物质模式具有重要影响,更不用说一系列更具争议的高级认知优势和劣势,如同理心和嫉妒。这就是为什么当我们讨论性腺激素在早期发育期间在大脑上留下的印记时,区分生物学”性别”(sex)和心理生物学”性别认同”(gender)的概念很重要。
在男孩中,这些差异中的大多数(但不是全部)源于Y染色体对发育中的男性所做的唯一主要事情。它促进(通过睾丸决定因子的分泌)具有在妊娠期间分泌睾酮能力的睾丸生长。在具有Y染色体的胎儿中,睾酮在出生前分泌(在人类妊娠的第二孕期出现惊人的激增;在大鼠中,这发生在第三孕期)。这种睾酮启动了大脑和身体的雄性化(Berta et al., 1990)。如果身体和脑心智组织都由相同的生物化学过程控制,事情就不会如此复杂,但实际上,它们各自走不同的路径。当睾酮被一种叫做芳香化酶(aromatase)的酶转化为雌激素(estrogen)时,大脑被雄性化(酶是可以启动和促进化学反应的化学物质,但它们不存在于最终的化学产物中)。相比之下,当睾酮被5-α还原酶(5-alpha-reductase)转化为双氢睾酮(DHT)时,身体被雄性化(Breedlove, 1992)。
因此,雌激素的早期激增(在流行观点中,它与女性性功能相关)对于雄性化胎儿大脑至关重要。如果怀有女性后代的人类母亲在妊娠的这个关键点被注射大量雌激素(就像在许多研究中对大鼠母亲所做的那样),身体上为女性的后代很可能会出生时具有男性典型态度。同样,如果男性胎儿自身的睾酮不能被转化为雌激素,他将会出生时具有女性化的大脑。因此,胚胎体内的睾酮到雌激素级联反应决定了其大脑的男性或女性特征。其中许多事实已在实验动物中得到很好的研究,包括某些环境压力源如何改变这些过程。但只有间接的(尽管迅速增加的)证据表明这些经验也适用于人类。
胎儿男性身体沿着不同的化学途径发育。在胎儿发育中,所有哺乳动物胚胎的身体最初在外观上都是女性。胚胎的身体通过睾酮转化为DHT而雄性化。如果不是因为男孩体内胎儿睾酮的激增,所有人类在身体上都会表现为女性。
因此,我们的遗传和妊娠经历包含形成我们心智和身体性别认同的潜力。然而,我们心智的个体性别特征在出生时并不明显。所有这些心理生物学调性都必须通过在世界上生活来演奏。家庭和文化最终将提供许多老师,但最早的是构建通往男性和女性的心理和生理路径的性染色体。
如上所述,当胎儿雄性大鼠正常发育时,在妊娠第19天会出现睾酮激增。芳香化酶将大部分睾酮转化为雌激素,这促进了男性大脑发育。同时,5-α还原酶将睾酮转化为DHT,这产生了雄性身体。然而,产前母体压力与遗传雄性胎儿中女性大脑的发育有关。如果母鼠在那个关键时期经历压力,睾酮释放可能会来得太早,在芳香化酶准备好将其转化为雌激素之前。因此,大脑中的雄性性欲(LUST)回路不能正常发育。然而,这种早期睾酮释放的大部分确实被转化为DHT,从而产生男性典型身体。对于人类,我们根本不知道这种转化的参数,因此可以想象,规范的人类发育在大多数时候更多地沿着更广泛的连续体进行。
无论如何,出生于高度紧张母亲的雄性大鼠具有正常的雄性身体,但当它们在青春期性成熟时,会表现出较少的雄性行为和更多的雌性行为。这表明它们的大脑没有完全雄性化。在正常的大鼠窝中(通常从几只到十几只幼鼠不等),80%的雄性大鼠将是”种鼠”,意味着它们在成年后将只表现出雄性性冲动,而20%将是”废鼠”,意味着它们将相对无性。当母鼠承受压力时,导致妊娠期间雌激素制造过程中断,只有20%的雄性将成为种鼠,约20%成为废鼠,而其余约60%将表现出双性恋和同性恋行为倾向(Ward, 1992)。同样,我们无法确定这如何适用于人类:很容易想象,由于妊娠期更长、大脑更大,以及发育在具有更大变化窗口的化学梯度背景下进行,人类可能比大鼠模型中相当简单直接的结果具有更大的变异性。尽管有这些警告,物种之间细节上的差异必然是巨大的。很明显,上述一般原则适用于哺乳动物物种。
胚胎化学可能无法解释所有同性恋甚至跨性别倾向,但它肯定解释了一些,也许是大多数跨性别倾向。但在人口层面的证据绝不是很多。例如,有记录表明,在二战最艰难的年份出生的德国男孩中,同性恋水平更高,当时孕妇承受了大量压力。但这是否是因为上述变量?我们不知道。但可以合理地假设,这些母亲在妊娠关键的第二个三月期(trimester)经历了过度压力,可能导致拥有雄性身体类型但雌性大脑倾向的婴儿数量超过正常水平。但这其中很多是推断。
现在让我们考虑相反的情况,当雌激素充足但DHT在胎儿生命的第二个三月期供应不足时。这种情况是由基因异常引起的,在多米尼加共和国生活的一小群人中得到了生动的描述。该群体中的雄性在基因上缺乏5-α-还原酶(5-alpha-reductase),这种酶有助于促进外部雄性身体特征(阴囊和阴茎)的发育。因为他们不缺乏芳香化酶(aromatase),男性的大脑将沿着典型的雄性路线发育。然而,由于缺乏DHT,这些男孩的身体在出生时呈现女性外观,没有睾丸(睾丸未下降留在腹腔内),阴茎呈雏形状态,可能被误认为是增大的阴蒂。因为这些生物学男孩看起来像女孩,而且没有办法看到他们心智大脑(MindBrain)的组织结构,所以他们被当作女孩抚养。
尽管如此,他们的Y染色体赋予在青春期变得活跃并产生睾酮,导致体毛增加、声音变低沉、阴茎增大,最后睾丸下降。由于这种在青春期的戏剧性转变,这些少年被称为guevedoces——字面意思是”12岁时的阴茎”(Marks, 2004)。典型的雄性性冲动也开始出现。因此,这些男孩的青春期性欲开始指向女性,尽管他们整个童年都被当作女孩抚养。这可能表明雄性大脑本能地准备好对人类女性的某些特征做出反应,比如面部和身体特征、声音语调等等。
值得注意的是,作为女孩抚养的guevedoces男孩能够轻松地承担典型的雄性角色。这部分是因为这种现象是众所周知的,社会支持已经到位。在我们自己社会的类似情况中并非如此。一个著名案例突显了这些问题,其中”系统”试图强迫雄性大脑承担女性身份:一个生物学男孩在出生后不久就被手术转换为女性,因为包皮环切手术失败(Diamond, 2004; Money, 1995)。小约翰尼变成了琼,并在所有家庭和文化期望”他”是”她”的情况下长大;但约翰尼/琼的情况并没有像典型的guevedoces那样顺利。约翰尼的医生认为性别认同是由文化决定的,他们坚持认为,努力将他作为女孩抚养将成功使约翰尼相信他是女孩。然而,约翰尼总是继续相信他是男孩,当他成年时,他坚持要求将他的身体恢复到与他生物学上预先倾向的雄性心理相对应的状态。还有其他类似的案例没有涉及任何手术失败;人们只是坚持认为他们身体的性特征和心智的性别特征不匹配。在许多其他可能导致这种”混淆”的原因中,有一种称为雄激素不敏感综合征(androgen insensitivity syndrome)的病症;在其最极端的形式中,这些雄性拥有女性生殖器以及未下降的睾丸。
从基因角度来说是女性的胎儿同样容易受到性别不一致的影响。如果基因为女性的胎儿在发育敏感期暴露于过多的雌激素,大脑将呈现出类似男性的特征,而身体仍保持女性化。这些女性在成熟后会优先表现出典型的男性行为(Gorski, 1988)。事实上,在20世纪40年代和50年代,女孩的假小子气质inadvertently被注射己烯雌酚(DES)所促进,这是一种曾用于预防流产的雌激素生成激素,特别是当母亲在怀孕第二孕期暴露于这种激素时(Ehrhardt et al., 1985)。这些性别异常通常不会因母亲自身的大量雌激素水平而发生,因为染色体XX配置会告知女性胎儿制造蛋白质(例如甲胎蛋白),以在发育早期阻止跨性别的化学影响。然而,当注射过量雌激素时,它们可以淹没这个保护系统,该系统不再能够”清除”所有那些”男性大脑汁液”。同样,压力和环境因素可能导致在女性身体中产生男性大脑。
不过,我们应该再次指出,这很可能发生在人类正常发育的广泛变异梯度上,完全无法确定任何单个个体是否偏离了典型结果。
从某种意义上说,我们在本章中一直在薄冰上滑行。人们对性倾向有强烈的感受。同性恋者和跨性别者在许多文化中处境艰难。他们经常被剥夺法律下的平等权利。在许多国家,他们不被允许与外表上看起来是同性的人结婚。他们受到歧视——这一切都发生在人类生活的第三层过程层面,这些层面通常超出了本书的关注重点。然而,这些第三层过程的文化现象是我们作为尊重各层面人类差异的人类所极为关心的。作为科学家,我们必须坚持让那些不理解或不尊重这种人类差异的人从无知、恐惧和仇恨的文化阴影中走出来,进入科学现实的阳光下。我们的文化生活充满了无知导致痛苦的人类故事。因此,也许我们这些有幸接受科学教育的人应该设法将一些光明照进现代文化许多角落中仍然笼罩着这个话题的阴影中。
想想”比利”·蒂普顿,他的人生故事在《“适合我”:比利·蒂普顿的双重生活》(Middlebrook, 1998)中有所描述。1914年出生时名为多萝西·露西尔·蒂普顿,她在十几岁时就成为一名出色的爵士音乐家。当她决定从事音乐职业时,她打扮成男人,显然是为了在专业圈子里更容易被接受。当不表演时,她保持女性身份。她在一段女同性恋关系中度过了数年,并在人生的某个时刻开始一直打扮成男人。随后又发生了几段与女性的关系。她通过束胸来伪装成男人。在性活动中,她不喜欢被触碰,可能使用了假阴茎。虽然她从未与任何恋人结婚,但比利的一位伴侣透露”他”是”我生命中最美妙的爱情”。比利和另一位伴侣收养了三个男孩,他们都视”他”为父亲,在”他”去世后发现”他”的真实性别身份时感到震惊。
我们对比利母亲在怀孕第二孕期的心理状态一无所知。作为胎儿,比利是否可能暴露于过量雌激素,从而在女性身体内编程了男性类型的大脑?我们关于大脑性别分化的知识诱使我们接受这样的间接证据:比利的大脑确实被男性化了,尽管他/她拥有女性的身体。然而,也许比利无法忍受生活在一个聪明女性无法像任何具有同等才华和思想的男性那样获得同等机会来表达她所有美好艺术激情和技能的时代。我们根本不知道。这些是永远无法追溯回答的问题。这就是我们在本书大部分内容中面临的问题。我们需要的那种基础知识,以真正理解哺乳动物进化中的心智要素,仅仅通过研究人类行为的无限复杂性是永远无法理解的。
胎儿发育早期的迷人细节告诉我们一个深刻的自然事实:胎儿发育过程中存在四种极端的性别结果(当然,中间还有更多温和的变化)。胎儿可以是典型的女性,拥有女性的身体和心智,或者可以是典型的男性,拥有男性的身体和心智。拥有女性身体和男性情感心智的婴儿,或者拥有男性身体和女性情感心智的婴儿确实更为罕见,但已经被充分理解,可以将它们视为正常的性别发育模式。在青春期,在睾丸和卵巢大量分泌性激素的影响下,早期的男性和女性印记开始活跃起来。这个性成熟的激活期(activational period)延续了胎儿发育时期的前意识大脑印记。尽管强烈经历的童年文化影响也在性别发育中发挥作用,但青春期激活了胎儿遗留的印记。就像一个古老的”顽皮乐团”,它开始在大脑深处充满欲望的隐秘区域演奏坚持不懈的生物乐曲。理解这一点,并将其接受为命运,就是拥有充分的智慧和宽容。不将这些变化视为”正常”,就是坚持认为文化规范比永恒的生物多样性更重要。
尽管所有哺乳动物在男性和女性性行为方面共享相当相似的原始过程大脑机制,但它们在生态环境和社会群体细节中表达这些冲动的方式却有巨大差异。即使在密切相关的物种之间,生殖策略也各不相同(Carter et al., 1995)。例如,长臂猿终生只与一个伴侣交配,但大猩猩更喜欢后宫型家庭结构,许多其他灵长类动物也是如此,比如白鼻长尾猴(Ceropithecus nictitans),其社会群体通常由一只雄性和最多九只雌性及其后代组成。猩猩倾向于独居,两性主要为了交配目的而聚在一起,而黑猩猩则非常社会化且滥交,相当随意地共享伴侣。因此,即使是我们最亲近的进化近亲,大型类人猿,也无法清楚地揭示我们内在的性本质。也许因为我们丰富的想象力和创造多样文化的能力,我们可能在人类中看到所有这些生殖策略的变体。
我们对支撑人类大脑中这些哺乳动物性行为变化的神经科学原因和相关性仍然基本无知,我们对支持大多数正常人类性行为变体的大脑活动同样不了解。例如,人类性活动和性唤起存在许多个体差异,这可能反映也可能不反映性化学的差异。人类男性的持续性交导致单次强烈高潮往往会带来长时间的满足感,导致一段时间的性不活跃(不应期(refractory period)),可能是由于大脑化学物质的消耗。其他哺乳动物物种的雄性,特别是实验室啮齿动物,需要多次高潮才能完全满足,也许是因为它们的性快感化学物质在大脑中更逐步地消耗。然而,人类女性也比男性更能够连续多次高潮,可能也是因为她们的情感化学物质消耗得不那么快。这可能意味着什么?也许我们人类是从滥交物种进化而来的,在那里女性生殖由多个性伴侣促进,就像黑猩猩中常见的那样。换句话说,也许女性多次高潮的能力可能反映了一种祖先能力,即被几个伴侣反复唤起性欲。当然,这是一个”就这样”的进化故事,就像进化心理学中的许多性感想法一样,因为大多数这样的假设无法通过严格的研究策略来检验。
我们根本无法确定我们自己的大脑在妊娠期间被男性化(去女性化)或女性化的程度。有大量数据表明,我们食指和无名指长度的不对称性(正式称为2D:4D比率,D代表手指digit)可能用于估计这些问题,但这远未确定。无论如何,食指(2D)比无名指(4D)短的程度可能反映了一个人在胎儿时期的男性化程度。关于这种手指长度测量的性别和性别差异有大量数据,这在19世纪首次被描述。要探索这些迷人的发现,只需用谷歌搜索”无名指和食指长度”或”2D:4D比率”,就可以思考这个迷人生物标志物的许多维度。这个比率的正常差异在男性中比女性更普遍,男性化程度也是性别认同的函数。据报道,这一测量在男同性恋者中往往女性化,在女同性恋者中男性化,但我们远未确信这可以作为大脑或身体男性化的选择性测量,或两者兼有,或两者都不是。验证该测量的意义很难获得。该测量还随地理-国籍、性格和除性行为之外的许多其他变量而变化(Manning, 2002)。此外,尽管它表面上简单明了,但存在大量测量问题,专家之间存在相当大的差异(Voracek et al., 2007)。尽管如此,如果能证明它是子宫内激素发生情况的有效生物标志物,那将是非常了不起的。
一般来说,我们应该对基于理论视角的问题过早定论保持警惕,尤其是那些难以用神经生物学术语检验的理论。这对于经典的性心理发展理论尤为突出,例如西格蒙德·弗洛伊德最初提出的那些理论。作为教学练习,我们通过解构弗洛伊德曾经颇具影响力的关于我们性本质的观点来结束本章,这些观点现在需要被视为创造性的理论推测,而非基于科学的专家意见。
在前面的章节中,我们指出神经科学提供的情绪分类法远比精神分析和许多其他精神病学理论所提供的基本情感清单更完整、更科学。当我们考虑到弗洛伊德提出只有两种驱力:力比多(libido)和破坏性时,这一事实尤为明显。在如此有限的范围内,弗洛伊德试图用他所谓的”组成本能”——成人生殖性欲的力比多前驱——来理解婴儿期和儿童期看似非性的方面。弗洛伊德(1905b/1968)提出了四种组成本能:口欲、肛欲、阳具欲和生殖欲。(他说的是阳具期而非生殖期,因为他有名——或臭名昭著——地认为,在童年的这个阶段——大约3-5岁时,阴茎是两性关注的焦点)。但”阴茎嫉妒”在女性心理发展中真的比年轻男孩的”乳房崇拜”更有影响力吗?我们对此表示怀疑。
组成本能理论基于三个考虑因素。第一个是对婴儿和儿童的观察。弗洛伊德观察到,口腔、肛门和阳具活动(后者包括手淫)在童年时期提供快感。由于他相信只存在两种驱力,这些快感被认为是力比多性质的。因此,当婴儿在吮吸或非营养性含嘴时表现出明显的快感,弗洛伊德认为这些是婴儿期的力比多快感。他注意到幼儿在排便时感到快感;因此,他将肛欲驱力视为一种力比多快感。阳具性欲大约在3-4岁时出现,此时小男孩发现了他的阴茎,小女孩发现了她的阴蒂,这显然是性行为。俄狄浦斯情结(Oedipus complex)与生殖期重叠,发生在儿童对异性父母产生性/浪漫依恋时。他对儿童和成人的精神分析调查使他确信,俄狄浦斯情结是性发展的正常阶段。
弗洛伊德相信组成本能存在的第二个原因是,他认为它们被整合到成人性行为中。例如,接吻是成人在性行为中参与的一种口腔活动。还有类似于童年阳具手淫的生殖器爱抚。弗洛伊德提出组成本能的第三个原因是,他认为它们在性变态(perversion)案例中占主导地位。弗洛伊德生活在维多利亚时代的维也纳社会,当时性变态被认为包括许多我们今天认为正常的性行为。例如,口交被认为是变态的,所有形式的同性恋也是如此。弗洛伊德认为,当力比多发展停滞时,组成本能的不成熟形式以构成变态的方式主导了成人的性生活。因此,他得出结论,性变态者在力比多发展的早期阶段性”固着”(弗洛伊德,1905b/1968)。
弗洛伊德的组成本能观点只有在人们接受只有两种驱力的前提下才说得通。然而,现代神经科学已经证明,快感也可以从非性的情绪系统中获得,例如恐慌/悲伤(PANIC/GRIEF)系统的积极面、或从玩耍(PLAY)、照料(CARE)或寻求(SEEKING)系统中获得。因此,说所有婴儿期的快感从根本上都是力比多性质的,这是不可信的。例如,当我们试图为口欲的力比多性质辩护时,我们就站在了非常不稳固的立场上。吮吸和非营养性含嘴可能不是力比多快感的表达——至少不完全是。相反,它们可能表达婴儿在靠近母亲时感受到的非性快感(恐慌/悲伤系统的积极面)。或者,这些活动可能反映了与进食相关的愉快的体内平衡情感。人们也可以想象,在进食之前,婴儿的寻求系统被愉快地唤起,而含嘴是这种愉快期待的一部分。再者,含嘴有时可能是一种玩耍形式——婴儿尽管运动能力相对有限,也能参与其中。
我们也有理由质疑弗洛伊德关于性器期性欲在3-4岁出现的时间表。后续研究表明,即使是婴儿,剧烈的摇晃也可能诱发性高潮(Kinsey et al., 1948; Martinson, 1994; Yates, 1978),而其他研究观察到婴儿早在男孩6个月、女孩10个月时就会持续触摸生殖器(Galenson & Roiphe, 1974)。当然,婴儿无法告诉研究人员他们是否经历过性高潮,人们也无法确定不太集中的生殖器触摸是否具有性刺激。毕竟,婴儿也会以类似的方式玩弄他们的耳朵(Levine, 1951)。还需要考虑的是,生命最初18个月内的一定程度的生殖器游戏是一个积极信号,因为它与良好的母婴关系和普遍的满足感相关。被忽视的婴儿不会触摸自己(Spitz & Wolf, 1946)。因此,如果婴儿感到满足,他们触摸自己是为了体验短暂愉悦的性感觉;这可能会增加满足感,但并不能将其视为成熟性欲(LUST)更典型的集中性活动。
现代神经科学告诉我们许多关于成人性行为的原理,也告诉我们许多关于性脑胚胎发育的知识,但它很少告诉我们关于青春期前的性发育——关于儿童性发展的信息。因此,我们无法从婴儿期和儿童期明显的力比多行为观察中得出确切结论。我们甚至不知道弗洛伊德提出的婴儿发展围绕口腔期、肛门期和性器期变化的观点是否正确。然而,即使口腔性、肛门性和生殖器性是儿童期的核心问题,也没有理由相信它们本质上都是纯粹力比多性质的。我们知道哺乳动物的情绪分类至少包含七种不同的基本情绪,其中四种(分离痛苦/悲伤PANIC/GRIEF的积极面、玩耍PLAY、照料CARE、探寻SEEK)产生的积极情感是非性的,因此不太可能所有婴儿期(或成人)的愉悦都像弗洛伊德所暗示的那样从根本上是力比多性质的。
因此,当我们重新考虑经典的性心理发展理论时,现代神经科学给我们提供了思考的素材。然而,它并没有告诉我们关于儿童期性心理发展的真实本质,或关于文化驱动的三级加工层面的脑心(BrainMind)涌现的任何方面。这使得心理治疗师处于一个不理想的境地,有许多未解答的问题——特别是关于性发展的问题。例如,同性恋是否涉及异性典型脑区和化学物质的激活?同性恋女孩的前下丘脑是否有更密集的细胞群,她们的大脑是否充满更多睾酮和加压素(vasopressin)?同性恋男孩是否处于相应的位置,腹内侧下丘脑有更高的敏感性和更多的催产素活动?我们对这些关于人类的问题没有答案。我们在其他几种动物物种中有答案,特别是实验室大鼠和小鼠。但是,当然,人类不是大鼠、海象或猴子。那么,我们应该多大程度上相信从被错误称为”低等物种”中得出的一般原理呢?
关于儿童期的性病理,我们同样无知。儿童期的性过度刺激是否会导致性脑区和化学物质的过早激活?为什么被父母忽视的儿童不愿触摸他们的生殖器?他们的性脑化学物质是否处于低水平,如果是,是什么导致它们减少?这些只是关于人类性发展和所有其他情绪如何涌现到三级加工思维和文化意识中的众多问题中的几个,这些问题仍有待充分回答。神经科学与精神分析的性心理发展理论之间仍然存在鸿沟,更不用说人类文化生活了。现代神经科学给我们提供了充分的理由来质疑关于本能成分的经典精神分析理论,但它还没有提供证据,也可能根本无法提供证据,来填补我们对祖先生存工具的知识与展现出无尽人类存在多样性的个体生活之间的巨大鸿沟。这凸显了神经科学分析需要与更复杂的心理学和社会学分析共同合作,才能获得对人类复杂性的更全面理解。
在本章中,我们讨论了现代神经科学为我们理解哺乳动物性行为提供的一些见解。我们已经看到,雄性和雌性大脑中的性回路和性化学物质是不同的。在雄性中,前下丘脑是性行为的焦点,睾酮介导加压素的产生,这解释了雄性性行为的大部分内容。在雌性中,腹内侧下丘脑是初级加工性控制中枢的一部分,主要的性化学物质是雌激素和孕酮。这些激素反过来介导催产素的活动,催产素是一种显著调控雌性性反应的神经调节剂。尽管催产素被普遍宣传为”爱的激素”,但我们认为这是对其作用的简化,因为它可能不会直接产生太多积极情感。相反,催产素可能增强内源性阿片类物质的活动,而正是这些物质产生了催产素给药引发的大部分积极情感。尽管如此,很明显催产素确实促进积极社交情感的产生,特别是对胜任母职很重要的信心和信任。我们预期阿片类物质也有同样作用,而且在高剂量时甚至会导致人们不需要其他人,从而导致成瘾者的社会孤立。
在哺乳动物中,性行为在初级加工层面是LUST回路的产物。但LUST是真正的情绪情感,还是更适合归类为体内平衡或感觉情感?我们认为它应该被视为情绪情感,因为它直接产生复杂的本能性行为,以及相关的原始情感,从纯粹的性欲到性高潮。这种心理行为行动倾向是初级加工情绪的标志。在许多物种的雌性中,性唤起促进性接受姿势的脊柱前凸反射,在雄性中则产生相应的求偶、爬跨、插入(成功的抽插)和射精等性行为。
性行为进一步复杂化(尤其在人类的三级加工层面),因为性身体和性大脑在子宫内沿着不同的轨迹发育。雄性大脑是在睾酮转化为雌激素时形成的,雄性身体是在睾酮转化为DHT时形成的。所有胎儿身体最初都是雌性,如果没有干扰,雌性身体将继续发育。然而,如果胎儿在妊娠中期的关键时期暴露于过多雌激素,雌性大脑和心智可能会被雄性化。
性行为这一主题,如同所有其他情绪一样,充满了未解之谜。是什么决定了密切相关的灵长类物种繁殖策略的多样性?是什么决定了特定物种内的不同策略?环境影响如何决定人类的性表达?这些差异的化学和神经相关因素可能是什么?对每位心理治疗师来说,一个关键问题是性驱力——LUST——在童年期间如何发展?人们可以提出许多类似的问题,所有这些问题都有待未来研究者去阐明。这是科学最美妙的一面。工作永远不会完成。神经科学确实如此。可以无止境地收获更精确的知识。
曾在我体内游弋的小眼睛,
在我声音中欢欣的小耳朵,
完美的脸颊、下巴和手指
以及带着早期话语的嘴唇
如此珍贵而湿润, . . .
那些永远逝去的日子里的孩子,
你尽情地笑和哭,
奔跑并四处攀爬
但总是绕回我身边,
因为我是你在地球上的第一个家. . . .
—安妮萨·米勒,《宝贝的爱》(1995)
哺乳动物之所以能够存在于地球上,是因为它们的大脑和身体准备好投入大量时间和精力来照顾后代,而后代如果没有这种奉献就根本无法生存。母性关注的投入并非偶然:它建立在一套坚实的本能大脑冲动之上,用以养育新生儿并与其建立联结。人类母性的奇迹,包括大家庭在养育中的作用,长期以来一直被人类学家所赞颂(Hrdy, 2009; Konner, 2010)。当然,在现代人类中,近亲和大家庭并不总是个人直接社区的一部分,这种原始动机常常必须在缺乏实际和情感支持的环境中追求——导致这些最基本和最强大的本能行动倾向与各种担忧和不安全感混杂在一起,可能削弱母性使命。在这种情况下,养育的喜悦可能被负面情绪的主导所掩盖。
然而,当一个人被支持性他人包围时,母性的情感交响曲必须被视为大自然的伟大馈赠之一。我们所有人都可能感受到这份馈赠在我们内心的印记,即使是那些永远不会生育的人。许多物种的父亲在大脑中都有潜在的母性回路,等待合适的环境来放大其潜力(de Jong et al., 2009)。因此,人们很容易论证,人类共情的根源深深扎根于所有哺乳动物产生关爱感的古老回路中,我们在其中将自己的幸福与他人的幸福联系起来(Decety & Ickes, 2009; Hein & Singer, 2008; Iacoboni, 2009a, 2009b)。他人的PANIC/GRIEF感受(见下一章)可能是促进共情奉献的最强大情绪共鸣之一。
在某些物种中,CARE冲动如此强大,以至于很容易延伸到其他物种的幼崽。目前,人们可以在互联网上找到许多哺乳动物跨物种母性奉献的非凡例子的摄影记录。在某些物种中,如实验室大鼠,母亲不会与自己的幼崽形成特定的联结,人们可以随意在不同窝之间交叉寄养依赖性幼崽。然而,在其他物种中,特别是那些可以说是’一出生就能站立’的有蹄类动物,母亲在出生后数小时内与后代形成专属的社会联结,在此时间框架之后,它们通常不会接受其他幼崽。然而,正如我们将看到的,对联结的神经和情绪机制的理解可以让我们”重新打开”联结窗口,并通过物理、药理和社会干预在这些物种中促进母性联结的建立。
在本章以及接下来聚焦于PANIC/GRIEF(恐慌/悲伤)和PLAY(玩耍)的章节中,我们将探讨产生非性社会联结的三个大脑系统。本章将重点讨论CARE系统(照料系统),它以母性奉献为典型代表。我们将讨论关于哺乳动物大脑如何产生养育冲动的新兴知识;然后我们将简要讨论这些化学物质如何控制社会学习和更高级的社会认知。在下一章中,我们将从另一面,即从婴儿的角度来考察:幼小动物如何与父母形成情感联结。然后我们将转向年长的动物,以及幼年和成年动物形成积极的非性联结或友谊的方式。我们最初将促进婴儿与母亲联结的原始情绪称为PANIC系统(恐慌系统)。使用这个不寻常的标签是为了强调一个事实:当大多数幼年哺乳动物和一些鸟类与照顾者分离时,产生的感觉可能类似于”恐慌发作”——这是一种与普通恐惧和焦虑截然不同的精神病发作(见第5章)。我们仍然认为这是描述”分离痛苦”系统所产生的原始情感的好标签。然而,由于这个标签引起了惊愕和困惑,我们在这里使用PANIC/GRIEF——或简称GRIEF(悲伤)。由于这种重新标记,一些论点可能更容易理解。例如,在年长的动物和人类中,随着情感联结的牢固建立,社会丧失可以激活更全面的痛苦情感,这些情感更容易用悲伤和哀痛来描述。在讨论完GRIEF系统之后,我们将用一章讨论PLAY系统,它促使人和动物,尤其是幼小的个体,参与以粗暴打闹游戏活动为代表的快乐竞争性互动。这种美妙的情绪能量使幼儿能够快速成为朋友,也使年长者能够做到这一点,只是更加渐进。
关于CARE系统,以及GRIEF和PLAY系统,仍有许多研究有待完成。这些对于理解社会依恋和可能促进抑郁的联结失败如此重要的初级过程情绪系统,在神经科学或生物精神病学中尚未得到广泛认可。然而,我们现在对这些非性社会系统已经有了足够的了解,可以在大脑中识别它们,理解它们运作的一些重要方式,并设想它们在人类心理健康和情绪障碍中可能扮演的角色。
我们也理解这些系统具有密切的相互关系。例如,CARE抑制GRIEF,而GRIEF减少PLAY。与LUST(性欲)一起,这些基本的社会参与系统是哺乳动物和鸟类社会依恋的基础。在人类中,至少这些系统也是各种爱的基础(Panksepp, 1998a)。我们说”至少”是因为我们不想否认其他物种也有这样的高级情感。但是动物心智的飘渺高级空间,用现有的探究方法并不容易进行科学研究。所有这些系统共享许多情感调节神经化学物质,如内源性阿片类物质和催产素。尽管最近有大量研究(Numan & Insel, 2003),但这些神经机制的大部分细节,以及导致CARE系统出现的进化路径,仍有待详细研究。然而,由于共享的神经化学物质和临近的解剖结构,我们不应忽视一个有争议的可能性:在哺乳动物大脑进化的漫长过程中,母性CARE部分地从雌性LUST的既有大脑机制和情感中出现。
谁能抗拒母亲与婴儿之间迷人的情感芭蕾?每个人都对来自对方的微妙交流极其敏感(Hrdy, 2009; Konner, 2010; Reddy, 2008)。婴儿皱眉的暗示或身体不舒服的扭动都会引起母亲安慰性的照料,而她的微笑会从婴儿那里产生一阵快乐的回应。这种精细调谐的情感互动,是人类之爱的原始来源之一,为大多数母亲提供了深刻的满足,并且对发育中婴儿的情感和身体健康至关重要。
然而,母性养育在动物王国中绝非普遍存在。许多动物,包括几乎所有爬行动物,几乎没有母性冲动。它们让幼崽在危险的环境中自生自灭,许多幼崽会死于捕食。相比之下,基本上所有哺乳动物(和鸟类)都照顾它们的幼崽,常常以牺牲自己的舒适为代价,有时甚至冒着生命危险。在哺乳动物中,初级过程的养育冲动在雌性中最强。然而,在鸟类中,父亲往往和母亲一样细心;这种现象也见于少数哺乳动物(de Jong et al., 2009)。在鱼类中,照料卵巢的工作通常完全留给父亲。这种养育冲动源于我们统称为CARE系统的固有大脑回路。
从历史上看,哺乳动物中主要过程CARE系统的存在引起了研究人员的注意,这源于一项发现:从产后雌性大鼠输血会在未生育的雌性大鼠中引发母性行为;这些行为包括筑巢、盘旋在幼崽上方以及收集从巢穴走失的幼崽(Rosenblatt, 1990)。我们仍然不知道输血中的哪些母性化学物质与未生育雌性的大脑系统相互作用以促进养育行为,但我们确实知道大脑内的催产素(oxytocin)是一种可以促进这种转变的化学物质。母性行为也可以通过电刺激大脑的特定区域来促进;这种刺激在情感上是积极的,因此具有奖赏性。这种提供CARE的冲动很可能源于SEEKING唤醒;SEEKING系统对于母亲必须做的许多事情至关重要,以便她们的后代能够茁壮成长,包括筑巢和找回幼崽。因此,CARE的大部分积极情感可能是由于大脑多巴胺的唤醒(见第3章),结合阿片类物质,以及催产素、催乳素和许多尚未被识别的大脑化学物质。
关于社会性大脑的主要过程方面的研究目前正在迅速推进。神经科学界正在认识到CARE和其他主要社会系统在大脑中的重要性。大多数神经科学家很容易接受LUST和RAGE大脑系统的存在,因为这两种情绪都清楚地表现在动物行为中,并且这些情绪显然对生存至关重要。在这一研究领域的坚实起步是在发现大脑阿片系统的社会功能后不久开始的(MacLean, 1990; Panksepp & Bishop, 1981),随后是最近对大脑催产素的研究(Carter, 1998; Insel & Young, 2001; Panksepp, 1998a)。尽管对主要过程社会系统的神经科学研究落后于对其他主要情感系统(如RAGE和FEAR)的研究,但它正在迅速赶上。然而,令人遗憾的是,许多研究人类和其他灵长类动物的研究者甚至还没有认识到主要过程社会系统的存在,更不用说其重要影响了,他们将母性和玩耍冲动设想为社会建构的。
当然,科学是相当保守的,这也是必须的。在许多研究和许多物种中积累的一致证据必须汇聚在一起,才能使一般的理论原则被吸收到标准的神经科学思想和实践中。新想法必须证明自己才能被接受。例如,正如我们所见,该领域仅仅是逐渐转向对多巴胺唤醒的SEEKING系统的概念化,该系统产生食欲性动机,同时仍然坚持长期受青睐的概念,如”大脑奖赏系统”,尽管后者存在无法解决的悖论。幸运的是,最近CARE系统已经成为大量神经科学研究的主题,这些研究由多伦多大学的Allison Fleming和波士顿学院的Michael Numan等先驱及其学生进行。(当然,实际动手研究工作的主要部分通常由博士后研究员、研究生、技术人员和无处不在的有才华的本科生完成。)此外,社会科学家对更高级、更晚近进化的社会性大脑的运作变得非常感兴趣——合作、共情和社会镜像的许多方面——这主要是因为现代脑成像技术的出现。因此,现在有大量关于灵长类和人类社会性大脑的更高级社会方面的论述(例如,Cacioppo & Patrick, 2008; de Waal, 2009; Tomasello, 2009),尽管这些研究者很少提到所有哺乳动物共有的更古老的主要过程社会冲动。社会科学家最终会弥补这一疏漏,因为他们必须这样做才能对我们的祖先根源有更全面的理解。例如,正如前面提到的,如果没有母性CARE和GRIEF的心理痛苦的基础,共情可能就不会存在(Panksepp, 1998a; Watt, 2007)。
下一章将探讨幼年动物的分离-痛苦(GRIEF)系统,该系统通过产生独特的情绪化叫声来表达它们对母亲关怀(CARE)的需求。我们从动物研究中得知,母亲会尽职地调查其幼崽发出痛苦叫声的位置,即使这些叫声是由录音机播放的。这种反应背后的适应性逻辑应该是显而易见的。痛苦的哭泣通过唤起父母的保护性注意力来促进婴儿的生存能力。然而,最近我们对这种反应的理解增加了新的层面。研究表明,当父母听到婴儿的哭声时,他们大脑中的分离-痛苦(GRIEF)区域会被激活(Swain et al., 2007),母亲比父亲反应更强烈。母亲通常也能将自己婴儿的哭声与陌生人的哭声区分开来。最近脑成像研究发现的一个含义是,父母,尤其是母亲,可能会直接体验到婴儿的痛苦,因为他们自己大脑中相应的情绪系统被婴儿的哭声唤醒。这一证据具有重要意义,原因有两个。首先,它似乎是有情感纽带的个体之间原始共情(empathy)的一个例子。其次,它强烈表明GRIEF(分离痛苦)的唤起会激活CARE(母性型养育)。因此,我们开始看到神经通路的轮廓,在这些通路中,人类共情的原始根源起源于大脑的CARE和GRIEF网络。
强大的哺乳动物养育冲动如何从爬行动物(众所周知不关心后代的动物)大脑的祖先起源演化而来?为什么哺乳动物母亲热衷于保护、珍惜甚至牺牲以确保后代的福祉?CARE系统是如何发展的?我们无法确定,但我们提出一个假设,即CARE的进化与哺乳动物祖先大脑中的LUST(性欲)回路密切交织。尽管CARE可以被视为一个非性的情绪系统,但如前所述,性欲的神经化学控制也位于养育行为的核心。换句话说,催产加压素(vasotocin)以及相关的古老神经肽如中间催产素(mesotocin),它们驱动爬行动物和鱼类的LUST和分娩系统,可能已经进化成催产素(oxytocin),催产素不仅促进雌性LUST,也是促进母性CARE的关键大脑系统(Uvnäs-Moberg, 1998)。
催产加压素是一种古老的激素,在爬行动物和鸟类中产生性冲动和调节良好的分娩反射。爬行动物通常不养育幼崽。然而,鸟类会这样做,这种养育态度是由催产加压素诱导的。为了说明催产加压素如何介导分娩行为,我们可以考虑它在海洋爬行动物(如海龟)众所周知的筑巢模式中的轨迹。在完成了数千英里的迁徙以到达其祖先分娩的海滩后,母海龟登上海滩并挖掘巢穴;当她这样做时,她的垂体后叶分泌的催产加压素不断增加。当她一个接一个地产下卵时,催产加压素水平进一步升高。然后,当她的分娩完成并覆盖卵时,催产加压素骤降至微不足道的水平。因此,她的母性冲动和职责就此结束。她返回大海。数周后,当她的后代孵化时,它们迅速向海浪奔去,容易受到攻击且没有父母保护。只有一小部分后代逃脱捕食并延续物种的生存。
在LUST章节中,我们提到催产加压素具有类似于催产素在哺乳动物中作用的镇静效果,并在许多鸟类物种中促进养育情绪(Adkins-Regan, 2009; Balthazart, et al., 1996; De Vries & Panzica, 2006)。实际上,催产加压素可能进化成了哺乳动物的化学物质催产素和加压素(vasopressin),它们分别在控制哺乳动物的雌性和雄性性欲方面发挥重要作用。在这里,我们特别关注催产素的进化联系,因为除了是产生雌性LUST的重要化学物质(Caldwell, 2002)外,催产素也是一种重要的母性化学物质。它通过产生子宫收缩在分娩中发挥核心作用,并在分娩后通过触发乳汁下降(当乳头被吮吸时)发挥作用。此外,催产素与其他母性化学物质一起,在注入未生育雌性的大脑时有助于促进母性情绪和行为(Pederson et al., 1982; Keverne & Kendrick, 1994)。
从祖先肽到催产素的进化化学连续体表明,哺乳动物大脑中的CARE系统可能部分地从爬行动物的LUST系统进化而来。一种功能从另一种表面上看起来完全不同的功能进化而来并不罕见。诺贝尔奖获得者、分子生物学家弗朗索瓦·雅各布(François Jacob)写道:“自然选择……像一个修补匠一样工作……他……利用周围找到的任何东西……来制造某种可行的物体……进化从一条腿制造出一只翅膀,或从一块颌骨制造出耳朵的一部分……自然选择……不会从零开始产生新事物。它在已经存在的基础上工作”(Jacob, 1977)。
Jacob谈到的是外适应(exaptation)原则,指的是进化变化中,现有结构看似激进的改变产生了可用于新适应目的的结构。外适应的一个例子是鱼类支撑鳃的弓形结构,进化成了哺乳动物中耳中用于听觉的骨骼。外适应指的是进化使物理结构适应新用途的令人惊讶的,而非显而易见的路径,包括将古老的大脑过程转变为执行新功能的系统。因此,虽然鳃弓在鱼类中帮助呼吸,但哺乳动物中耳中产生的骨骼则促进听觉。尽管我们可能感到不适或有其他审美上的反对意见,但看似与性无关的照料(CARE)系统可能是从祖先物种的性欲(LUST)系统进化而来,这并不违反自然法则。
与所有进化步骤一样,通过外适应带来的变化会根据其适应价值而被保留或放弃——变化的保留将取决于它如何帮助动物在其环境中生存。可以推测,对幼崽的养育照料具有适应性,因为父母的照顾和保护为所有哺乳动物物种的生存提供了决定性的竞争优势。如果一个幼小的动物只能通过从另一个动物那里获取食物来生存,那么没有什么比社会联结和母爱更能确保这种分享的发生。正如我们将看到的,所有哺乳动物的情绪健康都与这种早期照顾和关爱的质量密切相关。
照料系统的发现表明,弗洛伊德的著名假设并非完全错误,他认为性是所有积极人际社会关系产生的基本冲动或驱动力。也许他的想法隐含着他没有认识到的性欲(LUST)和照料(CARE)之间的内在关系。然而,其他基本的社会情绪系统并未出现在他对人性的看法中。例如,对社会联结非常重要的恐慌/悲伤(PANIC/GRIEF)和玩耍(PLAY)系统,与性欲(LUST)系统没有明显的关系。弗洛伊德假设所有非性爱,甚至母爱,都是潜在性冲动的升华(sublimation)。他坚持认为升华——将基本情绪能量引导到社会有用目的——发生在性冲动被转化为能够服务于非性目的的社会价值观时(Moore & Fine, 1990)。例如,大多数社会都有乱伦禁忌。弗洛伊德认为,禁止乱伦的道德规范可能源于母亲获得性满足的基本愿望,将其塑造为照顾孩子的非性满足。因此,经典弗洛伊德理论家相信,无疑现在仍有一些人相信,母性冲动(以及其他柏拉图式的依恋)是潜在性冲动的升华变体。我们现在可以用新的进化方式来看待这个旧假设。
尽管照料(CARE)可能是从性欲(LUST)进化而来,但这两个系统现在在大脑中已经足够独特,并且执行不同的功能。性欲(LUST)系统产生性冲动,而照料(CARE)系统产生非性的温柔,尽管由于共享的调节脑化学物质,一些相关感受可能具有类别相似性。无论如何,我们现在知道,许多社会依恋来自性冲动升华之外的其他来源,包括下两章总结的悲伤(GRIEF)和玩耍(PLAY)系统。当然,性也会导致社会依恋。人们可能想知道婚姻制度是性冲动的表达,还是性能量的升华。我们可以合理地得出结论,婚姻包含两者:它可以成为竞争冲动和需求之间的实际妥协,这些冲动和需求包括性伴侣的快乐以及繁殖和照料的经济考量。这可能是紧迫的进化要求之间的实际文化妥协。
催产素(oxytocin)是主要的母性化学物质之一,在雌性大脑中的产量大于雄性大脑(Jirkowski et al., 1988)。雌激素介导整个下丘脑前部细胞区域催产素的产生,包括室旁核(PVN)和背侧视前区(dPOA)。这两个催产素能大脑区域对照料行为的相对重要性已在大鼠损伤研究中得到证明。室旁核(PVN)的损伤可以显著减少初次做母亲的大鼠的母性行为,但对有经验的母亲则不然。然而,背侧视前区(dPOA)的损伤可以完全消除母性行为(完整概述见Numan & Insel, 2003)。
催产素与神经系统的所有化学信使一样,如果不与特定的化学受体结合,它将毫无用处。雌激素和孕酮水平控制着催产素释放的许多区域中催产素受体的数量,包括终纹床核(BNST)和腹内侧下丘脑(VMH)。终纹床核(BNST)似乎在调节分离痛苦方面发挥着更重要的作用。在这个大脑区域,催产素似乎激发了母亲在婴儿走失和哭泣时感受到的焦虑:母亲会经历一种令人寒栗、恐惧的感觉,直到她们找到自己的婴儿,而这种母性体验支持婴儿的生存。
尽管刚才提到的大脑结构是关键的关爱系统参与者,但该系统的回路在大脑的内侧皮层下区域广泛延伸,与许多对有效母育至关重要的子系统相连。例如,有一个独特的回路控制泌乳反射。这个回路从中脑外侧区域下降到支配乳头的脊髓节段(Hansen & Kohler, 1984),从生理上让母亲做好哺乳准备。虽然只有母亲能哺育幼儿,但养育回路并非雌性的专属领域。雌性和雄性都有能力养育后代。而且,如前所述,许多关爱回路也存在于雄性大脑中(de Jong et al., 2009)。
关爱回路的一个方面值得特别强调。该系统的一个分支从产生多巴胺的腹侧被盖区(VTA)通过下丘脑延伸(Numan, 1990)到寻求系统的核心。关爱系统的这一部分很可能唤起寻求冲动,这无疑促进了目标导向的母性觅食倾向,这对筑巢和找回幼崽尤为重要。事实上,向腹侧被盖区注射催产素会促进这种母性行为,表明寻求系统对母性生活的实际和食欲需求很敏感。我们再次看到,大多数其他情绪系统利用原始寻求系统来实现其充满情感、面向行动的功能的方式之一。
母性冲动也得到附近各种情绪系统的支持,如分离痛苦的恐慌/悲伤系统。如前所述,该系统包括终纹床核,该核也富含催产素网络。正如我们将在下一章看到的,催产素在减少分离痛苦方面非常有效。大概,当悲伤系统被唤起时,由此产生的负面情感状态可以增强养育成人的照顾冲动。也许这就是为什么能够认同婴儿痛苦的母亲会有动力养育和安慰他们。这个假设值得比目前得到的更多实验关注。在大多数已研究的物种中,婴儿的分离叫声会引起母亲的强烈关注和接近行为,父亲的反应程度稍弱。如前所述,尚不清楚这在大脑中是如何发生的,但终纹床核的联系提供了一个有希望的开端。
虽然雌性可能更具母性和关爱,但雄性在体质上也能够养育。例如,通过简单地让幼年雄性大鼠和未育雌性大鼠(这两组通常会避开婴儿)每天接触幼鼠,就可以诱发它们的养育行为(Rosenblatt, 1967)。这个过程被称为敏化(sensitization)。没有人知道为什么接触幼年动物会使幼年大鼠的关爱系统敏化,但我们假设它以某种方式促进并强化了已知能唤起该系统的化学变化,例如催产素活性的增加。敏化在非常年幼的雄性中更成功,也许是因为在青春期开始时冲刷雄性大脑的睾酮会促进明显的攻击性情感色彩,这往往会抵消养育感受。(记住,雄性中的睾酮促进加压素合成,这不是一种养育化学物质,尽管它可能在父母保护幼崽方面起重要作用。)相比之下,未育雌性在青春期后更容易被敏化,因为青春期雌性身体产生雌激素,雌激素促进催产素的产生。
有充分理由相信,关爱系统在养育照顾者(包括雌性和雄性)中产生积极情感。除了由催产素驱动外,它还由内源性阿片类物质驱动。事实上,内源性阿片类物质在所有积极的社交互动中都发挥作用。催产素和内源性阿片类物质都是舒缓的”感觉良好”化学物质,已知可以抑制攻击性和易怒性(McCarthy, 1990; Siegel, 2005)。这些化学物质处于高水平的关爱母亲表现出自信的能干态度(Kinsley & Lambert, 2006, 2008),“照顾和交友”的冲动(Taylor et al., 2000),有时甚至可以升级到一种母性狂喜。
然而,成年雄性大脑中睾酮的攻击性效应可以抵消关爱冲动,甚至促进杀婴倾向。在动物王国,雄性经常杀死自己物种的幼崽,但通常不会杀死自己的后代。这可能是幼年动物在成年雄性面前通常表现出比在雌性面前更多恐惧的原因之一—这种倾向可以进一步削弱幼年动物与成年雄性之间的联系。然而,催产素似乎能抑制雄性杀婴的倾向(McCarthy et al., 1992)。如上一章所述,雄性大鼠在交配后不太可能杀婴。这种和平倾向逐渐增加,并在雄性自己的后代出生时达到峰值(Mennella & Moltz, 1988)。我们知道性活动会导致雄性大脑中催产素的产生。也可能雄性大脑中催产素系统的这种升高活性会继续增加,直到它们的后代出生所需的时间。这可能是雄性在受精后几周对幼年动物攻击性较低的原因吗?我们不知道,但这是一个合理的假设,值得热情的情感神经科学家探索。
我们已经强调了催产素在关爱唤醒中的作用,这种作用最典型的体现就是母亲养育新生儿的冲动,同时它对母亲和孩子都有丰富的积极抗压效果(Uvnäs-Moberg, 1998)。催产素的生成由雌激素控制,雌激素在整个孕期保持在适度水平,随着分娩临近而增加。因此,在妊娠末期会出现由雌激素介导的催产素活动激增。有趣的是,多年来研究人员并不认为催产素是一种重要的母性化学物质。许多人认为催产素的唯一来源是垂体后叶。当从哺乳期母亲身上切除这个腺体并未抑制母性行为时(Slotnick, 1975),科学家们认为母性行为并不依赖于催产素。只有在发现大脑深处存在催产素能系统(使用催产素的系统)后,神经科学家才开始考虑催产素可能在大脑中分泌,并可能在产生筑巢、哺育幼崽、盘旋在它们上方提供温暖等母性行为中发挥核心作用(大脑和/或脊髓中的作用)。
然而,要明确催产素在母性行为中的确切作用需要付出一些努力,因为其效果并不总是清晰的。虽然直接向大鼠和绵羊的脑室系统注入催产素通常会促进母性倾向(Pedersen et al., 1982, 1992),但有时这样的实验并不产生母性行为(Bolwerk & Swanson, 1984)。当研究人员发现未产雌性大鼠通常觉得新生幼崽的气味令其厌恶,而这本身就可以抵消注射催产素产生的母性倾向时,这种明显的矛盾得到了解决(Fleming & Rosenblatt, 1974)。然而,当母鼠第一次分娩时,它对幼崽气味的习惯性厌恶被吸引所取代。当未产雌鼠输入产后大鼠的血液以及通过每日接触幼崽而敏化时,也会看到这种相同的现象。导致这种对新生儿厌恶快速逆转的完整因素阵列尚不清楚,但显然这种变化并非仅由催产素引起。因此,催产素以外的化学物质必定在产生母性冲动中发挥着重要作用。
我们可以通过研究分娩期间和之后发生的化学变化来了解这些化学因素可能是什么,分娩是诱发关爱行为的典型体验。分娩在大脑中产生了戏剧性的转变。孕酮水平以及雌激素和催产素水平在整个孕期都很高。然而,随着分娩临近,孕酮水平会骤降。众所周知,高剂量的孕酮在大脑中起镇静作用,几乎像麻醉剂一样。也许这种激素的消退突显了母亲身份需要提高警觉性和对细节的关注这一事实。另一方面,催乳素随着分娩临近而急剧上升,诱导乳汁的生成并在产生母性感受和行为中发挥作用。虽然这类研究主要在大鼠和绵羊身上进行,但哺乳动物的相似性表明这些发现很可能适用于包括人类在内的其他哺乳动物。
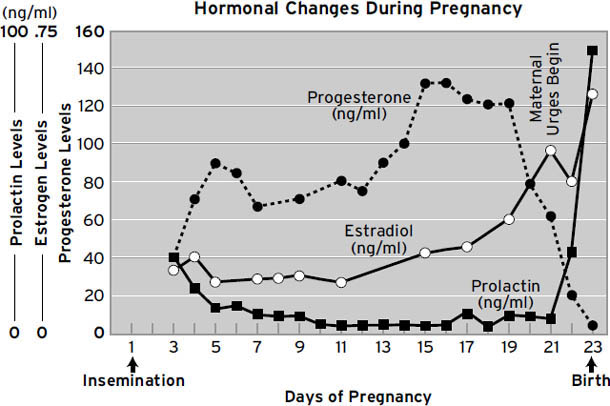
图8.1. 大鼠妊娠期间孕酮、雌二醇和催乳素的循环水平。这些激素在妊娠期间的变化,以及孕酮的快速下降和分娩前几天雌激素和催乳素的大幅升高,建立了促进母性行为的生理条件。人们认为,母亲大脑中催产素的激活是实现这种关爱动机变化的最重要效应之一(改编自Rosenblatt, 1990的原始数据,如Panksepp, 1998a所示;经牛津大学出版社许可重新发布)。
目前尚不清楚这些化学转变是否解释了雌性大鼠对幼崽气味吸引力的变化。然而,我们确实知道这些化学转变在许多母性行为和情感的产生中发挥着关键作用。此外,这些转变可以被人工诱导。只需用正确的激素注射模式模拟分娩前的激素变化即可;这是通过快速提高雌激素、催产素和催乳素水平并降低孕酮水平来实现的。这种化学物质丰富和耗竭的模式可靠地诱导产生母性渴望的大脑转变。因此,催产素只是母性化学物质之一,也许它并不那么至关重要。事实上,催产素对于大鼠产生母性冲动并非必需。通过消除制造催产素的相关基因而使催产素系统被”敲除”的小鼠仍然可以表现出充分的母性行为。然而,它们的幼崽无法存活,因为没有催产素就没有乳汁生产;这些小鼠幼崽必须有”乳母”才能存活(Pedersen et al., 2006)。当然,这些动物的母性行为可能不如大脑充满催产素的母亲那样令其满足、尽心和有活力(Nishimori, et al., 2008)。
其他研究帮助界定了催产素在创造关爱态度方面的局限性。例如,虽然催产素明显有助于启动母性冲动,但一旦母性行为建立起来,催产素对于胜任的母性行为就不再那么重要了。如果在母鼠第一窝幼崽出生时,通过给予阻断催产素受体的药物来抑制催产素在母鼠大脑中的影响,母鼠不会迅速表现出母性热情。她似乎对幼崽的心理和生理诱惑产生了抵抗。如果几天后进行同样的操作,母性行为就不会受到严重破坏。它已经成为一种习惯。如前所述,如果在分娩前损伤下丘脑的室旁核(PVN)——大脑中一个含有大量催产素分泌神经细胞的关键区域——也会出现母性情感的缺失。这种操作不会破坏分娩过程本身(van Leengoed et al., 1987),但确实会干扰最初的母性能力。然而,如果允许初产母鼠在这种受限的脑损伤之前与幼崽相处几天,她的母性能力就完全不会受到破坏(Insel, 1990; Insel & Harbaugh, 1989)。显然,母性关爱系统帮助大脑快速学习关于母性能力的许多知识。然而,这些养育倾向很快就会根深蒂固地成为习惯,以至于不再依赖催产素的”魔力”。
然而,人们不禁要问,阻断有经验母鼠的催产素作用是否要付出长期代价。当母鼠有过往的母性经验时,催产素在短期内对于胜任的母性行为并非必需。但催产素对于长期的母性能力是否必需呢?在正常情况下,即使在母性行为已经习得之后,催产素仍然会在母鼠大脑中释放。据推测,这会产生养育冲动和相关的情感感受,对母鼠来说具有高度奖赏性。长期阻断催产素可能会干扰母鼠履行母性职责时的愉悦感。这种主观愉悦感的减弱可能会导致母性能力随时间恶化。这种情况在动物模型中还没有得到充分研究。我们也缺乏证据表明这些原则在多大程度上适用于人类。
诚然,催产素在维持母性情感和行为方面的具体长期作用尚不清楚。但大脑化学物质经常通过协作发挥作用。例如,有证据表明催产素会增强阿片类物质的作用。因此,我们得出一个合理的假设:催产素可能在长期内支持母性行为,因为母性能力与所有积极的社会关系一样,会被极低剂量的阿片类物质所增强(Panksepp, 1998a)。我们将在恐慌/悲伤章节中看到,当动物从事积极的社会互动(如相互梳理毛发)时,它们的大脑会分泌内源性阿片类物质。
目前尚不清楚母性满足感是否由大脑自身的阿片类物质介导。然而已知的是,极低剂量的阿片类药物会增强许多积极的社会互动,包括母性行为和玩耍。另一方面,即使是稍高剂量的阿片类物质也会诱导一种极乐但社交反应迟钝的迟缓状态:在这些情况下,幼年动物表现出较少的玩耍,动物总体上不太合群,母鼠表现出较少的母性行为。据推测,当母鼠处于完全满足的阿片类状态时,她们无法体验到由良好母性行为产生和所需的情感波动变化。然而,以微小剂量给予时,阿片类物质会促进以友好方式互动的冲动。因此,低剂量的阿片类物质也能增强母性行为就不足为奇了。如果最终确凿地发现催产素通过内源性阿片类大脑化学物质促进这种增强作用,也不会令人惊讶。
正如我们在欲望章节中讨论催产素作用时所提到的,催产素的力量有时可能是间接的,通过增强内源性阿片类物质的作用来实现。反复暴露于高剂量阿片类物质后,大脑通常会对这些药物产生越来越高的耐受性或不敏感性。这种耐受性是成瘾者需要越来越大剂量才能获得他们所渴望的积极感受的主要原因。催产素会降低阿片类耐受性,这意味着在催产素水平充足的情况下,少量阿片类物质会继续产生舒适、愉悦的效果(Kovács & Van Ree, 1985)。哺乳期母鼠的大脑会分泌催产素和可能的内源性阿片类物质,这种组合可以在很长一段时间内维持令人满意、舒适的效果。这可能是为什么哺乳对许多母鼠来说仍然是一种愉快的体验。如果催产素被阻断,那么内源性阿片类物质持续产生的愉悦效果可能会逐渐减弱,导致过早断奶。这也可能会整体降低母性表现,导致幼崽出现各种发育问题。
当然,我们确实看到一些母亲以敷衍而非共情的方式照顾她们的婴儿。这些母亲的大脑中母性化学物质可能不如那些表现出更明显奉献精神的母亲充足。初级过程母性化学物质的缺陷可能导致情感敏感性降低。我们可以推测,这类化学物质水平较低的母亲只要孩子没有经历异常程度的痛苦,就能提供足够的养育,但某些母亲在更极端的条件下可能无法提供共情支持。相反,当悲伤(GRIEF)在更极端的情感条件下启动时,冷漠的养育可能会被抛到一边,有力地弥合这种原始的共情差距。同样,这些问题需要通过更多研究来解决。
显然,我们对催产素(oxytocin)和其他社会化学物质作用的理解仍不完整。我们可以确信催产素对母性行为的初始产生很重要,因为在动物模型中,催产素阻断剂在减少养育行为的开始方面发挥着决定性作用。然而,单独的催产素并不能消除处女雌鼠对幼鼠气味的最初厌恶,这表明在母性的这些关键初始阶段还有其他化学物质在起作用。内源性阿片类物质的释放肯定是维持母性冲动的反馈的一部分:如果给动物注射非常低剂量的阿片类药物,母性行为会变得特别旺盛,但稍高的剂量会削弱它,这可能部分是因为母性动机受到大脑阿片释放自然波动的调节。高于正常水平会普遍降低社交欲望。此外,虽然催产素对于有经验的母亲产生母性行为不是必需的,但如果催产素系统普遍无反应,母性行为就会很弱(Nishimori, et al., 2008)。
研究人员目前正在调查催产素回路的许多角落和缝隙的作用,以阐明催产素分泌产生母性行为变化的确切方式。我们之前提到催产素进入腹侧被盖区(VTA)的多巴胺神经元,这些细胞在促进寻求(SEEKING)冲动方面非常重要。此外,在分娩后,关键的催产素回路通过间隙连接(gap junctions)的发展而同步,间隙连接是相邻神经元之间的直接原生质桥梁。间隙连接允许神经元之间快速的非突触协调(Modney & Hatton, 1990)。这有助于催产素能神经元协同作用。例如,通过这种同步化,婴儿的触摸会迅速引发射乳反射。然而,催产素回路的大部分仍然是个谜,特别是在调节人类社会感受和行为方面。
关于其他与妊娠相关的化学物质:催乳素(prolactin)和随着临产而变化的各种类固醇,我们所知更少。然而,已知孕酮(progesterone)和雌激素(estrogen)都能促进某些大脑催产素系统的重塑。我们也知道催乳素促进乳汁合成,同时在已详细研究的几个物种中促进母性行为,特别是鸟类。虽然催乳素是一个非常大的分子,但它会从血流中主动吸收进入大脑,直接注射到大脑会促进动物的母性倾向(Walsh et al., 1987)。而且,如上所述,孕酮水平的降低对母性行为的开始也很重要(Sheehan & Numan, 2002)。
这些神经化学研究大多在大鼠、绵羊和鸟类身上进行,但关于这些系统在人类大脑中的运作,越来越多的信息正在变得可用。目前正在进行大量关于鼻内给药催产素的研究(这是已知的将神经肽送入人类大脑的唯一方法)。这项研究的总体发现是人们倾向于变得更亲社会,即不那么有攻击性,更信任他人,并且在处理社交事务时总体上更自信(MacDonald & MacDonald, 2010)。这些结果与我们对动物大脑中这些问题的深入理解基本一致。因此,考虑到哺乳动物情感系统的同源性,以及所有哺乳动物(包括我们自己的物种)普遍表现出母性行为(物种间的细节有很大差异),很可能类似的原则在所有哺乳动物的大脑中起作用。换句话说,相同的母性化学物质交响乐在整个哺乳动物物种中促进关怀(CARE)系统的活动。
当然,在人类中无法判断养育行为有多少是由照顾(CARE)冲动驱使的,又有多少是由有意识的认知决策引导的。作为所有生物中最聪明的物种,人类能够从认知上理解父母双方照顾孩子的重要性。这一事实导致早期研究者提出这样的观点:人类没有母性本能,而是完全通过学习才对孩子产生依恋。传统上,人类男性不照顾婴儿。在现代,对养育孩子重要性的有意识认识说服许多父亲参与育儿。另一方面,母亲有更强的生物冲动去接触婴儿并提供照顾。因此,在认知和文化问题的巨大复杂性之下,存在着从事育儿的生物情感动机。由于这种生物差异,大多数参与婴儿照顾的人类父亲可能以一种更常规、不那么深刻情感化和共情的方式来养育孩子,而不像母亲那样。母亲通常表现出更自然的温暖和与婴儿在一起的渴望。通常也是母亲更持久地与婴儿进行敏感的情感交流,特别是与快乐的婴儿交流,但也与处于痛苦中的婴儿交流。
有充分的理由相信,许多母性化学物质在人类母亲的母性冲动中起着核心作用。这些神经化学物质促使临近预产期的孕妇在孩子出生前”布置她们的巢穴”。当婴儿出生时,一系列原始过程的大脑化学物质通常有助于确保母亲的照顾所带来的快乐超过负担。当然,因为我们是聪明、有思想的生物,我们人类也足够聪明,在婴儿到来前几个月就开始计划。我们购买衣服、婴儿床、摇篮、尿布等等。然而,即使是人类母亲似乎也会经历分娩前的一段时期,在这段时期她们会进行一连串强迫性的准备活动来迎接婴儿的到来。这可能是由于预示分娩过程的许多化学变化。进化似乎并不完全依赖学习来确保母亲为婴儿到来等关键生活事件做好准备。例如,母性化学物质对大脑的影响甚至使雌性更不容易焦虑(Kinsley & Lambert, 2006)。这造就了更好的母亲,而好母亲对孩子投入的额外关注为她们后代的心理和神经优势带来了终生的益处。然而,学习是每个情感系统的一部分。在复杂的认知结构方面尤其如此,这些结构对每个个体都是独特的,它们缓冲或加剧我们的基本冲动,并影响我们对这些本能指令的行为反应。
最后,从临床角度来看,在妊娠末期和分娩期间通常会改变水平的化学物质,通常会促进母亲能力,但也可能出错。由此产生的身心状况有时是有害的。一些母亲因尚未完全了解的原因陷入抑郁。我们确实知道,一些产后抑郁和精神病病例与循环中高水平的β-酪啡肽相关,这是一种源自牛奶的阿片肽。然而,我们并不知道这导致了抑郁。这种状况可能对母亲和孩子都产生灾难性影响,通常通过服用传统抗抑郁药和/或心理治疗来治疗。如果我们更好地了解照顾(CARE)和分娩系统的药理学基础,我们也许能够以更具体和有效的方式治疗有害的母性反应。需要进行的一项研究是评估鼻内给予催产素是否能够缓解那些在孩子出生后不久经历抑郁的母亲的绝望感。在精神分析治疗的背景下,Andrea Clarici在意大利东北部美丽的亚得里亚海边的里雅斯特的研究小组已经启动了这样一个项目。我们热切期待结果。此外,精神分裂症通常以社会纽带失败为特征,最近发现鼻内催产素能够缓解那些陷入精神病的人的阳性症状(例如幻觉)和阴性症状(社交退缩)(Feifel et al., 2010)。
尽管在所有被研究的哺乳动物物种中,母性脑回路都是相似的,但与所有情绪系统一样,每个物种显然都有独特的特征,促进不同强度和模式的母性行为。例如,兔子的育儿时间短暂且不频繁,每天只给隐蔽的幼崽喂食一次,其余时间用于啃食营养价值一般的食物。母兔似乎也缺乏动机或神经行为装备来找回从隐蔽巢穴中分散的小兔子。我们不知道兔子是否真的与它们的后代建立联结。事实上,社会联结只在某些物种中得到研究,令人惊讶的是,我们甚至不知道大鼠和小鼠等常见实验动物是否真的与它们的后代建立联结。在生命最初几周,当它们的后代仍是”早产儿”(运动能力不足,因此无法自己走失)时,它们实际上不需要建立联结。相比之下,联结对食草动物(如有蹄类动物)至关重要,这些动物的后代在出生几小时后就能蹦跳和奔跑,因此容易走失。这些早熟物种的母亲通常会快速且专一地与自己的后代建立联结,而那些幼崽出生时不成熟(晚成性)的物种则乐于收养其他幼崽。
由于生态因素,不同物种表现出不同的联结窗口——母亲和婴儿能够彼此依恋的最佳时间间隔。当动物以晚成状态出生,眼睛和耳朵仍然闭合时(这在捕食者物种中很常见),幼崽无法远离巢穴。在这些动物中,联结窗口很大,可以持续出生后许多周。对于有早熟后代的食草动物(如绵羊)和鸟类(如小鸡和小鸭,它们可以在出生后不久就与母亲一起觅食),联结窗口在出生后几小时内关闭(对于绵羊)或最多一天后关闭(对于小鸡和小鸭)。这个短暂的联结窗口反映了母婴联结和照料回路与婴儿出生时的活动能力相匹配。如前所述,这类猎物物种通常出生时活动能力很强,因此它们可以跟上不断移动、觅食或逃避捕食者的母亲。它们经常被同类包围,成群结队,这减少了被捕食的危险,但可能带来其他问题。在这种情况下,幼崽很容易在出生后不久就与父母分离,从而在众多动物中走失。因此,在早熟物种中必须非常快速地形成联结,因为幼崽很容易自己走失。在人类和有不成熟幼崽的生物中,短暂的联结时间窗口就不那么重要了。
在绵羊中,母亲在出生后很快与幼崽建立联结,并能通过气味识别自己的幼崽。这种安排的缺点是,如果母亲在出生后不久失去与幼崽的接触几个小时(这可以通过实验完成),它们通常会在重逢后忽视后代,实际上如果后代试图吃奶会拒绝它们。换句话说,那些不幸在母亲有机会将它们识别为自己的后代之前走失的幼崽,在重逢时会被当作陌生者对待。显然,我们人类和许多其他杂食动物的行为更像肉食物种,其幼崽通常出生时非常不成熟。因此,我们可以更容易地将陌生者收养到我们的照料圈中。我们将回到有蹄类动物的短联结窗口如何帮助研究人员解码联结的神经化学本质,但现在让我们关注人类养育子女的一些关键方面,这些方面可能对我们自己相对开放的社会联结系统有影响。
人类婴儿有一个非常大且长的社会联结窗口。他们也可以很容易地与非父母照料者建立联结,尽管对生存至关重要的最初社会联结最常见于母亲和亲生婴儿之间。有理由相信,在我们的祖先环境中,养育子女更多是一种自发的群体活动,而不像我们许多现代文化中那样。例如,在传统文化中,婴儿由大家庭照顾并不罕见。在大部落家庭的安全范围内,独立性也受到鼓励。婴儿通常在1岁时就接受如厕训练,此后不久他们就被鼓励作为紧密联系的大家庭的相对独立成员行事(Hrdy, 2009; Konner, 2010)。在我们西方的核心家庭文化中——这可能是在过去千年中出现的——大多数1岁孩子的父母还没有开始考虑如厕训练。儿童被允许在童年的前两三年保持婴儿行为。父母在这些成长期间对他们的监督也非常密切。我们还珍视亲子关系的排他性,很少给孩子在更大社区中独立行动的机会。显然,社会成熟必然会受到许多人养育儿童的社会环境的促进,正如一句著名的非洲谚语所说:生一个孩子只需要一个女人,但养育一个孩子需要整个村庄。我们的文化似乎不再遵循这样的格言,这可能只会放大养育子女的阴暗面,我们现在将考虑这一点。
母性冲动虽然强大,但并非绝对。有些怀孕是意外的。有些人类母亲会遗弃她们的婴儿,将他们托付给大家庭成员、教会或社会服务机构照顾。这种情况通常发生在母亲没有足够资源抚养孩子的时候。这种做法在孩子年龄较大时也变得更加普遍,尤其是当孩子随着青春期的到来而接近独立时。纵观历史,虐待儿童长期以来一直是许多文化景观的一部分,当时同理心的文化价值较低(Rifkin, 2009)。这种做法在我们史前祖先的过去是否不那么常见很难说,但当人类群体主要由大家庭组成时,这似乎是可能的。
资源可获得性始终是母亲关注的问题。在资源短缺时期,老鼠母亲吃掉幼崽的情况并不少见,尤其是当资源匮乏到严重损害母亲将后代抚养到青春期的能力时。另一个常见行为是多胞胎的母亲忽视较弱个体的需求,而没有明显的悔意。这种情况很少发生在一次只生一个后代的物种中。但在人类中偶尔也会发生。在某些传统社会中,婴儿的命运往往取决于经济或实际考虑而非生物学因素。例如,在不太遥远的过去,一些社会,如加拿大北部的内特西利克爱斯基摩人,认可杀婴行为,特别是杀害女婴,以避免未来的社会问题。那些几乎没有希望找到合适配偶的女婴,因为部落中没有出生年龄相当的男婴,会被遗弃在雪中等死,父母几乎没有表现出明显的痛苦或悔恨(Riches, 1974)。长期的社会关切压倒了短期的情感关切。直到今天,在重男轻女的社会中,女婴而非男婴被杀害的可能性要高得多。这种做法的先例可以在一些动物物种中找到:如我们上面所述,一些母亲会杀死较弱的幼崽。当环境资源稀缺时,这种做法可以增加幸存后代的成功概率。因此,对后代的投入程度只是部分情感问题。
然而,在人类母婴依恋形成过程中的情感问题是巨大的——我们有着延长的婴儿期和童年期——特别是现在我们的孩子不是在大家庭中抚养的。人类的大脑结合机制,就像许多其他食肉动物和杂食动物一样,构成了一个漫长的事件过程。一般来说,母亲似乎在情感层面上与婴儿建立联系的速度相当快。另一方面,婴儿有一个相当更宽和更灵活的结合窗口期。人类婴儿的依恋通常要到大约一岁时才完全形成,这使得他们在此之前可以被寄养到有支持能力的家庭而不必太担心。然而,一旦形成,依恋的安全性就至关重要(如下一章所述)。了解依恋如何在婴儿和母亲的大脑中形成是至关重要的。关于人类大脑中这些神经过程,我们基本上没有直接证据。因此,有必要从动物模型研究中进行推广。只有当我们认识到我们正处于这一发现领域的起点,大部分前沿领域还在我们面前时,才能对我们现有数据的相关性进行合理和平衡的评估。我们对这些重要的心智大脑功能的理解才刚刚起步。它们只在少数几个物种中得到了详细研究。而最好的研究是在那些表现出狭窄结合窗口期的物种中进行的;这些物种可能不是阐明人类结合的最佳模型。
让我们沿着这些思路深入到潜在的神经机制中。
我们现在知道催产素(oxytocin)(Popik et al., 1992)和加压素(vasopressin)(Dantzer et al., 1987, 1988)都能增强社会记忆,因为当这些神经肽的活性被阻断时,在迄今为止研究的所有物种中,社会记忆都是微弱的且形成缓慢。因为加压素和催产素都能促进动物的积极社会和性行为(第7章),这些相同的化学物质有助于促进友好的社会纽带并参与创建关于这些纽带的记忆,这也许并不令人特别惊讶。当然,这是自然经济方式的另一个例子,给了我们一些希望,即我们从其他动物身上学到的东西也将适用于我们。
大约从哺乳动物分娩开始,大多数母亲体内的雌激素水平升高,这反过来促进催产素以及编码新兴母性冲动所需受体的产生。由于催产素促进社会记忆的形成,可以合理地推测它也使母亲能够记住自己的后代。对产后母羊的研究表明确实如此。除了在产生生理和情感母性反应中的作用外,催产素还与去甲肾上腺素(NE)在社会过程中的利用有关(Kendrick et al., 1992; Levy et al., 1993)。去甲肾上腺素是另一种大脑化学物质,在母羊嗅觉记忆的形成中发挥重要作用。当母羊嗅球中形成促进与后代积极社会互动的新神经通路时,就会形成积极的嗅觉社会记忆。如果去甲肾上腺素活性被阻断(通过拮抗剂普萘洛尔),这些通路就不会那么多。嗅觉记忆因此受到损害。结果,接受去甲肾上腺素阻断剂的母羊明显不太能够区分自己和其他羔羊(Levy et al., 1995)。在正常情况下,这些大脑机制在绵羊等有蹄类动物中运作非常迅速,有助于确保母亲为自己的后代提供专属的哺乳权。
去甲肾上腺素似乎以下列方式促进嗅觉记忆的形成:当在嗅球中释放时,去甲肾上腺素降低GABA的活性,而GABA是哺乳动物神经系统中主要的抑制性神经递质。当GABA在嗅球中的抑制性影响减少时,相关的气味编码神经元变得更加活跃,放电更加剧烈。这种快速放电稳定了编码羔羊气味的神经通路。当通路稳定时,它会随时间持续,这意味着它成为记忆通路。显然母亲就是通过这种方式对婴儿独特的气味特征产生依恋的。类似的过程可能也在婴儿中运作,甚至在人类中也是如此,尽管人类与其他哺乳动物相比嗅觉相对较弱。相当多的研究表明,人类婴儿对母亲乳房的气味发展出选择性吸引力。大概这种吸引力通过大脑中催产素释放产生的积极情感感受得到强化,以及通过相关的”感觉良好”神经肽(如内源性阿片类物质)的唤起得到强化。在许多物种中,这种社会吸引力通过温柔的触摸得到进一步强化,触摸刺激已知能促进大脑阿片类物质和催产素的释放(Panksepp, Bean et al., 1980; Matthiesen et al., 2001)。
事实上,社会联结可能是一种成瘾现象,这是Panksepp和他的学生们首先推测的。社会依恋与人们对阿片类物质使用形成的联结之间的关系具有非常相似的特征(见图8.2)。正是这个想法促使我们开始对社会依恋本质进行首次神经科学探究(见第九章)。然而,涉及许多其他化学物质这一事实并不令人惊讶。每个大脑功能都由多种大脑化学物质介导。尽管阿片类物质假说经受住了时间的考验,正如我们已经看到的,催产素在使母亲过渡到与婴儿持续的照料关系方面非常有影响力,产生持续的养育。

图8.2. 第一个关于社会联结是成瘾现象理论的概念总结,基于阿片类物质依赖的动态特征与社会依恋关键特征之间的主要相似性。两者显示出非常相似的心理动态,这表明阿片类物质和其他一些成瘾在情感上如此引人注目,是因为它们利用了相同的大脑情绪系统。这个想法最初是在20世纪70年代末提出的(摘自Panksepp, 1998a;经牛津大学出版社许可再版)。
我们之前提到母羊有一个非常短的联结窗口期(bonding window),持续不超过几个小时。如果母亲在这个时间窗口内无法接触到她的羔羊,促进母性吸引力的社会记忆通路将不会在她的嗅球中形成,她也不会对羔羊的气味产生诱人的印记。然而,研究人员发现,如果对母羊的阴道和子宫颈提供刺激(Keverne et al., 1983; Kendrick et al., 1992),通过一个直接称为阴道-子宫颈刺激(VCS)的程序,联结窗口期可以被人为地重新打开几个小时。VCS在母羊的中枢神经系统中产生许多反应,其中之一是催产素释放激增(Levy et al., 1995)。因为正常的哺乳动物分娩过程会刺激母羊的阴道和子宫颈,VCS释放的催产素似乎有助于产生一个新的约2小时的社会联结窗口期,在此期间幼年动物的气味再次变得有吸引力。一项在婴儿中显示类似依恋反应的发现表明,在大鼠中,向幼鼠施用催产素可以使与母亲相关的气味更具吸引力(Nelson & Panksepp, 1996)。
虽然有蹄类动物似乎主要通过嗅觉机制建立联结,许多啮齿动物也是如此,但人类通过视觉、声音和触觉建立联结。我们目前还不知道通过这些感官建立的联结是否也通过催产素机制运作。然而,我们确实知道,触觉已被发现能够激活阿片类物质,这在社会联结中与催产素同样重要(见[第9章])。声音作为联结的来源,具有特别有趣的意义,因为母亲声音中引人入胜的语调可能是语言学习和我们对音乐热爱的入口(Panksepp, 2008b; Panksepp & Trevarthen, 2009)。听觉系统中的阿片类物质极其丰富,特别是在下丘的中脑下层(Panksepp & Bishop, 1981)。胎儿在出生前就开始整合子宫外的声音,甚至能识别母亲的声音(Busnel et al., 1992; Kisilevsky & Davies 2007),并可能对那些旋律性的语调——即妈妈语——产生印记,这最终将为语言习得开启全部潜能。
我们已经注意到,许多感官通道都能促进社会依恋。我们认为这部分是通过提升大脑中的催产素活性来实现的。我们还讨论了CARE敏感化,即接触非常幼小的动物可以逐渐唤起幼年大鼠和成年处女雌性大鼠的养育行为。在上一节中,我们注意到迷走神经宫颈刺激(VCS)会重新打开母羊的联结窗口,尽管我们还不知道VCS是否是促进催产素驱动联结的普遍机制。可能有许多外部刺激能够唤起CARE系统,但其中一个肯定应该得到进一步研究的是音乐。
有许多理由相信舒缓的音乐可以在大脑中释放催产素。农民们长期以来一直声称,他们的奶牛在听特定类型的音乐时会产更多奶(这一说法没有充分的文献证据),在我们关于分离痛苦的研究中,我们发现音乐可以减少幼年小鸡因分离引起的鸣叫(这是一个非常显著的效果)。在进行那些奇特的实验时,我们还发现,当我们给新生小鸡播放音乐时,我们可以唤起与直接向它们大脑中注入催产素时相同的明确行为特征。在出生后的第一周,接触音乐或催产素或后叶加压素的幼年小鸡表现出三种非常明显的高水平行为:(i)非常频繁的侧向摇头,(ii)打哈欠的频率大幅增加,暗示一种放松的心理状态,以及(iii)扇动翅膀的频率适度增加,暗示社交抑制的减少(Panksepp & Bernatzky, 2002)。事实上,最后一种行为,可能反映了类似自信的东西,如果在小型社会群体中而不是单独测试动物,这种行为会大幅增加(Panksepp, 1992)。自信是联结化学物质的几个关键功能之一——它们使动物感到社交舒适和自信,因为它们在大脑中提供了一个情感上安全的神经化学基础。正如我们将在下一章讨论的,脑内注入催产素在减少幼年动物的分离痛苦方面非常有效。
音乐唤起大脑催产素的力量可能是2004年国家地理拍摄的一部精彩半纪录片《哭泣骆驼的故事》核心的隐含”力量”。这部电影展示了蒙古戈壁沙漠的一个游牧牧羊人家庭如何诱导一只母骆驼在经历了非常艰难的、长达2天的分娩后与她的新生白色小骆驼建立联结。也许是因为分娩的长期压力远远超过了有蹄类动物通常短暂的联结窗口,母骆驼拒绝接受和哺育她的小骆驼。被拒绝的小骆驼可怜地哀鸣了很多天,通常与母亲保持相当远的距离。牧羊人不得不手工提供奶水。为了解决这一困境,游牧民进行了一场传统的”团聚仪式”,他们请来了一位著名的蒙古音乐家演奏动人的背景旋律,同时这个家庭的女主人对母骆驼唱摇篮曲,一边轻轻抚摸她的脖子和身体。在整个感人的跨物种互动过程中,小骆驼被引导去寻找母亲的乳头。母亲和幼儿被鼓励相互接触。这些牧羊人借助舒缓的音乐和触摸,试图再次打开联结窗口。所建立的深层情感和谐慢慢产生了魔力,不仅对人类观众产生了影响(这部电影获得了奥斯卡奖提名),也对母骆驼产生了影响。慢慢地,骆驼的心向小骆驼敞开,她接受了他进入持久的母婴联结的怀抱。这里有一个对人类的教训。
动物研究清楚地表明,一旦母亲在第一个后代出生后表现出称职和尽心的母性行为,她的母性能力就会永久性地提升。这是终身情绪学习的一个显著例子。这些母性经验累积的益处如何在神经系统中编码尚不清楚。但可以推测,这部分是由于底层CARE回路和相关记忆网络的持久变化,这些网络编码各种母性技能,以及以更自信和有效的方式应对世界的能力。在正式的恐惧测试中,如高架十字迷宫(见第5章),母性使动物更加勇敢。她们对食物位置的记忆也比未生育雌性更好,这可以通过在八臂放射迷宫中的表现来评估,在那里可以轻松量化找到食物的速度和模式(Kinsley & Lambert, 2006)。
尽管这种母性冲动强化的潜在神经生物学本质尚不清楚,但已经进行了大量重要研究来评估母性养育对后代体质强度的长期益处。从母亲那里获得丰富且持续的温柔关爱的婴儿得到了一份巨大的礼物。他们在余生中都会在情感和身体上受益。良好的母性养育对婴儿有益应该不足为奇,但支持这一事实的神经科学细节确实令人惊叹。
关键工作由麦吉尔大学Michael Meaney的实验室完成。Meaney及其同事评估了大鼠中母性触摸的数量,特别是肛门-生殖器舔舐——这是母鼠母性日常活动的重要组成部分——如何影响幼鼠日后的情感和认知能力(Meaney, 2001, 2010)。简而言之,Meaney及其同事发现,在被母亲充分舔舐和照料的幼鼠大脑中会出现许多终身益处。被充分舔舐的幼鼠长大后焦虑较少,对压力的抵抗力更强,并且在整个生命过程中更有能力表现出学习和其他适应性行为。这些效应伴随着大脑中许多可证实的变化,包括(i)应激激素减少(即促肾上腺皮质激素释放因子[CRF]和促肾上腺皮质激素[ACTH]),(ii)更多GABA受体位点,促进焦虑减少,以及(iii)更多谷氨酸和去甲肾上腺素受体,促进学习。在情感上,这些动物焦虑较少,在各种引发恐惧的情境中表现出更多活动和无畏,以及更好的学习和表现(Champagne et al., 2003; Zhang & Meaney, 2010)。
简而言之,充足的母性照料启动了一系列基因表达模式的表观遗传变化,使”被充分关爱”的动物更具韧性,对各种压力源具有强大的终身抵抗力。没有获得充分母性关爱的动物在情感上更脆弱,因此更容易被压力性生活事件所压垮。在下一章中,我们将更详细地讨论缺乏安全依恋如何影响人类婴儿的心智和大脑发育。
一些介导这些效应的神经遗传机制也通过研究得以揭示。动物的社会气质可以通过促进或减少催产素或加压素的基因表达来改变(Donaldson & Young, 2008)。这些变化的神经化学潮流不仅改变了初级过程的社会反应性,而且还渗透到整个神经系统,调节许多高级大脑过程,如社会记忆和认知(Ross & Young, 2009)。
尽管我们报道的重点主要集中在哺乳动物情绪的初级过程方面,但学习始终是这些系统在动物生活中表现自己的一个特征。因此,现实世界中的CARE会因某些经历而扩展,因其他经历而收窄。在一项开创性研究中,Lonstein和De Vries (2000)记录了这是如何发生在草原田鼠身上的,在那里父母通常共同承担养育责任,以及养育幼仔的倾向如何根据生活经验而变化。研究发现包括:(i)年轻的未生育雌性草原田鼠更具养育性;(ii)仅仅在断奶后让未生育雌性接触幼鼠2天就会增加日后的母性反应;(iii)与父母和兄弟姐妹一起长大的年轻未生育雌性表现出特别出色的养育行为;以及(iv)仅仅与父母双方一起长大就足以增加这些雌性的CARE动机;但是(v)这种提升只有在父母双方都在早期发育期间在场时才会出现。这些发现有力地说明了内在”家庭价值观”——最明显的那种——在增加年轻个体的养育动机方面的作用。在最近的研究中,双亲早期经历对草原田鼠各种关爱行为的促进作用,以及对大脑化学物质(如催产素和促肾上腺皮质激素应激系统)的持久有益影响已被证实(Ahern & Young, 2009)。
正如达尔文自然选择理论的洞见阐明了地球生命的故事一样,神经科学也丰富了我们对大自然通过漫长进化历程在哺乳动物大脑中构建的适应机制的理解。幼年动物达到生殖成熟并为其遗传后代提供养育的能力,与父母关爱的质量密切相关。母性和父性关爱(CARE)滋养大脑的方式,对于理解利他主义、同情心和共情如何成为可能具有重要意义。母性带来的对大脑和行为的终生益处是非凡的(Fleming et al., 1999; Kinsley et al., 2008)。
在未来的岁月里,我们无疑会对关爱系统(CARE system)了解更多。新的治疗方法可能会改变大脑的关爱化学机制和相关的社会情感系统。这些干预措施可以帮助父母更持续地体验养育情感,并更有效地展现支持性行为。促进催产素活性可能会增进那种接纳性的、积极的、亲社会的感受,这些感受能增强人们对更大情感开放能力的信心。事实上,在最近的一系列研究中发现,母亲的血浆催产素会随着与婴儿的大量亲密接触而增加(Feldman et al., 2010)。同样,鼻内催产素促进了父亲与孩子玩耍的质量(Naber et al., 2010)。许多研究这种神经肽的研究者开始怀疑,催产素可能在未来的心理治疗干预中发挥重要作用,旨在增强那些陷入个人怀疑和不安全感的人的积极社会情感(见Feifel et al., 2010; Panksepp, 2009c; Young & Wang, 2004)。随着脑科学的进步,许多其他可行的工具将变得可用,以帮助健康地重新整合和重新语境化困扰生活的情感和认知维度。
长期以来,人们都知道,当临床医生懂得如何以无条件的接纳、共情的敏感性和对来访者情感生活的充分关注来对待他们时,最有效的心理治疗就会发生。简而言之,有效的心理治疗师分享他们的关爱能力,以及调动积极情绪治愈力量的能力。这个教训不仅适用于那些专业重点是帮助治愈心灵的人,也适用于那些更多关注身体健康而非心理健康的忙碌临床医生,他们往往没有足够的时间关注来访者的情感问题(Goleman, 2006)。当然,充满爱的触碰不需要太多时间。但它确实需要持续性。
如果守护天使尚在
那为何我孤身一人
心怀忧伤,双眼空洞
曾经闪耀的光芒
已不复存在
在我如今被诅咒的生命中?
如果爱是一切,那是什么日食
让关爱黯淡,让一切坠落?
是什么心跳跳过
让打击倾盆而下
夺走了我的一切? . . .
—— Anesa Miller,《悲伤时光》(1995)
抑郁症的主要来源之一是因未解决的悲伤而吞噬心灵的心理痛苦。上述诗歌的前两节传达了深刻失去的痛苦——在这个案例中,是失去一个孩子的悲痛。这首诗的结尾写道:“如果时间能治愈一切,那就让它在我无法感知或看见的翅膀上流逝……陪伴和善意给予片刻的慰藉,就像天使,缓慢而安静地穿越悲伤的时光。”
这首诗写于1991年,作者是我(JP)的伴侣Anesa,写于我女儿Tiina和其他三名美丽的青少年去世几个月后。他们都被一个绰号”Suds”的酗酒者杀死,他的血液酒精浓度远远超过法定限度,大多数男人在这种情况下都会失去意识。他最近离婚了,整晚都在愤怒的情绪中喝酒,部分原因是他的妻子在那个耶稣受难日不让他见自己的孩子。他非常愤怒,醉得一塌糊涂就上了路。午夜时分,我接到医院的电话:“出了事故,可能我女儿牵涉其中。”我冲到那里。Tiina和两个朋友已经死了,一个正在垂死。还有一个幸存了下来,奇迹般地几乎没有受到身体伤害,因为撞击在他们车的侧面。
那天晚上剩下的时间我一个人;Anesa在华盛顿特区参加会议。她赶忙回家。我的悲伤和愤怒似乎无穷无尽……持续了很长时间。其他三个家庭也被摧毁了。一切都是因为一个鲁莽的醉汉像”从地狱飞出的蝙蝠”一样超速行驶,一个”牛仔”警察紧随其后,在夜晚不顾一切地追赶Suds沿着一条偏僻的乡间公路中间行驶,没有打开顶灯或警报器。至少,这是一个有爱心的家庭的证词,他们在事故发生的十字路口附近的家中吃耶稣受难日晚餐。执法机构试图压制不当警察行为的证据,医生拒绝公布康复中的醉汉的血液酒精结果,理由是”医患保密”,社区因此两极分化。有一段时间,我们的小大学城因”上层”的故意误导信息而分裂,社会结构的这种撕裂并未因当局的行动而轻易愈合。
那天晚上,我第一次像孩童时那样哭了。在很长一段时间里,我经历了深深的悲伤和抑郁,几乎看不到解决的希望。更令我困惑的是,这样的事情竟然发生在我身上——作为一名神经科学家,我正试图从实证角度阐明大脑中古老的分离痛苦机制,这是我们最早社会联结的主要情感来源之一。如果没有充满爱的伴侣和关心我的朋友,慢性抑郁肯定会降临。抗抑郁药物也部分缓解了我的黑暗时期(需要明智使用,即对症使用,而不是持续高剂量使用,否则可能会改变大脑神经化学平衡,造成额外的问题)。人们认为,哺乳动物大脑的分离痛苦机制打开了通向人类悲伤的大门(Freed & Mann, 2007),进而导致持续的抑郁绝望,尤其是当最初的分离痛苦源于生命早期失去父母时(Bowlby, 1960, 1980; Heim et al., 2004; Watt & Panksepp, 2009)。
我们在本章将要讨论的内容,在詹姆斯·桑德斯的戏剧《下次我会为你歌唱》(1962)中有一段精彩的描述:
每件事物背后都隐藏着某种特质,你可以相信也可以不相信,我们可以称之为悲伤。它总是在那里,就在表面之下,就在表象背后,有时几乎要显露出来,你能隐约看到它的形状,就像在平静的日子里,透过观赏池塘的水面,你有时能看到鲤鱼黑暗、粗大、非人性的轮廓缓慢滑过;这时你突然意识到,即使在阳光下水面闪闪发光时,即使你在逗弄那些古怪的鸭子和傲慢的天鹅时,鲤鱼一直都在下面,看不见。这种特质在等待时机。如果你确实瞥见了它,你可能会假装没注意到,或者突然转身在草地上和孩子们嬉戏,毫无理由地大笑。这种特质的名字就是悲伤。
本章探讨我们爱与玩耍能力的阴暗面。生活中有一个简单的事实,它带来深远的神经后果和心理健康影响:我们会依恋——我们会爱上——那些养育和善待我们的人。越来越清楚的是,母亲和其他充满爱的人是能够给予我们幸福生活的人(见第8章;Hrdy, 2009)。关爱情感和社会联结的进化,可能也是放大了我们哺乳动物悲伤能力的通道。人们认为,哺乳动物大脑的分离痛苦机制打开了通向人类悲伤的大门(Freed & Mann, 2007),进而导致持续的抑郁绝望,尤其是当最初的分离痛苦源于生命早期失去父母时(Bowlby, 1960, 1980; Heim et al., 2004; Watt & Panksepp, 2009)。
我们最早的社会联结,当它们坚固而安全时,会滋养我们一生的心理健康(Bowlby, 1980)。安全而温暖的母子关系是幸福生活的首要关键。对人类而言,这些拯救生命的联结在出生后开始缓慢形成,因为我们出生时在生理和心理上都不成熟——从某种意义上说,我们这些杂食动物和肉食动物都像早产儿一样出生。大多数食草动物在出生后不久就能跟着母亲奔跑,联结迅速形成并通过哺乳得以巩固——第一次吸吮尤为重要。相比之下,我们人类要到出生后半年左右才真正开始对纯粹的社会分离做出哭泣反应,而不仅仅是对身体不适——当母亲或其他主要照顾者将我们独自留在陌生地方时,表现出分离引发的痛苦发声(DVs)。玛丽·安斯沃思及其同事(Ainsworth, 1982; Ainsworth & Boston, 1952)首先研究了人类儿童的这种哭泣和抱怨。
一些动物行为学家已经注意到,幼小动物在与母亲分离时会发出痛苦发声,这是所有养过小鸡和小鸭的人都知道的事实。事实上,康拉德·洛伦茨(1935)演示了幼鹅如何依恋(对其”印记”)母亲,紧紧跟在她身后,如果仅分离几秒钟就会持续哭叫和寻找她。对这些恐慌性分离叫声的大脑机制的研究,可能是理解心理痛苦和悲伤的大脑机制最严谨的神经科学途径,这些机制似乎使许多生物体倾向于抑郁(MacDonald & Jensen-Campbell, 2011; Panksepp, 1981b, 1998a, 2010b, 2011a; Watt & Panksepp, 2009)。
如果没有稳定的社会照料和与所爱之人的安全联结,人类婴儿会憔悴并死去(Bowlby, 1953; Spitz & Wolf, 1946)。源于爱的依恋的温暖情感安全感——“安全基地”的主要机制——在2到3岁左右逐渐转化为更高形式的意识。在儿童期的前六年中,早期的社会丧失——过度的分离痛苦/悲伤——会使儿童对长期焦虑和不安全感敏感,通常预示着日后的抑郁症。另一方面,充满爱的社会依恋会增强大脑的积极情感力量,促进玩耍系统的健康运作(见下一章),这是帮助人类乃至所有哺乳动物成为我们现在这样复杂社会生物的基本心理力量。我们对针对自己的冷漠情感姿态反应强烈;任何暗示回避甚至更温和形式的社会排斥都会被体验为心理上的痛苦(Eisenberger, 2010)。随着高级心智功能的发展,即随着新皮层的发育编程,我们成为深刻关心社会网络质量的主体间生物(关于精彩的大众总结,见Goleman, 2006)。
我们刚刚开始理解社会丧失引起的深刻悲伤的大脑机制。对悲伤和悲痛期间招募的高级大脑机制的精彩成像研究(Freed et al., 2009)往往未能充分反映分离痛苦的古老皮层下机制(Panksepp, Herman et al., 1980),这一神经解剖学轨迹最终通过人类大脑成像得到证实,如图9.1所总结(Damasio et al., 2000; Panksepp, 2003a)。在可比的测试条件下,志愿者被要求从记忆中产生四种不同的情绪——即悲伤、快乐、愤怒和恐惧(见图12.1)——在强烈的悲伤感受中,悲伤系统表现出最清晰和最广泛的唤起。人类悲伤系统的总体解剖结构与在动物中绘制的介导分离叫声的系统相同。这个感受社会隔离刺痛的关键系统似乎在进化上源自介导身体疼痛情感强度的大脑系统。同样值得注意的是,这个古老的皮层下疼痛情感系统不同于循环到高级大脑区域以介导疼痛认知-辨别方面的系统。
如前所述,我们以前称这个系统为惊恐系统,因为当幼年动物被遗弃时,它们会经历一种特殊形式的惊恐焦虑——一种激动的惊慌状态。我们偏爱这个术语,因为有充分的理由怀疑惊恐发作部分源于这个主要情绪系统的过度唤起性(关于最近的概述,见Preter & Klein, 2008)。然而,许多读者发现这个标签令人困惑,可能是因为当成年人被剥夺陪伴时,他们倾向于感到孤独和悲伤,而不是像小孩子那样惊慌。当然,这只反映了成年人的三级过程沉思,他们有一生的方法来认知调整社会丧失,这是幼儿尚未学会的课程(关于人类孤独研究的当代讨论,见Cacioppo & Patrick, 2008)。尽管如此,由于我们使用惊恐一词的意图存在混淆,我们决定在本章中简单地将这个惊恐/悲伤系统称为悲伤系统。
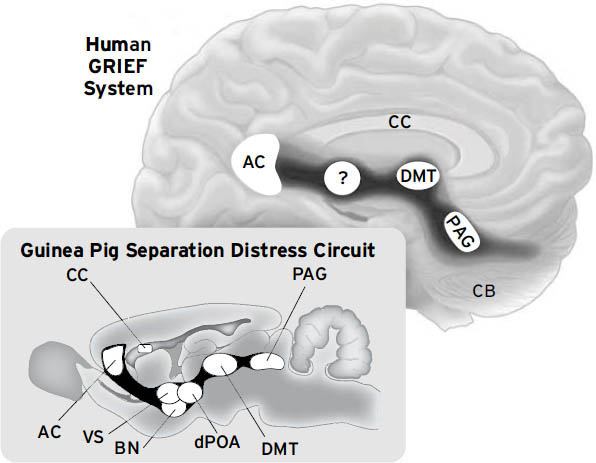
图9.1. 豚鼠大脑区域的示意图总结,在这些区域可以轻易唤起幼年豚鼠的社会分离诱导发声(如Barbara Herman的博士论文所绘制,鲍灵格林州立大学,1980)。在家养小鸡中观察到非常相似的解剖结构(Paul Bishop的博士论文,BGSU,1984)。当Antonio Damasio及其同事(2000)发表感到非常悲伤的人类PET扫描图像时,显示了非常相似的解剖结构,表明所有温血脊椎动物都共享介导悲伤系统的社会凝聚力。激活的区域位于前扣带回(AC)、背内侧丘脑(DMT)、导水管周围灰质(PAG),以及小脑最古老部分的区域(CB)。动物显示出非常相似的解剖结构,包括腹侧隔区(VS)、背侧视前区(dPOA)和终纹床核(BN),这些在人类PET图像上太小而无法准确识别。因此,在人类描述中有问号(该图首次发表于Panksepp, 2003;经美国科学促进会许可改编)。
这个系统有两个突出且相反的方面。首先,悲伤系统的唤起使我们感到失落和痛苦。但是,当痛苦得到缓解时——当我们再次被情感包裹在安全依恋中时——我们感到深深的安慰和安全感,可能是通过释放照料化学物质如内源性阿片类物质和催产素。在我们的一生中,当与我们爱和信任的人在一起时,我们会继续感到完整和自在。与我们的情感亲属在一起时,我们有”一切都好”的感觉。正是这种感觉,主要由上述社会神经化学物质介导,加强了社会联结,如前两章所讨论的。
我们研究团队在20世纪70年代中期对GRIEF系统及其神经化学控制机制的识别,为我们古老社交心智的情感机制提供了第一批神经科学见解。社会联结(social bonding)在某种程度上是一个成瘾过程变得清晰起来,它从一些同样促进麻醉品成瘾的大脑系统中获得情感强度。最近,很明显SEEKING的积极情感也对成年社交-性依恋有所贡献(Insel, 2003)。
虽然分离痛苦在大多数被研究的幼年哺乳动物中都有明显表现,在这些动物身上目睹这一现象可能令人动容,但当我们听到自己的孩子哭泣时最为令人心碎。任何经常去超市的母亲都能识别出在繁忙购物者的混乱中走失的孩子那种心理痛苦的尖叫。这些哭声牵动我们的心弦。与被拒绝要求零食的幼儿的抗议哭声不同,甚至与摔倒受伤的孩子的强烈痛苦哭声也不同,走失儿童的哭声有着无可置疑的恐慌音调和紧迫感。这种唤起的感觉似乎与我们的FEAR系统所能产生的焦虑关系不大。孩子不会像面对危险源那样躲藏或逃跑。它不会为了避免被捕食者注意而静止不动。相反,孩子往往会疯狂地四处奔跑(可能是SEEKING反应),哭泣并吸引注意力。在许多物种中,一旦母亲进入视线,或在短暂与母亲重聚后,哭声会变得更加强烈——这种现象称为”母性强化(maternal potentiation)“,这可能是该系统的次级、习得性反应,而非初级反应(即需要先前的社会联结)。那些不显示这种特定母性促进的物种,如大多数实验室小鼠和大鼠,大概没有真正的社会联结,即母亲和婴儿之间的独特纽带。它们只是表现出一种更普遍的社会奖赏过程。
许多研究人员可能会失望地得知,那些如此方便的实验室小鼠和大鼠并不是研究幼年社会联结机制的理想物种。它们有时表现出所谓的母性强化(Shair, 2007),但似乎它们的叫声是被任何雌性强化的,而不仅仅是幼崽自己的母亲。这可能只是一种普遍的多巴胺介导的SEEKING反应,而不是痛苦的放大。换句话说,目前还完全不清楚通过短暂的母性重聚偶尔观察到的叫声反弹是否特定于自己的母亲。因此,确定其他常用于模拟此类社会过程的动物的社会依恋特异性至关重要,以确保它们确实类似于人类的情况。这是我们花费大量时间评估另一种方便的小鼠大小物种——八齿鼠(Octadon degu)的效用的原因之一,该物种表现出丰富的社交行为,包括在生命的重叠阶段的真正联结、分离痛苦和社交游戏(Colonnello, et al, 2011)。
幼年动物在与母亲分离时为什么哭泣是相当明显的。这种童年痛苦具有适应价值,因为当非常幼小的动物感到害怕和孤独时,它们的哭声会提醒父母前来营救。想象一下开阔海面上的幼年海獭的惊人画面,它完全依赖母亲获取食物和照顾。当母亲潜入深处寻找食物时,她必须让婴儿独自待上许多分钟。在她不在时,幼崽会变得焦躁并持续哭泣。这些痛苦发声(DVs),一些科学家也称之为”隔离叫声(isolation calls)“,提醒母亲在哪里能找到她的宝宝。如果没有这种情感交流,在潜水时迷失方向的母海獭可能会与她的婴儿永久分离,婴儿就会永远失去。因此,婴儿的安全,实际上是生存,与将其与母亲连接在一起的音频-声音依恋纽带明确相关。
所有哺乳动物都是如此。在生命开始时,我们的依赖性是完全的,我们的生存依赖于与照顾我们的人建立的社会纽带。最重要的母婴情感纽带可以通过婴儿在母亲不在时通常发生的痛苦哭声轻松监测。事实上,这些痛苦发声是GRIEF唤起的主要标志。正是通过研究引发这种痛苦哭声的大脑网络,我们首次了解了可能导致抑郁的精神痛苦的解剖学和化学机制。同样,当幼儿得到不良照顾时——当被遗弃、忽视或虐待时——他们会经历由这些相同网络产生的持续不安全感和渴望,这可能会促进终身人格问题。而GRIEF的持续唤起可能促进慢性情绪障碍(Watt & Panksepp, 2009),可能是通过最终耗尽SEEKING系统的生命喜悦资源(Coenen, et al., 2011; Panksepp & Watt, 2011)。
悲伤(GRIEF)的能力并不局限于哺乳动物。鸟类的大脑深处也刻有社交需求的情感印记,这要归功于鸟类和哺乳动物共同的未知远古祖先。虽然我们对这些遥远的亲缘关系知之甚少,但我们确实知道,鸟类和哺乳动物的腹侧被盖区(DVs)起源于非常相似的大脑区域,并受相同的神经化学物质调节(Panksepp, Herman et al., 1980; Panksepp, Normansell, et al., 1988)。当这个悲伤系统被激活时,动物会强烈渴望寻求重聚。这些事实戏剧性地表明,悲伤是许多脊椎动物脑心(BrainMind)的一个古老情感系统,它介导一种可以很好地描述为”心理痛苦”的强烈情感。虽然这种情绪有着非常古老的根源,但它的影响在我们构建现代文明的方式中显而易见(Rifkin, 2009)。这个古老情感系统的功能对人类一生中的幸福感和痛苦感有着广泛的影响(关于该领域心理学研究的广度和深度的最新总结,见MacDonald & Jensen-Campbell, 2011)。
悲伤的感觉虽然痛苦,但对幼体的生存至关重要。此外,悲伤急性痛苦的缓解——从社交隔离的痛苦中解脱——可能告诉我们很多关于爱的本质。正如前面提到的,一些促进社交重聚引发积极情感的大脑化学物质类似于阿片类物质,人们很容易滥用这些药物,因为它们提供强大的情感慰藉感。当人类和动物相互接触并形成积极的社会纽带时,脑阿片类物质,加上催产素,可能还有脑催乳素,都会分泌到需要帮助的悲伤网络的化学受体上。
这些照料化学物质的作用是我们从与家人和朋友的温暖社会关系中获得情感慰藉的一个主要原因。哺乳动物和鸟类的亲属群体对彼此的陪伴”上瘾”,从而形成社会纽带,使它们能够生活在和谐的社会中(Panksepp, 1981a; Panksepp, Herman et al., 1980)。显然,生活在社会中增强了所有社会性物种的生存,因此我们有充分的理由相信,我们与他人共同生活的渴望——这种亲和力首先通过母子关系得到确认——是我们情感大脑的自然过程。这种感觉不是我们必须学习的东西,但我们确实必须学习与谁能以这种开放和”亲密”的方式相处(Reddy, 2008)。我们还必须学会建立培养我们本性中这种更好一面的社会结构(Rifkin, 2009)。
值得深思的是,在二十世纪中叶,行为科学家,甚至包括弗洛伊德这样的精神分析学家(至少在他最初的设想中),都认为社会纽带的形成纯粹是由于被良好喂养的结果,坦率地说。这个观点认为,幼儿爱他们的父母仅仅是因为父母提供营养、住所和温暖。据推测,这些”强化”经历在人类中是普遍的,因为人类婴儿在身体上不成熟,依赖父母满足他们对食物、水和温暖的需求。换句话说,行为主义者假设儿童通过与传统奖励的习得联想来爱他们的父母——仅仅因为父母提供维持生命的必需品。同样,行为主义者认为,如果照料者不满足孩子的身体需求,孩子就没有理由与照料者建立纽带。他们没有想到孩子或幼小动物除了满足身体需求之外,还有对社会依恋的内在需求。一些最臭名昭著的建议是由行为主义之父约翰·华生(John Watson)提供的,他在抚养自己的孩子时几乎不给予情感。他关于正确育儿的最著名建议是”永远不要拥抱和亲吻他们,永远不要让他们坐在你的腿上。如果必须的话,在他们说晚安时在额头上吻他们一次。早上和他们握手。如果他们在一项困难任务中做得非常出色,就拍拍他们的头”(摘自他的《婴幼儿心理护理》,1928年,该书在出版后几个月内就售出了超过十万册)。他自己的所有孩子都有严重的情感问题,可能是因为父母的冷漠,包括一个多次尝试自杀的女儿和一个成功自杀的儿子。
当勒内·斯皮茨(Rene Spitz)的经典研究揭示,在提供良好身体照料但缺乏情感的孤儿院长大的人类婴儿无法正常发育时,占主导地位的行为主义观点逐渐失去了阵地(Spitz & Wolf, 1946)。没有关爱的人际接触,许多婴儿过早死亡,而其他婴儿在成长过程中表现出严重的情感异常。近年来,我们在罗马尼亚独裁者齐奥塞斯库被推翻之前存在的孤儿院等地方再次观察到这一点。正如斯皮茨40年前在德国发现的那样,这样的婴儿在没有持续人类之爱的慰藉下会衰弱并无法茁壮成长。为了茁壮成长,婴儿显然需要情感滋养以及身体必需品。
与人类发育迟缓和情感发育障碍类似的症候群也存在于其他动物中。上世纪中叶的研究人员开始认识到,社会隔离会对幼年动物产生深刻的行为改变:伴随持续哭泣的焦躁不安,同时大量释放肾上腺应激激素。这些效应强烈表明存在基本的神经基质,它们旨在建立安全的社会纽带。从1960年代到1980年代,富有洞察力的精神分析学家、心理学家和生物学家开始断言,社会纽带源于我们对彼此深刻而天生的需求。精神分析学家约翰·鲍尔比(John Bowlby, 1960, 1980)在人类发展精神病理学领域的高度影响力的工作中强调,婴儿与父母之间糟糕的情感依恋会在儿童成长过程中引发各种严重的心理困难,这些困难可能持续存在并继续影响受影响个体的终生幸福感。
动物研究的结果与勒内·斯皮茨(Rene Spitz)的孤儿院观察一样可靠和明确。心理学家哈里·哈洛(Harry Harlow, 1958)关于隔离幼年恒河猴的著名研究,以及生物学家约翰·保罗·斯科特(John Paul Scott)关于羔羊和狗的工作,证明了当幼年动物与母亲分离时,它们会哭泣数小时,甚至数天(Scott & Fuller, 1998)。它们不再进食,被隔离的猴子陷入类似严重抑郁的绝望状态。缺乏母爱的幼猴会寻求任何可以找到的安慰,包括柔软的、无生命的”毛巾布妈妈”,而不是提供营养但不提供安慰的坚硬铁丝妈妈。当这种社会隔离持续几个月时,猴子在社会适应方面表现出终生的问题。即使幼猴与同龄伙伴而非母亲一起饲养,这些严重缺陷也只能部分逆转(Suomi, 2006)。这些影响是跨代的。当情感被剥夺的雌性长大并成为母亲时,发现了最可怕的终生问题之一。由于自身童年的剥夺,这些母亲无法对其后代做出充分的回应。例如,在隔离中长大的雌性恒河猴在与幼崽相处时往往胆怯且过度兴奋,母亲经常忽视或虐待幼崽(Harlow, 1958; Suomi, 2006)。
我们将尝试展示自鲍尔比首次发表关于依恋和发展的见解以来,我们在理解GRIEF(悲伤)的神经本质方面取得了多大进展。我们不再将婴儿对养育照料的需求简单地视为对身体照料需求的衍生物。勒内·斯皮茨及其同事(1946)的早期观察表明,仅仅照顾身体需求并不能防止因缺乏充满爱的人际接触而产生的”发育迟缓”综合征。我们终于理解,大脑中存在特定的网络产生我们对他人的需求。这些网络由特定的神经化学物质控制。这些化学物质的失衡会促进心理困扰,如果长期持续,则会导致疾病。我们还将讨论第一个社会关系的关键性质,即婴儿与其母亲或主要照料者之间的关系。我们将看到,无论好坏,这种关系在决定婴儿大脑发育方式方面具有决定性作用。
整个自然界充满了生命的普遍原则,这些原则被包裹在大量多样化的细节中。我们在CARE(关怀)章节中指出,不同物种的母亲以不同方式建立纽带。草食动物母亲在出生后很快与幼崽建立纽带,因为幼崽出生时非常成熟,从生命之初就可能走失。这些母亲快速与后代形成依恋,不仅是为了能够一起逃离捕食者,也是为了在后代走失时能够找到并找回它们;母亲还选择性地建立纽带,以便选择性地为自己的”孩子”提供资源。肉食动物母亲的后代出生时非常不成熟,眼睛和耳朵都尚未睁开——还有其他晚成性物种如猫、狗和人类——有更大(更长)的纽带建立窗口期。这些婴儿在生命早期不容易走失,所以母亲有充足的时间与婴儿建立纽带,而婴儿通常直到接近可以独自走失的年龄时才开始与母亲形成特定的纽带。因此,人类婴儿只有在出生后第一年的下半年才表现出与母亲的强烈纽带。在此之前,婴儿似乎对任何持续照顾他们的人都很满意。
因此,如前所述,科学家们必须关注一些他们青睐的”人工”物种,如实验室大鼠和小鼠,这些动物通常已经为研究目的繁育了数百代,可能只保留有残留的悲伤系统(GRIEF systems)(Panksepp et al., 1992; Panksepp, 2003b)。这也是它们非常适合行为研究的原因之一,这类研究需要”未受污染”的动物(尤其是在行为主义时代),这些动物单独饲养在无菌笼中时也能很好地存活。与幼年豚鼠、小鸡和灵长类动物不同,年幼的实验室大鼠在单独饲养时不会因抑郁性绝望而消瘦。当然,它们也不像许多爬行动物那样在社交上冷漠。它们确实非常喜欢社交陪伴,如果长期单独饲养,它们会有些抑郁。正如我们将在下一章看到的,它们特别渴望玩耍等社交互动,而社会隔离会强烈增强玩耍行为(在强烈依恋的物种中,隔离只会促进依偎和黏附)。因此,如果有选择,幼年大鼠总是会选择与友好的同伴在一起。事实上,在条件性位置偏好(conditioned place preference)研究中,社交陪伴作为一种强大的奖励,这种效应在雌性中比雄性更强(Panksepp et al., 1997)。
重要的是要认识到,幼年大鼠和小鼠并不表现出真正的分离叫声,尽管当它们单独留在巢外时,确实会发出一些微小的超声波”咔嗒”声。这些不是真正的分离叫声,因为这些幼崽太不成熟,无法自己走失。这可能只是反映了身体不适。在两周龄之前,幼年大鼠无法自行进行适当的体温调节,它们需要向母亲发出信号,表明自己被拖出巢穴并且正在变冷。这些动物没有功能性的分离痛苦和社会依恋系统。事实上,当它们长到足以走失的年龄,大约15天大时,它们的身体不适叫声会消失,但不会被社交分离叫声所取代。尽管在非常小的时候(例如,特别是在它们约14天睁眼之前的一周)反复经历持续早期隔离的幼年实验室大鼠确实表现出长期的抑郁型行为和大脑变化,但这些效应可能主要是由于一般性压力(寒冷和没有母亲舔舐),而不是特别由于社会隔离痛苦(Heim & Nemeroff, 1999)。如果使用实验室啮齿动物来尝试模拟人类依恋和抑郁过程,这是一个大问题。
显然,幼年实验室大鼠的社交动机存在某些特殊之处:它们似乎不需要特定的同伴——甚至不需要它们的母亲。对它们来说,任何其他母鼠似乎都能提供足够的安慰来消除痛苦指标。这与我们人类物种截然不同。一旦形成特定的社会纽带,即使周围有很多人,人类儿童也可能会哭泣,就像超市的例子一样。只有孩子的妈妈或熟悉的照顾者才能提供完全的安慰。许多其他物种也会形成这类特定的社会纽带。但实验室大鼠和小鼠的情况并非如此。它们所需要的只是任何其他友好动物的存在就能显得情感满足,至少在喂食时间之前是这样。显然,实验室大鼠和小鼠的分离痛苦反应相对较弱,目前没有证据表明它们像许多其他啮齿动物一样具有特定的母性社会依恋系统(见Colonnello, et al., 2011)。也许它们的社交欲望主要通过通用的寻求系统(SEEKING system)运作,该系统也有助于调节成年社会纽带(见第7章)。越来越清楚的是,成年社会性-性纽带是由大脑多巴胺介导的寻求行为促进的(Insel, 2003)。而婴儿-母亲纽带的情况并非如此,其中内源性阿片类物质和催产素至关重要(Nelson & Panksepp, 1998)。
我们通常从外部事件的角度来思考痛苦和幸福:我们感到痛苦是因为有人背叛了我们,我们感到快乐是因为朋友给予了关心,甚至是充满感情的支持。但悲伤系统,像所有初级加工情绪系统一样,最初是”无对象的”——在生命早期有一个时间点,它可以很容易地与任何支持和关心的个体建立联系。即使是一个虐待性的个体也比没有个体要好。当那个个体不在身边时——当幼年动物被完全单独留下时——它们的痛苦会变得强烈,除非被照顾者救助,否则它们会长时间哭泣。这些模式为什么以及如何展开?
最具信息量的神经科学证据来自于对一种基本行为测量的详细分析:幼年动物在社会隔离时引发的分离叫声。我们对这个脑系统了解很多,因为我们可以通过电刺激或药物刺激特定的脑区,从而唤起分离痛苦。对几种动物的研究揭示了情绪系统的神经解剖学位置,该系统介导分离叫声(DVs)以及由社会排斥和失去引起的独特社会感受。在20世纪70年代末,我们确定了可以通过电刺激唤起分离叫声的特定脑区:特别是导水管周围灰质(PAG)和周围的中脑区域、背内侧丘脑、腹侧隔区、背侧视前区以及终纹床核(bed nucleus of stria terminalis)的位置(Panksepp, Normansell, et al., 1988)。在高等物种中,分离叫声也可以通过刺激前扣带回以及散布在杏仁核和下丘脑的一些位置来唤起。换句话说,像其他情绪原型一样,悲伤系统(GRIEF system)由一个广泛的情绪网络组成,主要集中在皮层思维帽下方的古老内侧脑区。对人类成年人悲伤的脑成像研究突出了非常相似的脑区,同时伴随着脑阿片活性的降低(Zubieta et al., 2003)。
有充分的理由相信悲伤系统是从原始的脑干疼痛网络进化而来的。一个原因是身体疼痛和分离叫声都很容易被阿片类药物缓解。分离痛苦回路集中在更古老的内侧脑区,如导水管周围灰质,它传递疼痛的深层情感体验,而不是由大脑更高区域介导的疼痛认知方面。例如,强烈的悲伤反应(分离叫声)以及恐惧和愤怒反应是由导水管周围灰质的背侧部分产生的,这也是一个产生身体疼痛大部分情感强度的脑区。因此,悲伤的心理痛苦可能与身体疼痛的古老情感信息有着强大的进化联系。这就是进化的运作方式,通过使用预先存在的解决方案来制造新的生存工具。
显然,基于悲伤的社会需求系统存在于广泛的脊椎动物物种中,包括我们人类。相关的脑刺激研究以及涉及内源性阿片类物质的神经化学分析首先在豚鼠中进行(Herman & Panksepp, 1981),然后在进化上非常遥远的物种——家鸡中得到重复验证(Panksepp et al., 1988)。现在已经在其他物种,特别是灵长类动物(Jürgens, 2002)中进行了足够的研究,使我们确信这种回路是所有具有强烈动机形成持久社会联结的脊椎动物的普遍属性。然而,动物对这个系统的依赖在整个生命周期中差异很大。幼年动物非常依赖它,但随着成熟,该系统的反应性会降低,部分原因是性类固醇的抑制作用,这使动物转向成年形式的社会性满足。
人们一定想知道随着幼年动物的成熟,这些系统会发生什么。许多成年人,尤其是男性,很少哭泣。有些男人在他们成年生活的大部分时间里都不会哭。这些痛苦回路发生了什么?它们萎缩消失了吗?它们还在那里但因为不使用或其他内在神经生物学原因而变得非常不敏感了吗?对成年豚鼠的研究清楚地表明这些回路仍然存在:对大脑中正确位置的针对性电刺激仍然可以让成年雄性像婴儿一样哭泣(Panksepp & Miller, 1996)。
然而,随着年龄的增长,这些回路的反应性远不如我们婴儿时期。我们怎么知道这一点?当人们使用局部脑刺激人为激活豚鼠的这些系统时,随着动物的成熟,需要越来越多的电流才能引发哭泣(Panksepp & Miller, 1996)。这个悲伤系统的敏感性随着动物经历青春期而逐渐降低,并且在雄性中比在雌性中变得更不敏感。这表明青春期性激素水平的增加起着关键作用。事实上,当年轻的雄性和雌性豚鼠被阉割(移除性腺和卵巢)时,分离痛苦系统的敏感性不会像完整动物那样降低太多(Sahley & Panksepp, 未发表数据, 1986)。在青春期,具有完整性腺的雄性在接受脑刺激时比完整的雌性哭得更少。结论很明确:“大男孩不哭”,不仅仅是因为他们被教导不要哭。他们不太可能哭是因为他们成熟的性腺在青春期分泌大量睾酮。这是否是男孩通常变得咄咄逼人、比女孩更缺乏社会敏感性和同理心的关键原因?也许是。最近的研究还表明,给女性使用睾酮往往会迅速引发更类似男性的心理态度(Bos, et al., 2011)。
同样,成年人比儿童更不容易哭泣,但他们仍然容易受到失去所爱之人所带来的悲伤和痛苦的影响。在人类研究中,对动物中包含GRIEF回路的大脑区域进行强烈的电刺激,可以立即使人进入抑郁绝望的状态,而当刺激停止时,这种状态会迅速消失(Bejjani et al., 1999)。目前,刺激前扣带回等大脑区域是治疗对抗抑郁药物无效的抑郁症患者的一种方法(Mayberg, 2009)。这类治疗的成功可能是由于破坏(抑制)了GRIEF的某些高级大脑机制,但这还需要未来的工作来确定。事实上,为帮助难治性抑郁症患者感觉好转而进行的各种脑损伤治疗,其效果可能都是通过损害GRIEF的大脑网络,或许还通过促进大脑SEEKING系统的活动来实现的,而SEEKING系统通常会在我们的心理生活中促进热情和积极活力的感受(Coenen, et al., 2011; Schoene-Bake et al., 2010)。
GRIEF系统的唤起感觉糟糕,而能够降低其唤起性的药物的效果则感觉良好。我们对这个系统了解得足够多,可以得出结论:人为操纵其神经化学物质可以增加或减少痛苦和社会动机。当特定的大脑化学物质处于低水平时,婴儿会发出分离呼叫(DVs),成年人会感到孤独。另一方面,当这些化学物质处于高水平时,婴儿会感到快乐,成年人会感到愉快和自足。在日常生活中,当婴儿得到温柔的照顾,当成年人得到家人和朋友的支持时,就会出现这些满足的反应。因此,积极的社会联结,或者说我们拥有”安全基地”的感觉,伴随着高水平的社会依恋化学物质。
特别是三种神经肽大脑化学物质已被证明能够强烈减少GRIEF(同时还有大量次要参与者)。这些神经肽中的第一种,也许是最强的一种,是内源性阿片类物质(opioids),它们的药理学形式(如吗啡和海洛因)可能具有很强的成瘾性。另外两种能够强烈减少分离痛苦的物质,催产素(oxytocin)和催乳素(prolactin),我们已经在介导CARE中遇到过,它们是重要的参与者。简而言之,如果处于痛苦中的婴儿体内的脑阿片类物质、催产素或催乳素升高,DVs就会减少,婴儿会放松并表现出舒适的迹象,这些迹象通常在享受母亲抚慰时表现出来(Panksepp, 1998a)。
从1970年代中期开始的实验,通过监测与正常社会环境(通常是母亲)短暂分离的幼年动物(幼犬、豚鼠和小鸡)表现出的DVs频率来监测社会痛苦(Herman & Panksepp, 1978; Panksepp, Herman et al., 1978; Panksepp, Vilberg et al., 1978)。通过使用分离痛苦呼叫作为社会联结所必需的潜在情绪的客观指标(Panksepp, Herman et al., 1980),研究发现,所有激活三种主要阿片受体之一——μ(mu)受体的大脑化学物质和药物,在减少所有三种测试物种的DVs方面都非常有效。这些效果是由于改变了大脑GRIEF网络的敏感性(Herman & Panksepp, 1981)。这些效果随后在啮齿动物和灵长类动物中得到了重复和扩展(Kalin et al., 1988; Kehoe & Blass, 1986; Keverne et al., 1997; Newman, 1988)。
这些发现表明阿片成瘾与社会依恋机制之间存在关系。这可能有助于解释为什么这么多人,尤其是孤独的人——那些有太多心理痛苦的人——会对麻醉品上瘾(Maté, 2008; Panksepp, 1981a)。刺激其他称为δ(delta)受体的阿片受体具有类似但较弱的效果。第三种阿片类物质,强啡肽(dynorphin),通过κ(kappa)受体运作,产生完全不同类型的感觉。产生的感觉通常非常令人厌恶,带有迷失方向、解离的感觉,使一些人感觉好像正在失去理智。现在看来,强啡肽在抑郁的动物和人类中可能变得过度活跃(Land et al., 2008; Watt & Panksepp, 2009)。
当麻醉品成瘾依赖于这样的系统并且是社会的祸害时,大脑中内源性阿片系统的存在可能服务于什么进化目的呢?一个初步线索是,阿片成瘾的动态与积极的社会关系之间存在显著的相似性。最初,药物成瘾始于一个时期,在这个时期,药物会产生一种强烈的欣快愉悦或情绪缓解反应,吸毒者开始珍视并强烈渴望这种感觉,因为当这些药物从身体中排出时,这种感觉就会消失。当药物离开系统时,会有一种不满足的、甚至是痛苦的情感残留(“对立过程”)。这导致反复使用,在某些人身上最终导致药物滥用。长期使用麻醉品通常会导致药物耐受性(tolerance),在此期间需要越来越多的药物才能产生现在熟悉和期望的积极感觉。现在,个体依赖药物来维持任何正常的感觉。然而,如果在产生耐受性后使成瘾者被剥夺药物,他或她将经历一段戒断期,伴随着不快的感觉和痛苦,这种感觉与失去朋友时的悲伤感觉并无太大差别(总结见图8.2)。
社会关系遵循类似的轨迹。最初是极具吸引力的社会联结感,随后随着人们习惯于对方而逐渐减弱,这可能类似于阿片类耐受。然而,如果随后关系受到威胁或终止,人们会经历一段分离痛苦期。这种痛苦的性质类似于幼儿因母婴分离引发的恐慌,以及成人的悲伤或哀痛。因此,最初假设社会联结是由内源性阿片类物质引发的愉悦感所介导的(Panksepp, Herman et al., 1978)。研究认为,大脑中丰富的阿片类物质产生温暖联结的社会关系中的满足感。阿片类物质的突然匮乏可能会引发我们在孤立或丧失亲人时感受到的恐慌和悲伤。
这一假设在历史上得到了支持,因为精神病学曾将阿片类药物用作抗抑郁药。实际上,在20世纪50年代早期现代情绪障碍精神药理学时代开始之前,它们是医生可用的唯一有效的精神科药物。尽管使用它们常常导致成瘾,但毫无疑问它们能让不快乐的患者感觉更好(Tenore, 2008)。此外,关于阿片类成瘾的心理学文献表明,阿片类药物的情绪效应与人们从支持性社会联结中获得的安全感非常相似。
许多药理学研究现已证实了社会情感和社会联结的阿片类假设——这是首个获得实证支持的社会依恋神经生物学理论(Panksepp, Herman et al., 1980),甚至已扩展到鸟类美妙的歌唱和社会联结(Riters, 2011)。这一观点还扩展到我们从许多其他感官情感中获得的愉悦,例如进食的愉悦(Avena et al., 2008),以及可能伴随长跑而来的兴奋感(Strassman et al., 1989)。推测是这些不同的效应源于许多不同脑区的阿片类活动。然而,刺激μ-阿片受体的化学物质显著减少分离痛苦的发现充满了精神病学意义。例如,临床抑郁可能在很大程度上反映了大脑中的愉悦缺陷。
令人欣慰的是,最近的脑成像研究表明,人类的悲伤和相关社会过程是由这些相同的脑区介导的(Damasio et al., 2000; Lorberbaum et al., 2002; Swain et al., 2007),并且人类的悲伤和抑郁伴随着低水平的脑阿片类物质(Kennedy et al., 2006; Zubieta et al., 2003)。特别重要的是要注意,脑阿片类物质的缺乏可能导致几种精神障碍。除了抑郁感外,越来越清楚的是,通过给予减少大脑阿片类活动的受体阻滞剂会促进恐慌发作的神经生理学(Preter, et al., 2011)。据推测,抑郁的人和缺乏足够社会支持的人大脑中舒缓的社会情感分子水平较低,使他们更容易滥用成瘾性药物。积极的社会活动,包括母性(Ferris et al., 2005),倾向于降低可卡因和阿片类成瘾的可能性,至少在动物模型中是如此。这可能是由于从积极的社会互动中获得的自然但在情感上相当的满足感。我们现在还知道,各种积极的社会互动,如玩耍,会导致大脑中内源性阿片类物质的释放,这可能对减少成瘾行为和其他精神问题的自然方式有进一步的意义(见第10章)。
内源性阿片类物质与成瘾性阿片类药物如此相似,它们介导社会关系这一事实表明,哺乳动物,特别是完全依赖他人的幼崽,实际上是对社会关系成瘾的。就其本质而言,哺乳动物和其他脊椎动物(如鸟类)通过阿片类介导的社会依赖维持家庭生活,当人们通过药理学满足这些系统时,这种依赖就会变成成瘾。因此,有些人更容易对阿片类药物成瘾的一个原因可能是他们没有从社会生活中获得足够的积极满足。许多人可能在自我治疗,因为他们有慢性的”心理痛苦”感,这种痛苦部分源于长期活跃的悲伤系统(GRIEF systems)。他们已经了解到,通过立即摄入分子而不是通过发展积极的社会关系,他们可以更快地获得缓解(Insel, 2003; Panksepp, 1981a)。
我们现在知道,各种舒适的环境刺激可以无条件地导致内源性阿片类物质β-内啡肽和其他舒适的脑化学物质的分泌。身体温暖、熟悉的母亲气味、舒缓的声音、吮吸,甚至甜的糖水都会使许多幼年哺乳动物(包括人类)的哭声减弱。例如,大鼠幼崽在吮吸后会表现出刻板的伸展反应和轻微的疼痛不敏感性(Smotherman & Robinson, 1992)。在人类婴儿中也观察到类似的镇痛效应(Gray et al., 2002)。这表明乳汁的注入诱导放松和舒适,这是阿片类药物的典型反应。如果在乳汁注入前给予阿片受体阻滞剂,这些满足的反应就会被抑制。
与其他情绪一样,这种原始加工情绪系统的影响会随着人类和动物的成熟而通过学习得到扩展。阿片类物质的释放无疑也会受到条件反射和各种学习经验的影响。例如,人们更容易去爱那些与自己以前爱过的人相似的人。即使是幼年大鼠也会对与母亲相关的气味表现出偏好(Nelson & Panksepp, 1998)。原始情绪系统是我们最基本的价值编码工具之一,每一种都通过亿万年的大脑进化形成了独特的效价(valenced)。这些系统产生的进化经验指导着所有其他心理装置的构建——即次级和三级认知策略,我们的大脑通过这些策略在表观遗传适应之上层层叠加适应。
我们可以很容易地想象,从生命的最早阶段开始,条件性次级学习是如何发生的,将阿片类物质的释放与各种条件刺激联系起来。例如,如果母亲在给婴儿喂奶时经常播放音乐,那么单独听到音乐声可能最终会导致婴儿大脑中阿片类物质的释放,从而产生舒缓的情感。因此,音乐作为一种条件刺激,可能在婴儿的神经发育过程中获得持久的、情感驱动的意义。当然,某些类型的舒缓音乐,作为”触摸”的特殊形式(听觉是一种皮肤振动感觉),可能具有释放阿片类物质的内在能力,而较低级的听觉中继站,如下丘(inferior colliculi),富含阿片受体(Panksepp & Bishop, 1981)。
稍加思考就会发现,条件刺激、因应这些刺激而产生的认知策略,以及在一生过程中这些态度之间发展起来的关系,在数量、范围和复杂性上都是惊人的。这些力量对人格发展和认知成熟具有重要意义。由于它们的长期影响,我们往往别无选择,只能遵循从大脑古老区域涌现出来的情感指令。正如生活的许多方面一样,情绪引领着方向。
随着良好依恋的儿童成长为成年人,他们学习社交技能以保持与朋友和亲人的亲密关系。这种学习过程必然涉及高阶社会情感的发展——从羞耻和害羞到共情——它们在这些关系中发挥作用。例如,如果一个孩子在他人眼中显得愚蠢,她可能会感到羞愧。如果她不确定自己喜欢的人会有什么反应,她可能会感到害羞,当然,孩子渴望与所爱之人保持亲密的愿望会促进共情。孩子还会发展出一些技能,使他们能够应对不可避免的孤独时期——也许通过转移注意力,或者通过进行令人满足的幻想和游戏。所有这些策略都与我们的社交参与水平和需求相关。从神经科学的角度来看,所有这些策略都反映了对维持情感平衡这一基本需求的反应,在某些情况下是对大脑中阿片类物质分泌的调节(Panksepp, Siviy et al., 1985)。
依恋不良的儿童往往无法发展这些能力,作为成年人,如果他们不得不独立生活,很容易变得抑郁甚至恐慌。即使有社交网络可用,关系也可能因其碎片化和常常带有攻击性的特质而受损。当这种负面倾向在人格发展中出现时,各种心理和药物治疗可以有效促进情感稳态([第12章])。
我们的爱恋依恋能力也可能采取许多不那么明显的社交形式,从我们已经看到的对音乐的热爱,到毒品成瘾的堕落。甚至宗教的情感吸引力和力量很可能也基于社会依恋系统,无疑还辅以大脑的其他社会情感(Thandeka, 2005, 2009)。考虑这样一个事实:社会隔离的幼猴那种弯腰、自我抱持的姿势与祈祷姿势基本相似——一种绝望和恳求的身体姿势,头低垂,手臂和双手紧握,表达了对慰藉的最原始需求。在人类中,这常常转化为对来自更高力量的关怀(CARE)的认知渴望。另一种常见的礼拜姿势,如神职人员布道时所见,双臂优雅地向天空举起,类似于婴儿向上伸手寻求照顾者如母亲的急切寻求姿势。
阿片类物质的分泌对触摸的舒缓效果特别敏感。我们都知道,可以通过抚摸来安慰家养动物。客观研究这种效果的一个简单方法是监测幼年动物在被抱起或未被抱起时的哭声。当然,效果是显著的。动物在被轻柔触摸时会迅速停止哭泣。有令人信服的证据表明,这种接触安慰部分是由大脑阿片系统的激活所介导的。例如,因与群体分离而焦躁不安的幼雏,在被人类抱起时会迅速安静下来,甚至舒适地闭上眼睛([图9.2])。阿片受体阻断剂会降低这种接触的有效性,增加使幼雏平静下来所需的时间。然而,即使通过纳曲酮(naltrexone)或纳洛酮(naloxone)(阻断阿片类物质效应的化学物质)完全阻断阿片系统,以这种方式被轻柔抱起的社会隔离鸟类最终也会安静下来,哭声远少于未被抱起的对照组鸟类。显然,除了阿片类物质之外,还有其他神经化学物质对接触安慰的感觉有所贡献。
图9.2. 当新生雏鸟被人类双手轻轻握住时,会表现出舒适反应,包括停止发声和闭眼。这些效应会被阿片受体阻断剂纳曲酮减弱,并被低剂量阿片类药物增强(绘图:Lonnie Rosenberg;摘自Panksepp, 1998a;经牛津大学出版社许可重新发表)。
触摸可以释放大脑中的阿片类物质,这一事实也在灵长类动物中得到了证实(Keverne et al., 1989)。事实上,给予阿片受体阻断剂纳洛酮往往会增加灵长类动物的梳理行为,这可能是为了对抗大脑中阿片类物质减少所带来的负面情感。灵长类动物相互梳理是为了获得社交舒适感。用神经科学术语来说,这意味着梳理行为是有奖赏性的,至少部分原因是它促进了大脑中阿片类物质的分泌。同样地,服用阿片类药物会降低被触摸的欲望,这可能是因为阿片类药物在动物大脑中诱发了一种阿片类自我满足感。事实上,上述研究中地位最高的猴子不希望被其他猴子梳理,而她的基线大脑阿片水平最高。她不需要被梳理,但她非常愿意梳理其他猴子。
在发现内源性阿片类物质在调节悲伤系统中的作用后不久,人们发现催产素(oxytocin)和催乳素(prolactin)是同样强效的该系统抑制剂,并且它们能够加强婴儿与母亲之间的社会纽带。我们在上文中提到,催产素和催乳素,以及阿片类物质,都能平息幼年动物的痛苦发声(DVs)。然而,仍需要进行大量研究。特别是我们需要更多地了解这些化学物质影响人类悲伤系统的机制。尽管现在普遍认为社会情感过程受到阿片类物质、催产素和催乳素的调节,但我们还不完全理解这些系统是如何相互作用的。
关于大脑催乳素在情感调节中的作用,我们所知甚少。然而,催产素已经受到了广泛研究(Insel, 2010),并已证明在平息痛苦发声和产生社会依恋方面发挥作用。此外,催产素作用的证据可以通过检查动物在生命周期不同时期的大脑来找到,当时它们表现出不同的社交倾向。在幼年大鼠的大脑中,当社会纽带对生存至关重要时,人们会在悲伤系统的大脑结构中发现高水平的催产素受体。在成年期,当社会纽带对生存不那么关键时,这些系统中的催产素受体密度较低。当动物年幼时,催产素可能在它们主要以初级过程为主的社交和情感生活中发挥更关键的作用,而不是在它们身体成熟并拥有丰富的认知备用机制和策略来维持情感平衡时。不过,催产素似乎会在一生中通过舒适的社交触摸而释放。例如,已知在腹部被抚摸的动物会向血液循环中释放更多催产素,接受按摩的人类也是如此(Uvnäs-Moberg, 1998)。
在具有不同社交气质的物种中,人们也会发现催产素受体的密度不同。例如,社会纽带强烈的草原田鼠(prairie vole)的大脑具有密集的催产素受体分布,更类似于幼年动物的大脑。另一方面,山地田鼠(montane voles)倾向于独居,除非有性动机,它们不仅总体催产素受体较少,而且这些受体在大脑中的分布也不同。在独居和成对结合的猴子以及野生小鼠中也观察到类似的催产素分布差异模式(Donaldson & Young, 2008; Ross & Young, 2009)。
尽管催产素研究近年来已成为热门话题(Carter, 1998; Insel, 2010; Nelson & Panksepp, 1998; Ross & Young, 2009),但这种化学物质如何产生情感变化绝非清晰。如前一章所述,催产素可以增强内源性阿片类物质的效应。动物通常会对阿片类物质产生习惯化(耐受性),这意味着一段时间后,药物会失去一些效力。这就是为什么成瘾者需要越来越多的药物,以及为什么新友谊最初的欣快感可能会随着时间的推移淡化为更平静的陪伴。催产素可以减少这种阿片类耐受性,使阿片类物质在更长时间内保持更强的效力。也许催产素增强了诸如β-内啡肽等阿片类物质的活性(Kovács et al., 1998)。如果是这样,催产素所能产生的情感舒适感——信任和安全感——很可能最终会被证明是由阿片类物质介导的。
催产素和阿片敏感性之间的这种关系对于大脑充满催产素的新手妈妈来说可能尤其重要。除了提供母性情绪外,催产素可能会延长阿片类物质的效应,这可能是母性情绪如此强烈和持久的原因之一。同样的原理可能在社会纽带中发挥作用。催产素可能会增加内源性阿片类物质的效力,使它们提供更强烈的舒适感——特别是对仍然完全依赖照顾的幼崽。
社会纽带与内源性阿片类物质之间的实质性联系促使Panksepp考虑,自闭症的社交孤立症状可能是由高水平的内源性阿片类物质引起和/或加剧的。事实上,近一半社交孤僻的自闭症儿童在低剂量纳曲酮治疗后表现出亲社会的临床改善(Panksepp, Lensing et al., 1991)。虽然过量的内源性阿片类物质不太可能是自闭症的主要原因,但在某些病例中非典型阿片类物质可能是一个因素(Bouvard et al., 1995)。这项关于自闭症的研究标志着为数不多的一次,动物大脑情绪系统的基础研究产生了有效的精神病学干预措施。然而,这并未成为标准治疗方法,因为从未进行过足够广泛的双盲安慰剂对照研究,尽管许多研究已经证实了最初的结果(Green & Hollander, 2010; Kolmen et al., 1997)。
最近,催产素在自闭症和儿童发展文献中占据了重要地位(Bartz & Hollander, 2008; Carter, 2007; Insel, 2010; Yamasue et al., 2009)。研究观察到自闭症儿童的血浆催产素水平低于正常水平,鼻内给予该神经肽后有一些适度的社交益处(Green & Hollander, 2010; Heinrichs et al., 2009; Rossignol, 2009)。然而,正如此类研究领域常见的那样,必然会出现无法重复和扩展这些发现的情况(Tansey et al., 2010)。自闭症是多种大脑-身体问题的复杂组合,有许多遗传因素,目前没有可靠的方法来区分众多亚型。当前的自闭症流行——大约每120个儿童中就有1个可能被诊断为自闭症谱系障碍——表明在我们的后现代社会或物理环境中正在发生某些事情,促使儿童无法在社交方面茁壮成长。正如我们在下一章强调的,一个大问题可能是我们不再允许儿童成为儿童——让强健的身体游戏成为他们日常社交饮食的一部分。
我们已经看到,内源性阿片类物质和催产素的缺乏可能导致非常年幼的个体产生孤独感甚至恐慌感。当悲伤系统被唤起时,其他大脑化学物质变得更加活跃,特别是压力神经肽——促肾上腺皮质激素释放因子(CRF)和谷氨酸,一种参与每种情绪反应的兴奋性神经递质。事实上,当这些化学物质被注入动物大脑时,它们非常强烈地促进分离叫声(Panksepp, 1998a; Panksepp & Bekkedal, 1997)。CRF产生正常的叫声(见图9.3),而谷氨酸激活后的发声在声学上往往是异常的(Normansell & Panksepp, 2011)。CRF是参与经典压力反应的激素,它激活垂体-肾上腺系统。压力激活下丘脑室旁核(PVN)中的神经元,该核含有大量CRF神经元。
这些PVN轴突投射下行至垂体前叶,导致促肾上腺皮质激素(ACTH)释放到血液中。ACTH是刺激肾上腺皮质的触发因素,肾上腺皮质位于肾脏正上方,释放皮质醇激素。这种类固醇帮助身体以能够应对多种压力情况的方式使用能量,包括分离痛苦。目前尚不清楚皮质醇如何帮助动物应对分离痛苦,但一个可能的选择在于海马体有许多皮质醇受体。海马体负责创建情景记忆,特别是关于空间关系和个人自传体验的记忆。也许由于分离痛苦的时期,皮质醇分泌升高使幼年动物对熟悉和令人安慰的老地方(如家)以及重聚时对父母的爱产生更强的记忆。少量分离焦虑可能有助于巩固社会记忆和社会纽带。另一个相关假说是,皮质醇促进认知,有助于找到家和社会重聚的益处。
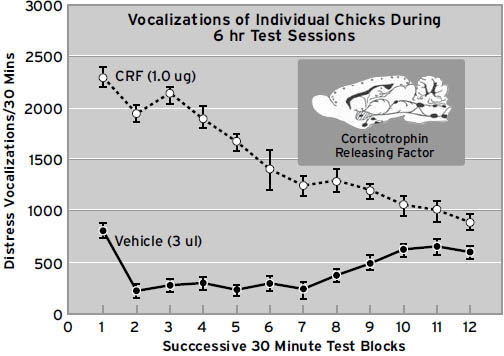
图9.3. 促肾上腺皮质激素释放因子(CRF)是垂体肾上腺压力反应的主要大脑激活剂,但它也有广泛的皮层下回路,如插图所示。当少量CRF被注射到3周龄幼鸡的脑脊液中时,它在与鸡群分离6小时后不再表现出许多分离痛苦发声(鸣叫)。(未发表数据,Panksepp, 1984。)
当压力系统运作良好时,皮质醇反馈到PVN中的大量受体,这种反馈作用使PVN停止向垂体前叶泵出CRF,从而停止产生ACTH。没有ACTH促进皮质醇的释放,整个压力反应就会减弱。然而,如果这种自我调节反馈效应受损,皮质醇的产生会持续不减,并最终对身体和大脑产生有害影响,造成慢性压力感(有时是躁狂兴奋)。在易感个体中,最终会导致抑郁甚至海马损伤;这些负面影响有时可以通过抗抑郁药逆转。
在极端情况下,长期高水平的皮质醇释放到循环系统中会导致海马体过度紧张,甚至造成永久性损伤。过量的皮质醇最终可能伤害甚至杀死海马体中的神经元,导致记忆丧失。由于伦理方面的考虑,缺乏可以通过这种方式研究的人类详细神经科学证据,但那些经历过战争创伤或其他暴行(如大屠杀幸存者)的人通过结构性脑成像常常表现出一定程度的海马萎缩,尽管实际损伤的证据很难证明。然而,对照研究表明,在没有养育关系的环境中长大的猴子可能会经历长期高水平的皮质醇,这确实会伤害海马体(Nelson & Bloom, 1997)。事实上,研究发现,受虐儿童以及长期遭受性虐待的成年人或经历过过度战时压力的士兵,往往比未受压力的个体拥有更小的海马区域(Conrad, 2008; Irle et al., 2009)。由于海马体对于创建许多类型的记忆至关重要,包括我们所有的情节性(自传体)记忆,这些研究表明,早年的长期社会剥夺可能会损害心智的发育成熟。通过这种方式,我们可以想象早期生活压力以及后期生活压力如何可能损害由海马体介导的认知功能。
垂体肾上腺CRF-ACTH-皮质醇系统的失衡也会随之导致统称为生物胺的化学物质耗竭,特别是去甲肾上腺素(norepinephrine, NE)、血清素(serotonin)(即5-HT或5-羟色胺)和多巴胺(dopamine, DA)。最初,CRF的释放会强烈激活这些神经系统。但当CRF释放持续时,这些系统的突触化学资源可能会耗竭。这导致大脑生化系统的许多继发性变化,包括神经生长因子水平降低和大脑炎症过程增加(Cirulli et al., 2009; Harro & Oreland, 2001; Miller et al., 2009)。当大脑的生物胺耗竭时,人类和动物也容易抑郁,这可能紧随持续过量的CRF释放之后出现。动物和人类的抑郁症状可以通过在体内建立这种生理变化模式来实验性诱导。事实上,长期施用CRF以及生物胺的耗竭会强力促进动物的抑郁反应,而施用CRF受体阻滞剂可以抵消人类的抑郁(Holsboer & Ising, 2008)。
我们现在将提供上述观点与理解和治疗抑郁症关系的总体概述。我们这样做基本上不引用文献,因为压力相关的抑郁诱发以及传统的抑郁症药物治疗已经在已引用的现有文献以及各种精神病学教科书中得到了非常广泛的涵盖(例如,Panksepp, 2004)。这个概述只是为讨论悲伤系统(GRIEF system)在抑郁症中的作用以及一些潜在的新型干预措施提供桥梁,例如尝试直接补充积极情感化学物质,同时减少消极情感化学物质的影响。
我们还不确切知道汇聚的压力相关脑变化最终如何导致表征临床抑郁症的持续心理变化。但我们确实知道,抵消生物胺低可用性的药物(即血清素、去甲肾上腺素和多巴胺再摄取抑制剂)往往具有抗抑郁作用。目前使用的大多数抗抑郁药物通过阻断生物胺的再摄取机制来促进其可用性。因此,抗抑郁药使生物胺能够在突触处停留更长时间,突触是神经元之间的间隙,这些化学信使在此处被释放和接收。最广泛使用的抗抑郁药是选择性血清素再摄取抑制剂(selective serotonin reuptake inhibitors, SSRIs),如百忧解(Prozac)。顾名思义,SSRIs具有选择性作用,仅使血清素在突触处更可用。然而,这些药物长期使用也会下调血清素受体,这可能启动大脑调整,当停止慢性用药时,最终可能产生不良后果。
其他类型的再摄取抑制剂影响所有这三种关键胺类的可用性,至少在短期内最有效,而那些作为单一胺类促进剂的药物效果较差。这些药物需要数周时间才能显示疗效的事实表明,治疗益处是继发于长期脑变化的,例如神经生长因子活性的逐渐增强,这应该有助于修复像海马体这样受压力损伤的脑区。然而,应该注意的是,短期使用SSRIs可以显著缓解动物的分离痛苦(separation distress),这导致使用氟西汀(fluoxetine)(商品名百忧解)等药物治疗宠物的分离痛苦问题。
其他抗抑郁药物,如单胺氧化酶(MAO)抑制剂,通过抑制降解这些神经递质的酶的作用,增加生物胺的可用性。单胺氧化酶抑制剂和其他更早的、SSRI之前的抗抑郁药物,由于与新药相比副作用更多,在精神病学中已不如过去那样广泛使用。然而,无论其具体机制如何,旧的和新的抗抑郁药物的效果都是使生物胺在突触中保持更长时间的可用性,从而为接收这些化学信使的神经元提供更多的刺激。同样,由这些药物引起的大脑调整所产生的持续长期后果仍有待充分研究。
关于这类药物如何产生抗抑郁效果,需要进一步阐述。如前所述,生物胺(如去甲肾上腺素、血清素和多巴胺等神经化学物质)通常充当神经递质,或称为导致神经元放电或抑制放电的化学物质。神经递质释放到突触后不久,会通过两种方式之一从突触中清除。首先,酶可以降解神经递质,将分子分解为非活性成分。其次,神经递质可以被释放它的神经元重新摄取(“再摄取”(reuptake)),从而使分子脱离作用。当生物胺充当神经递质时,可以通过这两种机制中的任何一种来促进它们在突触处的可用性——即通过抑制其化学失活或通过抑制其返回到起源神经元。然而,如前所述,这些药物的强效抗抑郁作用可能需要数周才能发生,这种延迟表明大脑中正在激活许多”下游”变化。这些更远端的变化可能最有益,包括许多修复过程,如减少脑组织炎症,以及新神经元的增殖和海马体中神经元的增长,海马体是正常和健康大脑功能所必需的区域。其他变化,如受体敏感性降低,从长远来看可能不那么理想。
虽然我们对大脑功能层面的抑郁症病因没有确定的答案,但有许多神经化学候选物。不幸的是,这些潜在的致病因素通常在没有任何支持性情感数据的情况下被提出。目前受关注的化学物质之一是被称为脑源性神经营养因子(BDNF)的大脑”肥料”(brain-derived neurotrophic factor),但还有许多其他生长因子候选物。这些都是”转录因子”(transcription factors),这意味着它们在基因调控的关键点发挥影响,在那里许多下游遗传途径被开启。一个显著的发现是,许多这些生长因子也会被具有抗抑郁效果的健康活动所激活,如运动。即使是幼年动物的身体玩耍也能促进大脑中BDNF的可用性(Gordon等人,2003年),并能增加其他生长因子如胰岛素样生长因子的水平(Burgdorf等人,2010年)。我们将在下一章强调玩耍的这些和其他功能。然而,很明显,感知到的社会支持是个体能够从重度抑郁疾病中恢复的最佳指标之一(Leskelä等人,2006年)。
目前的抗抑郁药物确实有助于逆转压力造成的一些破坏。然而,我们知道,当悲伤(GRIEF)被唤起时——这是压力诱导抑郁症的关键因素——会出现安慰性化学物质如内源性阿片类物质的耗竭(Kennedy等人,2006年;Zubieta等人,2003年)。我们还从大量研究中了解到,内源性阿片类物质和阿片制剂可以产生快速的抗抑郁样效果。这些事实表明,可能还有另一种对抗抑郁症的方法,即通过增强大脑阿片系统的活性。如前所述,在现代抗抑郁药物出现之前的日子里,增强阿片系统激活通常是通过施用阿片类药物来实现的。然而,由于其臭名昭著的成瘾性,阿片类药物的施用被放弃,转而采用那些相对较弱的现代抗抑郁药物,这些药物旨在干预生物胺的活性不足(尽管如前所述,这并不意味着抑郁症主要是由于这些递质的缺乏)。
当今的神经科学家应该重新考虑这样一种可能性:抑郁症在很大程度上是由于大脑中快乐化学物质的缺乏,特别是那些支持社会纽带安全感的物质。例如,如果强烈而持续的悲伤期通过大脑μ-阿片活性的急剧下降而在一定程度上促进抑郁症,我们终于有办法解决这种失衡。现代神经科学已经发现了能够促进μ-阿片活性而不会有严重成瘾危险的药物。一个例子是混合型阿片受体激动剂/拮抗剂丁丙诺啡(buprenorphine)(现在被广泛用于治疗麻醉品成瘾)。这种药物只在极低剂量时促进阿片活性,而在高剂量时则阻断受体活性。在低剂量下,丁丙诺啡被发现对多次其他药物治疗失败的人来说是一种强效抗抑郁药(Bodkin et al., 1995)。混合激动剂/拮抗剂作用意味着人们不会对这种药物产生强烈成瘾,因此它可以安全地用作戒断阿片类药物成瘾者的药物。该药物提供足够的阿片活性,从而提供心理缓解,以防止阿片类药物戒断的痛苦戒断效应。因此,丁丙诺啡或许也可以更广泛地用作一种有效的快速起效的抗抑郁药,特别是在那些从传统疗法中得不到缓解的个体中。然而,如果没有适当的双盲研究和随后的政府监管批准,它在标准精神病学实践中的广泛使用是不太可能的。显然,在这种方法获得普遍医学认可之前,需要进行一些良好对照的研究。
在这种情况下,值得注意的是,安慰剂效应在抗抑郁试验中非常常见,通常效果如此强烈,以至于其益处与广泛使用的SSRIs(选择性血清素再摄取抑制剂)所获得的益处一样强。这部分反映了日常轻度抑郁反应通常是自限性疾病的事实。此外,从目前的社会大脑分析来看,安慰剂的抗抑郁益处是可以预期的,因为安慰剂部分通过内源性阿片释放起作用(总结见Panksepp, 2006c, 2011a)。积极的社会互动释放大脑阿片,提供积极的社会感受,而安慰剂效应可能部分反映了这样一种认知:心理健康专业人员和其他重要他人关心自己的抑郁情绪。这种被关心的感知可能增加大脑阿片的释放,使抑郁的人感觉更好。换句话说,抑郁症中的安慰剂效应至少部分反映了社会支持激活大脑阿片系统的能力。当然,这也是人际关系的情感质量在心理治疗干预结果中如此重要的原因之一。
然而,μ-阿片耗竭并不是神经科学家正在探索的唯一与抑郁相关的阿片失衡。从强烈悲伤到绝望和抑郁的转变也伴随着SEEKING(寻求)系统唤醒的减弱。由强啡肽(dynorphin)增强效应引起的κ-阿片活性升高,已被确定为SEEKING系统参与的化学变化基础。如上所述,强啡肽与κ-阿片受体结合。目前有相当多的兴奋点在于,发现医学上安全的拮抗剂能够阻断κ-阿片受体,将抑制强啡肽的作用,并被证明作为抗抑郁药非常有用(Knoll & Carlezon, 2010)。事实上,这也是丁丙诺啡的药理特性之一,它在相同剂量下可以阻断引起烦躁的κ-受体,同时通过μ-受体促进积极的快感。
SEEKING唤醒减弱的作用可能突出了抑郁症一个鲜为人知的”适应性”效应。进化精神病学家已经考虑这样一种可能性:即使严重抑郁症可能对心理经济产生各种有害影响,但当社会支持丧失时,某种程度的抑郁可能促进生存(这一想法最初由约翰·鲍尔比提出,并由Watt & Panksepp, 2009在神经科学上发展)。例如,在一段强烈的分离痛苦和发声抗议期之后,表明最初的恐慌反应,这有助于父母找到失散的后代,退行到行为抑制的绝望或沮丧阶段以保存身体资源可能是适应性的。这种抑郁状态可能有助于阻止无助的生物体进一步偏离安全区域。沉默也能最大限度地减少被捕食者发现的可能性。同样,另一个在减少痛苦发声方面非常有效的消极情绪过程是FEAR(恐惧)。总之,如果最初的抗议没有实现团聚,沉默的绝望反应可能是一种有用的次要策略,以优化父母最终找到失散后代活着的可能性。
现代神经科学现在允许我们区分由GRIEF(悲伤)和FEAR(恐惧)引起的焦虑——区分隔离恐慌和人们在预期受伤、死亡或其他即将发生的厌恶事件时感受到的恐惧。当然,这两个系统是相互作用的。例如,经常被单独留下的儿童会经历分离焦虑,但他们也会害怕被单独留下的前景,并再次感到痛苦。换句话说,他们可以在更高的认知层面上害怕GRIEF。
这两个系统在神经解剖学和化学方面也存在一些重叠。事实上,许多情感系统中都存在解剖学和化学上的重叠(例如,所有情感系统都涉及进化上古老的中脑导水管周围灰质PAG,这是初级过程情感生活的核心,而神经递质GABA、去甲肾上腺素、血清素,可能还有多巴胺都调节着所有情感系统)。尽管如此,FEAR和GRIEF系统在解剖学和化学基础上仍可以区分。CRF可以激活这两个系统,但这两个系统在许多其他方面的化学性质也不同。例如,在极低剂量下,阿片类药物能强烈减少分离哭泣。但需要更高的剂量才能适度减少焦虑或预期恐惧的行为指标。另一方面,经典的苯二氮卓类抗焦虑药物,如氯氮卓(利眠宁Librium)和地西泮(安定Valium),在低剂量下能有效减少FEAR反应,但在减少表明GRIEF的分离叫声方面效果不佳。在行为方面,我们刚才提到分离痛苦可能导致习得性焦虑:人们很容易对感到恐慌产生恐惧。从恐惧到绝望的类似因果路径并不明显,因为当动物极度恐惧时,痛苦叫声通常会减少。当然这符合良好的进化意义,因为动物在危险时会感到恐惧,如果它们在这些危险时刻发出叫声,就更可能吸引捕食者的注意。不过,在更高的认知(三级过程)层面上,如果慢性焦虑也使人类和动物更容易抑郁,这并不会让任何人感到惊讶。
无论如何,有很多理由相信,精神病学家称为”惊恐发作(panic attacks)“的神秘不安感,可能主要来自社交分离痛苦GRIEF网络的突然唤醒,而不是像许多理论家目前认为的那样来自FEAR网络。控制惊恐发作的神经化学与控制一般焦虑的神经化学之间的药理学区别,最早是由精神病学家唐纳德·克莱因在1960年代初期的细致工作揭示的(详见Panksepp, 1998a)。克莱因发现,新发现的苯二氮卓类抗焦虑(抗FEAR)药物,如利眠宁和安定,对惊恐发作的发生率几乎没有益处。然而,三环抗抑郁药丙咪嗪在平息此类发作方面非常有效。尽管焦虑患者最初声称三环药物对他们没有益处,但护士报告说这些患者抱怨惊恐发作的频率减少了。实际上,统计结果显示,患者在服用丙咪嗪后确实惊恐发作次数减少了很多。显然,患者没有关注到这些改善,因为这种药物没有减少与该疾病相关的预期焦虑——即对发作本身的恐惧。这些患者仍然害怕惊恐发作;也许他们也需要针对FEAR的药物治疗。
在这种背景下,重要的是要注意丙咪嗪在减少包括狗和灵长类在内的许多物种的分离痛苦方面非常有效(见Panksepp, 1998a)。这表明惊恐发作,就像分离叫声一样,是唤醒的GRIEF而不是FEAR的功能。支持这一假设的证据表明,惊恐发作的生理方面可以通过阿片类活性降低来促进(Preter & Klein, 2008)。还有进一步的临床证据表明惊恐发作与社交丧失有关:那些遭受惊恐发作的人通常有童年分离焦虑的历史。此外,惊恐发作和分离痛苦都让人感觉好像自己的舒适或稳定中心被突然移除了。两者都伴随着虚弱感和呼吸急促,通常伴有哽咽感,好像喉咙里有肿块。
GRIEF系统的失衡在广泛的情绪障碍中起着关键作用,因为许多精神疾病根源于无法享受温暖人际依恋的安全感。我们已经提到了惊恐发作、抑郁症和自闭症等不同的疾病。也许一系列社交恐惧症和人格障碍也将被纳入GRIEF病理学的范畴。我们无法开始审视所有这些情况。相反,我们将关注第一个也是最关键的社会纽带——婴儿/母亲关系。儿童情感忽视,甚至更严重的虐待,所造成的精神后果是巨大的(Heim, et al., 2010)。
在过去几十年中,发展心理学家构建了关于儿童与母亲或主要照料者之间社会依恋本质的连贯理论观点。他们观察到儿童表现出各种依恋”风格”或气质,这些具有强烈的环境前因。安全型依恋儿童对从父母或照料者那里获得社会支持充满信心。他们通常外向,倾向于以乐观和热情面对生活。这些儿童成长为适应良好的成年人,因为他们从”安全基地”开始。一般来说,他们拥有良好的关系,在生活追求中取得成功。
然而,当母亲未能养育好孩子时,孩子长大后会形成不良的依恋关系。不安全依恋的儿童表现出两种主要的情绪和行为模式。一些儿童过度黏人,似乎需要比通常更多的照顾者关注。另一些儿童选择保持距离,回避社交场合,大概是因为他们不确定能否获得自己渴望的积极支持和反馈(Ainsworth, 1982)。在过去30到40年里,大量关于人类婴儿的临床研究强调了母亲需要对孩子的情绪给予共情关注(Beebe & Lachmann, 1988)。当母亲在情感上与婴儿保持同频时,就会促进情绪健康。
当然,这些理论依赖于行为观察和关于儿童心理活动的描述性心理推断。直到近年来,神经科学家才能够将依恋理论转化为大脑中发生的具体变化。神经科学家用表观遗传(epigenesis)来理解这些大脑变化——由经验导致的基因表达。我们之前提到,基因表达是一个休眠基因变得活跃的过程。表观遗传涉及经验依赖的基因表达;它是出生后因孩子在世界中的经历而发生的基因表达。
表观遗传可能看起来是个奇怪的概念,因为我们习惯于认为我们与生俱来的遗传禀赋会精确且不可磨灭地决定我们一生中将展现的特质。有些基因确实如此。例如,表现出分离痛苦(DVs)的能力肯定由出生后不久就变得活跃的大脑网络决定,这就是为什么几乎所有幼年哺乳动物和鸟类在被单独留下时都会哭泣。因此,构建这类大脑系统的信息肯定是由基因决定的。然而,人类或动物一生中的某些经历可以通过表观遗传机制使基因表达得更强或更弱,这彻底改变了我们对先天与后天的理解(Szyf et al., 2008)。因此,当一个先前休眠的基因在特定脑回路中表达时,它可以产生脑细胞以前从未产生过的蛋白质和神经肽。许多这些被激活的神经化学通路肯定会改变情感脑心(BrainMind)功能。因此,我们看到表观遗传过程如何影响情绪行为和感受。
当幼年动物受到母亲照料时,这些经历会导致影响大脑功能的基因活性发生表观遗传变化。这种可变的大脑功能可以产生个体特有的特征和行为——比如上面提到的依恋风格。表观遗传可以在受到良好母育的婴儿中创造健康的大脑,但它也可能引发各种类型的心理问题。研究表明,如果母亲情绪低落因而对婴儿缺乏回应,人们会在孩子的行为以及大脑组织中看到异常(Meaney, 2001; Tronick, 2007)。也许这些孩子会发展出不敏感的大脑催产素或阿片系统。如前所述,催产素无法安抚痛苦的母亲,从而无法为她的孩子提供安慰,这可能是因为她的催产素系统不再能够补充和增强内源性阿片类物质的作用。有许多可能性需要考虑。
一个众所周知的社会问题是,年幼时受到不良照料和形成不安全依恋的成年女性母性冲动发展不良。在某种程度上,被母亲不良养育的经历可能会在幼年女性的大脑中造成表观遗传变化,使她们在有了自己的孩子时不太可能成为最好的母亲。早期母育不良的一个结果是大量行为变化,这些变化部分源于大脑对压力反应方式的表观遗传变化(Meaney, 2001; Szyf et al., 2008)。关于这个主题的大部分研究都是在大鼠身上进行的,但最近有报道称,受虐儿童成年后最终自杀的大脑表现出的表观遗传变化与受到较少母性照料的大鼠观察到的变化非常相似(McGowan et al., 2009)。我们之前关于皮质醇对海马体有害影响的讨论可能反映了表观遗传的另一个例子。这项研究帮助我们更好地理解为什么受虐的年轻人往往成为表现不佳的父母,延续着儿童忽视甚至虐待的跨代循环。不安全依恋,也许是由于低阿片反应性,可能会代代相传。作为一个群体,自杀者的大脑显示出低阿片活性的指标(Gross-Isseroff et al., 1998),药物增强大脑阿片满足感,比如使用低剂量的”安全”阿片类药物如丁丙诺啡(buprenorphine),可能可以减少自杀想法。当然,这也可以通过关爱他人更持续的积极情感关注来实现。
大多数关于依恋的生物学研究强调最近进化的高级脑区的变化:新皮层,特别是两个大脑半球的功能。这些研究由临床医生艾伦·肖尔(Alan Schore)进行了广泛总结(另见McGilchrist, 2009),指出大脑的许多部分在出生时并未完全形成,这些区域的发育由经验依赖的表观遗传变化介导。与越来越多的神经科学家一起,肖尔研究了母子互动的质量如何控制大脑的发育,无论是好是坏(Schore, 2001)。
对大脑高级区域的研究表明,出生时只有初级体感皮层(primary somatosensory cortex)在代谢上高度活跃(Chugani, 1996)。新皮层(neocortex)的其余部分仍在发育过程中。Schore将注意力集中在右侧大脑半球,该半球在生命的前18个月比左半球表现出更强劲的生长高峰。右半球在生命的前3年保持主导地位(Chiron et al., 1997)。显然,这是婴幼儿开始与父母建立关系的时期。右半球对生活保持着更情绪化、更整体性的态度,而较晚成熟的左半球最终提供更多分析性认知技能,远离社会敏感性。Schore关注大脑右侧,因为研究表明,这一侧对外部刺激特别具有情绪反应性,比如童年早期的养育性触觉体验(Kalogeras et al., 1996)。这些体验深受母亲养育质量的影响,至少在大鼠中具有永久性的大脑效应(Meaney, 2001)。儿童大脑的发育依赖于经验,并直接受到母子关系输入的影响。
Schore进一步认为,眶额皮层(orbitofrontal cortex, OFC)作为一个古老的皮层结构,在第一年的最后一个季度到第二年中期经历一个关键的成熟期。同样,这是母子关系蓬勃发展的时期。在此时间框架内与母亲的经历产生表观遗传变化,这些变化有助于眶额皮层的发育或发育不全,该皮层在处理人际信号及其情绪意义方面发挥关键作用。当大脑的这个区域受损时,人们可能表现出较差的社会调节能力,转向更情绪化冲动,甚至反社会的气质极端(Adolphs et al., 2003)。发育良好的眶额皮层还调节自主神经系统的许多方面,自主神经系统产生情绪体验的生理成分(Porges, 2009b)。因此,眶额皮层在情绪调节中发挥关键作用(Schore, 1994)。如果不良的母子互动干扰了这个大脑区域的成熟,孩子可能在一生中都会遇到调节情绪的困难。由于情绪调节是心理健康的核心特征,眶额皮层的良好功能至关重要。尽管我们目前还不了解大脑成熟这些关键方面所涉及的表观遗传变化的细节,但我们可以假设这些变化提供了导致大脑健康或不健康发育的关键机制。
尽管新皮层、眶额皮层和海马体功能受损的幽灵令人不安,但动物研究表明了一个更可怕的场景。实验证据表明,经历极端体验的人类或动物可能会发展出原始过程边缘情绪网络的长期敏感性增加或降低。这些对皮层下情绪系统的改变也是由表观遗传介导的。例如,如果一只动物经历了许多可怕的经历,其恐惧系统可能会被永久性敏化;这样的动物很容易受到惊吓(LeDoux, 2002)。这种表观遗传变化可能导致各种其他情绪系统的病理性过度敏感和过度反应,特别是那些调节分离痛苦/悲伤反应的系统。
如果我们考虑皮层下情绪系统对成熟中的新皮层施加决定性影响的可能性,那么大脑深层情绪区域的持久发育变化就显得更加重要。有人提出,早期依恋困难可能导致边缘情绪区域与新皮层之间的神经连接减弱(Schore, 1994)。这可能意味着大脑的皮层下区域对皮层发育的贡献较少。相反,由于新皮层通常抑制边缘表达,这也可能有助于解释为什么经历过依恋困难的人通常在情绪上缺乏抑制。
悲伤可能是人脑最强大的情感网络,我们永远无法完全被现代文化制度的许多安全网所屏蔽。事实上,蓝调音乐等文化形式从业者的许多艺术丰富和富有成效的职业生涯证明了这一基本人类体验特征的力量。在最好的脑成像工作中,悲伤比任何其他情绪更显著地”点亮”我们的大脑(Damasio et al., 2000)。我们在这些图像中看到的模式与我们通过研究同类动物所了解的分离痛苦/悲伤系统的神经解剖学很好地匹配(图9.1)。当我们与所爱的人建立安全依恋时,我们获得了一生的礼物。当依恋过程受损时,高级心理装置内精神痛苦的多样表现可能导致终生的慢性痛苦感。这种痛苦常常妨碍我们与他人建立关系的方式。尽管如此,尽管存在这些变化,人类是一个非常有韧性的物种,因此上述所有对情绪健康的影响在个体之间差异很大。
心理治疗师认为移情对他们的工作至关重要。移情是指患者倾向于在治疗环境中重现他们与重要他人建立的相处方式。患者对治疗师的看法和感受往往反映了患者对家庭成员,特别是对父母的感受和看法(Pulver, 1995)。换句话说,所有患者,实际上所有人,都会发展出习惯性的情绪反应方式来回应他人,而这些情绪习惯在很大程度上是由早期关系塑造的。如果你在健康的环境中长大,你会以开放和接纳的方式对待整个世界。这是一种移情反应,因为你甚至可能对不择手段的人保持开放。事实上,当你第一次遇到善于操纵的人时,你可能很难理解他们。相反,在险恶环境中长大的孩子往往会以怀疑和敌意看待每个人,他们可能无法理解或相信任何人有善意的动机。移情是一种普遍现象;我们经常积极地认同我们钦佩的其他人,它在心理治疗中是有用的,可能也是必不可少的,因为它给患者和治疗师提供了机会来处理麻烦的过去经历留下的情感和行为痕迹。
在精神分析的早期,治疗师被敦促隐藏他们个性的某些方面。这个想法是,治疗师的个性应该是一块”白板”(tabula rasa),患者可以在上面投射他或她的移情反应,而不必担心被治疗师的性格所污染。这个模型仍然有一定的价值,因为治疗师不应该用自己的问题给患者增加负担。毕竟,必须记住,两人中有一个是来访者,通常是付费来访者。
然而,治疗师应该保持情感中立的观念不仅是不可能的,而且对治疗工作有害。保持中立是不可能的,因为所有人(所有哺乳动物)都有对积极关系的内在需求。因此,不微笑和/或没有情感的治疗师会被负面看待。这种治疗立场会排斥大多数患者,只会吸引那些发展出受虐倾向的人,或者那些渴望在不制造麻烦的情况下从心理治疗培训中”毕业”的人。治疗师对患者持有积极态度并据此行事是正常的,也是必要的。当然,积极的倾向总是有可能使治疗师对患者性格中的负面特征视而不见。然而,当治疗成功时,应该在不破坏整体积极治疗关系的情况下处理这些负面特征。这种积极的联盟可以为患者提供一个安全港湾,让他们认识到自己转移到他人身上的包袱实际上是一系列习惯、信念和感受,它们属于完全不同的关系,甚至可能是不同的时代。
在整本书中,我们一直强调希望未来心理治疗和精神药理学能够携手合作。事实上,一些新兴药物已经可以促进标准心理治疗方法——例如,D-环丝氨酸(D-cycloserine)在暴露疗法期间给药时可以在神经层面促进治疗性改变(Norberg et al., 2008)。可以想象,通过适量剂量的安全阿片类药物实现短期药理学诱导的情感稳态,促进积极情感和信心,可能会促进治疗过程,特别是如果用于以更积极的情感背景重新巩固麻烦记忆的情况下(见[第6章])。同样,需要更多研究来了解快速起效的情感药物如丁丙诺啡(buprenorphine)是否可以用于促进悲伤障碍的长期稳态。当然,这些心理治疗的药理学辅助手段只有在治疗本身是健全的情况下才能产生有益的结果。然而,这些药物可能在短期内提供动力,使一些患者能够克服仅靠心理治疗难以跨越的情感”障碍”。
特别紧迫的是进行适当设计的研究,尝试在来访者开始洞察他或她的问题时促进获益的巩固。可能适量剂量的悲伤抑制剂,如鼻内催产素(oxytocin)(目前严格的实验性药物)或极低剂量的丙咪嗪(imipramine),甚至可能是许多其他常见抗抑郁药或抗焦虑药的温和但快速的抗压力精神效果,在时间上以有利的方式明智地结合使用,可能有助于促进持久的治疗性改变。积极自我形象的记忆重新巩固也可能通过敏感地结合使用精神药理学和心理治疗手段来促进。这也可能使我们远离目前”向来访者长期投药”的标准做法,而不处理他们的心理生活。这些是未来研究的关键重要方向,并有一些有希望的先例:例如,正如反复显示的那样,抗抑郁药和心理治疗的联合使用比单独任何一种干预更有效(例如,Holtzheimer & Nemeroff, 2006)。
GRIEF系统对心理健康至关重要,它可能是导致抑郁和某些类型慢性焦虑的最重要系统之一。另一个则是SEEKING系统(Panksepp & Watt, 2011)。所有哺乳动物都需要与他人建立联结,而爱的典型哺乳动物纽带是母亲与婴儿之间的关系。在这种关系中,母亲的CARE系统培育并敏感地回应婴儿的GRIEF系统——回应婴儿感受亲密和依恋的需求。这不仅仅是被喂养和庇护的稳态需求,而是与母亲或照顾者建立安全温暖依恋的情感需求。
悲伤教会我们很多关于爱的东西。当我们仅仅想象与最亲近的人分离时,可能会感到悲伤或警觉。这种感觉告诉我们,我们与所爱之人的情感纽带很可能经受住失望、失落和生活中许多变化的考验。一旦我们意识到最好的抗抑郁化学物质将是那些能调动积极社会情感系统力量的药物,比如内源性神经化学物质如催产素,它可以缓解分离痛苦的心理痛楚,我们就会在开发新的抗抑郁药物方面取得更多进展,例如谷氨酸受体拮抗剂(Machado-Vieira et al., 2009; Skolnick, 2009)和更间接的调节剂(Burgdorf et al., 2011),在这个过程中我们可能会学会使用高效药物,如”安全”阿片类药物(Bodkin et al., 1995)。当然,最好的良药是我们从爱的关系中获得的温暖和慰藉。它们滋养人类的灵魂。
让我们重申,虽然GRIEF系统可以产生一种”焦虑”形式,但这种焦虑与来自FEAR系统的恐惧预期并不相同。FEAR和GRIEF由不同的大脑结构以及在大脑不同部位运作的重叠但也不同的脑化学物质集合所支持。我们尚未完全理解GRIEF系统在临床抑郁和慢性焦虑中所起的作用。我们在利用现有药物和设计治疗这些病症的新有效药物方面还有很长的路要走。事实上,我们并不真正了解当前抗抑郁药物是如何起作用的;显然不是由于它们的短期药理作用。有些人认为这些药物启动了有助于修复受损大脑功能的生长因子,这反映在许多抗抑郁药促进海马神经增殖的能力上(Boldrini et al., 2009)。
另一方面,我们很清楚适量的阿片类药物是有效的抗抑郁药,能迅速见效,而目前的药物通常需要数周才能完全发挥作用。这种快速起效的抗抑郁药是迫切需要的。不幸的是,阿片类药物在大量服用时可能成为滥用药物,这一事实促使研究人员忽视了它们在精神医学用途方面的巨大潜力。除了丁丙诺啡等安全阿片类药物满足这一需求的潜力外,谷氨酸阻滞剂作为快速抗抑郁药的出现(可能通过阻断GRIEF的心理痛苦)正在引领安全、非致幻治疗策略的发现,这些策略也认真对待动物大脑中的积极情感过程(Burgdorf et al., 2011)。希望对GRIEF系统的更好理解也能鼓励精神病学研究人员将注意力转向这些药物可能带来的有益效果。
同样,痛苦记忆在积极情感态度的背景下被重新语境化得越多,客户从长远来看可能会越好(见第12章)。正如我们将在下一章看到的,游戏性是我们大脑能产生的最积极的社会情感感受之一,但尚未在心理治疗背景中得到系统或良好的应用。肯定有办法使这种强大的积极情感成为治疗互动中更常见的方面。我们或许应该记住诺曼·卡曾斯(Norman Cousins,1983)的著名观点:笑声可能是我们最好的良药之一。
多么大胆啊
为永恒而游戏,
像峡谷倾泻而下那样游戏,
像河流奔涌那样游戏。
——鲍里斯·帕斯捷尔纳克(Boris Pasternak),《酒神节》,Anesa Miller为米哈伊尔·爱泼斯坦(Mikhail Epstein, 1993)翻译
在追求自由的过程中,俄罗斯伟大的小说家和诗人鲍里斯·帕斯捷尔纳克(1890-1960;1958年诺贝尔文学奖获得者)在《酒神节》中写下了上述关于深度游戏的诗句。哲学家米哈伊尔·爱泼斯坦(1993)继而反思道:“帕斯捷尔纳克的诗句……传达了自然的游戏,这是文化可能做到的理想——游戏,不是像下棋者那样游戏,而是像河流那样游戏。”所有哺乳动物幼崽在它们的心智中发现,用爱泼斯坦的话说,“一个野性的、淘气的、喧闹的生灵”,与”整个自然——如同一个调皮的孩子”的自发性产生共鸣。1
问孩子们他们最喜欢什么,他们几乎无一例外的回答是”玩!“各种形式的玩耍活动给所有年幼的哺乳动物带来巨大的快乐。在最早的年龄段,身体上的打闹嬉戏——也被称为粗暴游戏(rough-and-tumble play)——是最有趣的活动,这一点从伴随这些活动的大量笑声就能看出,男孩和女孩几乎一样多([图10.1])。心理学家对游戏写了很多,但他们不知道存在多少个初级过程的游戏系统。可能只有一个——就是本章描述的身体游戏系统。然而,成熟的人类心智中的玩耍性延伸到了我们高级心理装置的最远处,到达了我们想象力的平流层,以至于我们可以用最聪明和最离谱的笑话来逗笑彼此。我们不会关注人类心理装置的那些更高级的积极心理问题,这些问题已经在各种积极心理学著作中得到了探讨(例如,Sheldon等人,2011)。
身体玩耍性是每个年幼哺乳动物的与生俱来的权利,也许还有许多其他动物。有两本近期出版的书和两本较早的书对跨物种的玩耍性提供了出色的总结,远超过本章所能总结的内容(Aldis,1975;Burghardt,2005;Fagen,1981;Pellis & Pellis,2009)。还有一本关于老鼠游戏行为的经典专著,仍然很值得一读(Groos,1898),以及一本关于人类游戏的后续研究(Groos,1901)。现在可以确定的是,在哺乳动物大脑中存在一个由基因决定的、介导积极情感的游戏网络(Burgdorf等人,2007;Panksepp,1998a),尽管许多细节仍有待科学分析。到目前为止,大部分神经科学研究都是用实验室老鼠完成的,所以我们无法确定这些经验在多大程度上适用于人类。事实上,我们对人类的初级过程粗暴游戏(以下称为游戏系统)几乎一无所知,尽管有一些关于笑声的相关研究。但由于游戏系统集中在皮层下脑区,就像所有其他基本情绪系统一样,我们可以预期许多一般原则,特别是关于神经解剖学、神经化学和原始情感(社交快乐)的原则,将适用于所有哺乳动物物种。至于我们如何在心智的三级过程网络中将玩耍性转化为幽默,则不会。
很难定义游戏,但当你看到它时就会认出来。也许最好的一般定义是Gordon Burghardt(2005)最近提出的,包含五个标准:(1)游戏的适应性功能在游戏发生时并不完全明显;(2)游戏是一种自发的活动,为了它本身而进行,因为它很有趣(令人愉快);(3)游戏是成年活动的夸张和不完整形式;(4)游戏展示了许多重复性活动,有大量变化,不像严肃的行为那样缺乏灵活性;(5)动物必须吃饱、舒适和健康才能进行游戏,所有压力源都会减少游戏。Burghardt(2005,第82页)试图将所有这些特质归纳为一句话:“游戏是重复的、功能不完全的行为,在结构、情境或个体发生上与更严肃的版本不同,并且是在动物处于放松或低压力环境时自愿发起的”[原文强调]。
图10.1. 年幼(4至7岁)女孩和男孩在半小时无玩具的自由身体游戏中的游戏时间进程;然而,每5分钟有音乐(爱尔兰吉格舞曲),在间隔的5分钟观察时段没有音乐,并对行为进行视频编码。在这里我们看到,笑声的数量在游戏过程中系统性地减少,男孩和女孩之间没有重大差异。此外,还记录了总共19种其他游戏姿态,几乎没有一种显示出性别差异,除了”从前面推”,女孩比男孩做得少。结论是,女孩和男孩在展示身体游戏的冲动上没有实质性差异,过去看到的性别差异可能是由于学习造成的(数据改编自Scott和Panksepp,2003)。
正如人们所看到的,动态的社交互动并未被列为标准,这使得Burghardt可以将探索性的乐趣纳入游戏概念,这可能主要是由于动物使用它们的寻求系统来获得个人乐趣。然而,对我们来说,社交形式的游戏,通常采取”打闹游戏”的形式,才是最引人注目和最快乐的游戏形式,有专门的大脑系统,也将探索性寻求的冲动纳入其领域。此外,对我们来说,初级过程的游戏必须部分由社交神经回路来定义,除了寻求系统之外,这些回路产生了年幼动物相互玩耍的动态特质。我们对社交玩耍性的最初描述之一是,当两只幼年大鼠”被放在一个非威胁性的环境中,它们很快就会开始展示激烈的打斗:动物们追逐并扑向彼此,有时是单方面的,有时是相互的,伴随着快速的角色转换。它们反复地戳和咬对方,通常是在颈背,但当一只动物被压制时也会咬腹侧表面”(Panksepp等人,1984,第466页)。这就是我们将在这里关注的那种游戏,因为它产生了最多的乐趣,正如老鼠在自发地沉溺于这种活动或被坚韧的人类以有趣的方式挠痒痒时发出的大量高频笑声类声音——50千赫兹的啁啾声——所突显的那样(Knutson,Burgdorf等人,1998;Panksepp & Burgdorf,2000)。
这些啁啾声与SEEKING系统的多巴胺奖励密切相关(Burgdorf等人,2007),有助于解释探索性游戏的乐趣,这种游戏以捕食练习的形式出现(例如小猫玩弄毛线球时)。我们现在知道,捕食行为,以及由此产生的幼年动物的追逐和扑跳形式,是SEEKING系统的发育产物(见第3章和第4章)。然而,正如我们稍后将讨论的,这种美妙的笑声般啁啾声现在可以用作直接测量大鼠积极情绪的指标(Panksepp & Burgdorf,2003),这甚至为理解成瘾药物的欣快愉悦打开了大门(Browning等人,2011;Burgdorf等人,2001;Panksepp, Knutson等人,2002)。
尽管探索性物品游戏可以带来极大乐趣,但没有什么能完全比得上全身心投入社交游戏时的那种彻底的欣快感,即使是未经训练的眼睛也能清楚地观察到这种状态。当你向院子里看去,看到松鼠在草地上跳跃和互相追逐时,几乎每天都能观察到这一点。
为什么会存在PLAY(游戏)冲动?它可能使幼崽能够学习非社交的身体技能,如狩猎、觅食等。它对于获得许多社交能力也肯定很重要,特别是初期的攻击性、求偶、性行为,以及在某些物种中的竞争技能,甚至可能是育儿技能。它可能是构建我们社交大脑许多高级功能的重要力量。游戏活动可能帮助幼年动物学会识别可以与之发展合作关系的个体,并知道应该避开谁。它们肯定通过游戏学习何时可以主导社交互动,何时应该优雅地退出、服从或接受失败。游戏也可能有其黑暗的一面。当动物游戏时,它们可能会学到可以欺负谁以及谁可以欺负它们。简而言之,大脑的PLAY网络可能有助于将个体融入分层的社会结构中,这将成为它们生活的舞台,这些网络还可能使它们做好准备应对生活中肯定会遇到的各种意外事件(Spinka等人,2001)。
PLAY冲动既强健又脆弱。它很脆弱,因为大量的环境操控都可以减少游戏——包括所有引发负面情绪状态的事件,如愤怒、恐惧、疼痛和分离痛苦;它对物种特有的恐惧刺激特别敏感,例如大鼠闻到捕食者的气味(见Panksepp,1998a,图1.1;Siviy等人,2006)。例如,如果实验室研究人员家里养了一只猫,而他上班前不小心更换衣服,他将很难研究大鼠的游戏,因为猫的气味本质上会吓到大鼠,而恐惧的大鼠根本不会游戏。同样,大鼠害怕光线充足的开放空间;它们在安全的洞穴中游戏,远离捕食者的注意。此外,饥饿是游戏的强大抑制剂(Siviy & Panksepp,1985),许多其他身体失衡也是如此,当然还包括疾病。这是一个普遍原则:游戏只在安全、有保障和感觉良好时才会发生,这使得游戏成为衡量所有不良事物的异常敏感的指标。然而,PLAY也是一个强健的系统:如果幼年动物健康且感觉良好,它们几乎总是会在有机会时一起游戏。
目前,人类儿童中一些更喧闹的打闹游戏形式往往被父母所阻止。他们很少考虑这样一个发育事实:身体游戏机会的减少可能会产生不良的成熟后果,例如控制不佳的多动冲动可能变得非常严重以至于被病理化,并且经常被贴上注意力缺陷多动障碍(Attention-Deficit Hyperactivity Disorder, ADHD)等标签(Panksepp,2007b)。表现出过度活动的儿童在服用诸如安非他明等药物后会变得更容易管理,而同样的药物会显著减少大鼠的游戏性(Panksepp, Burgdorf等人,2002)。相反,在ADHD的大鼠模型中,充足的每日游戏可以减轻多动症状(Panksepp等人,2003)。对幼年人类游戏的进一步研究可能使我们能够通过给予被诊断为ADHD的儿童更多的游戏机会来帮助他们,而不是使用抑制游戏冲动的药物。但在我们讨论复杂的社会文化问题之前,让我们先检查表明PLAY是哺乳动物脑心(BrainMind)祖传天赋的证据。
研究表明,游戏的欲望会随着幼年大鼠被剥夺”尽情玩耍”机会的时间延长而系统性增加。实验室大鼠的GRIEF(悲伤)系统相对较弱(可能是由于选择性育种使其能够独自生活良好);这种弱点似乎使它们对PLAY研究特别有用。为了增强动物的PLAY冲动,研究人员必须让它们在社交隔离中待一段时间,这种情况会使许多动物,尤其是灵长类动物,感到孤独和痛苦。灵长类动物有高度发达的GRIEF系统,它们彼此之间有强烈的联结。经过长时间隔离后,幼年猴子会变得沮丧,重聚后它们会挤在一起,最初不愿意游戏(Evans,1967)。显然,它们对社交温暖、支持和归属的基本需求必须得到满足,然后才会再次感到有游戏的心情。在与同类(同一物种的其他个体)相处一段时间后,它们的社交信心得以恢复,沉溺于无忧无虑游戏的冲动重新出现(Chalmove,1978;Novak,1979)。可以预期,人类儿童的行为方式也会大致相同。
然而,幼年大鼠在情感上并不那么依赖。因为它们不会因分离而遭受太多痛苦,玩耍的冲动立即显现出来。经过一段时间的分离(即使只有3到8小时,但最多不超过一整天)后,它们的玩耍系统处于超速运转状态,一旦有玩伴进入活动区域,它们就会迅速进行粗野打闹式的玩耍。即使幼年大鼠从大约2周龄时眼睛和耳朵睁开开始,一直被完全隔离饲养到25天大(此时群居大鼠开始表现出玩耍冲动),它们也不会明显抑郁。相反,它们的玩耍冲动一直在积累,当与另一只大鼠配对时,它们会在几秒钟内以极大的热情进行正常的玩耍(Ikemoto & Panksepp, 1992)。因此,我们可以得出结论,玩耍的冲动不是习得的。它是先天的。证据表明,玩耍是由基因决定的初级过程社交冲动之一。
正如玩耍冲动在社交隔离或其他玩耍剥夺期间系统性地积累一样,当成对的幼年实验大鼠被允许自由玩耍半小时后,玩耍的欲望也会系统性地减少(Burgdorf et al., 2006; Panksepp & Beatty, 1980)。这表明增强的玩耍冲动就像一种饥饿感——对玩耍的特定饥饿感,而不仅仅是一般的社交需求。当幼年大鼠被饲养在可以通过屏障相互接触的环境中时,它们仍然会对玩耍产生渴望,这一事实凸显了这一点。如果大鼠被饲养在繁忙的”攀爬架”类型的生活环境中,它们可以有非常亲密的身体接触,但狭窄的空间禁止粗野打闹,也会出现类似的情况。当被释放到开放的活动区域时,它们会热切地玩耍。同样,与不太爱玩耍的成年大鼠持续生活在一起的幼年大鼠也会积累玩耍欲望。尽管它们有充分的身体接触和许多其他身体及社交互动的机会,但一旦有机会,它们仍会兴致勃勃地玩耍(Hole & Einon, 1984; Panksepp et al., 1984)。
尽管不同哺乳动物物种表现出的特定玩耍模式存在许多差异,但粗野打闹式玩耍在不同物种之间具有动态相似性(Burghardt, 2005; Pellis & Pellis, 2009)。这是一种带有强烈竞争性的欢乐社交活动。
因此,此类活动的进化根源可能要追溯到所有哺乳动物共享的古老同源玩耍回路。尽管不同物种之间玩耍片段的具体细节可能大相径庭,但都暗示着对未来需求的练习。例如,猫等捕食性物种喜欢物体玩耍,如它们拍打众所周知的毛线球所体现的那样,这是对它们成年后所需行为模式的重要练习(Byers & Walker, 1995)。相比之下,羚羊等猎物物种表现出大量的奔跑以及快速的扭转和转向,这些是它们躲避捕食者时所需的技能(Byers, 1997)。除哺乳动物外的其他生物,尤其是鸟类,也可能表现出社交玩耍,但鸟类的玩耍较难预测;它需要大型自由开放的空间,因此更难以进行科学研究(Aldis, 1975)。
大鼠表现出模拟攻击和急切逃避的平衡混合,后者是逃跑/逃避行为。在大鼠玩耍中,人们通常会看到朝向和远离玩伴的快速冲刺活动。有时一只动物会将另一只”撞倒”,这会导致一连串的嬉戏追逐。动物们轮流追逐彼此,伴随着快速的旋转、摔跤和角色转换。它们经常扑到彼此的背上,仿佛在征求激烈的互动。这些背部接触可以很容易地量化,通常被用作表明玩耍冲动的玩耍征求的明确测量指标。通常,玩耍征求的接受者要么逃跑,要么侧向扭转;随后会发生一场摔跤,其中一只动物最终仰面朝天,另一只动物在上面(有关玩耍的非常详细的逐帧分析和许多其他引人入胜的事实,请参见Pellis & Pellis, 2009)。这种压制姿势也可以很容易地量化,是特定玩耍活动完成的最清晰测量指标(见图10.2)。此外,大鼠在玩耍时会发出大量欢快的50-kHz超声波啁啾声;当它们刚开始相互了解时,会发出相对”平坦的叫声”,而在激烈玩耍中则发出更欢快的”调频”叫声。正如稍后将要讨论的,有许多可靠的实证理由相信这是一种祖先的笑声形式,与寻求冲动相关(Panksepp, 2007c; Panksepp & Burgdorf, 2003)。
图10.2. 我们工作中用于量化粗野打闹式玩耍的两种主要玩耍姿势。当动物开始玩耍时,它们首先扑向彼此,尤其是颈背部(背部接触),当它们只是探索新地方时,通常会发出适量的”平坦”55-kHz超声波发声(也称为50-kHz USVs或啁啾声),如右上方的”声谱图”(声音频率随时间变化图)所示。然而,当大鼠真正投入到粗野打闹式社交玩耍中时,它们会四处奔跑,相互追逐和摔跤,最容易测量玩耍的指标是”压制”的次数。在这种欢乐的玩耍中,会有大量调频(FM) USVs,这是这些动物积极情感的直接指标。右下方的声谱图描绘了这些的经典示例,尽管确切的声波波形存在相当大的变化(由Lonnie Rosenberg绘制,发表于Panksepp, 1998a;经牛津大学出版社许可重新发表)。
大鼠的游戏行为在其一生中表现出特征性的发育过程,游戏量在幼年早期增加,在青春期保持稳定,随着动物进入青春期而减少(Barrett & Bateson, 1978; Panksepp, 1981c; Thor & Holloway, 1984a, 1984b)。我们目前对控制这种倒U型成熟功能的神经生物学因素知之甚少。推测这与伴随大脑跨发育成熟的目前未确定的神经化学变化有关(Panksepp et al., 1997)——这种成熟在一定程度上是由游戏期间释放的神经营养因子促进的(Burgdorf et al., 2010; Gordon et al., 2003),这些因子甚至可以促进海马等区域的神经元萌发(Wöhr et al., 2009)。此外,与其他情绪一样,新皮层发育和许多高级脑功能的出现往往会抑制皮层下过程,如引起游戏(PLAY)的过程。这种发育轨迹是合理的。随着动物越来越依赖习得的行为策略,初级过程的冲动会被高级脑功能更好地调节。例如,在成年大鼠中,大的额叶损伤,以及许多高级和低级脑影响进行交流的隔区损伤,会显著增加游戏的冲动(Panksepp et al., 1984, 1994)。这表明这些脑区参与了正常情况下随着动物成熟而减少游戏的发育过程。
如果允许两只大鼠反复一起游戏,游戏优势就会出现(Panksepp, Jalowiec et al., 1985; Pellis & Pellis, 1987)。经过几次游戏后,一只大鼠往往会成为”获胜者”,这意味着它在压制(pins)过程中更频繁地处于上方。平均差异是”获胜者”约70%的时间处于上方,而”失败者”成功率较低,在总压制次数中约30%的时间处于上方。有趣的是,游戏的继续似乎需要较强的伙伴愿意让自己处于不利地位。如果较强的动物不表现出这种互惠性——如果它成为”霸凌者”并渴望一直处于上方——那么游戏活动就会逐渐减少,因为不太成功的动物开始忽视获胜者的邀请。没有人想和霸凌者一起玩。
发育和社会心理学家将人类游戏分为几类:探索性、关系性、建设性、戏剧性/象征性游戏,以及在幼年动物中最容易看到的粗暴游戏(rough-and-tumble play)(Slade & Wolf, 1994)。这些心理学分类法中存在两个常见问题:首先,心理学家经常将游戏(PLAY)与单纯的好奇心混淆——与寻求系统(SEEKING)促进的探究活动的唤起混淆(Welker, 1971; Weisler & McCall, 1976)。其次,许多人将游戏(PLAY)误解为一种攻击形式,这反映在粗暴游戏的常见标签”打斗游戏”中(Aldis, 1975)。尽管很少有人会将游戏(PLAY)视为愤怒系统(RAGE)的表现,但许多物种中常见的争夺优势的竞争类型,特别是在性准备度高时,可能在某种程度上与在幼年游戏期间磨练的行为改进有关,这种观点可能有相当的真实性。然而,正如我们稍后将讨论的,粗暴游戏(PLAY)与任何愤怒型攻击无关,尽管长时间的游戏回合确实经常以一只动物比另一只动物抱怨更多而结束。
让我们考虑第一个问题,即将游戏(PLAY)与好奇心混淆——与寻求系统(SEEKING)的单纯唤起混淆(我们应该注意到,寻求系统显然在游戏期间促进并活跃)。有确凿证据表明游戏(PLAY)和寻求(SEEKING)是不同的,尽管是相互作用的系统。当被放置在新环境中时,动物通常表现出强烈的探索活动,在熟悉周围环境之前几乎没有游戏倾向。神经化学证据也可能支持游戏(PLAY)和寻求(SEEKING)之间的区别。我们已经看到多巴胺为寻求系统(SEEKING)提供燃料,而苯丙胺等精神兴奋剂会强烈增加大脑多巴胺活性。多巴胺活性的增加产生旺盛的探索行为,同时显著减少游戏(Beatty et al., 1982)。然而,阻断多巴胺受体也会减少游戏(Siviy, 2010)。
尽管精神兴奋剂会减少游戏,但多巴胺系统在正常游戏(PLAY)期间仍然被唤起(Panksepp, 1993)。最近的研究表明,大鼠在游戏期间发出的高频(50-kHz)超声啁啾声受到大脑多巴胺唤起的有力促进(Brudzynski et al., 2010; Burgdorf et al., 2001, 2007)。多巴胺在游戏中的作用(可能通过寻求唤起)的进一步支持是各种多巴胺受体阻断剂在减少游戏方面非常有效(Siviy, 2010)。然而,这是可以预料的,因为粗暴游戏涉及大量的来回活动和频繁的愉快期待时刻,而这种情绪状态是由多巴胺推动的。换句话说,多巴胺参与游戏(PLAY)唤起的事实并不意味着多巴胺导致游戏。多巴胺是响应许多积极激励而分泌的,包括游戏机会。因此,游戏期间多巴胺的分泌可能只是表明动物正在从事一项需要大量积极期待和欣快感的活动。
然而,即使多巴胺确实能够积极唤起玩耍冲动,研究也肯定尚未确定在社交玩耍期间是否与非社交探索期间激活的是同一群多巴胺神经元。可能某些类型的多巴胺活动唤起玩耍系统,而其他类型则唤起寻求系统。这些问题只能通过进一步的研究来解决。
多巴胺的作用给我们带来了一个难题:某些多巴胺活动与玩耍相关,但当给动物使用精神兴奋剂(psychostimulants)时,这会大大增强多巴胺活动,玩耍冲动却会减弱。那么,当高水平的多巴胺唤起会减少玩耍时,我们如何理解多巴胺有助于推动玩耍这一事实呢?一种可能是,精神兴奋剂可能会将动态驱动玩耍冲动的情感以持续性(tonic)方式(没有波动)唤起到一个非常高且持续的水平,从而降低了表达玩耍所需的神经化学灵活性。这可能通过”冻结”其正常功能的能力来抑制玩耍。在这些高唤起水平下,玩耍系统可能无法对多巴胺的阶段性(phasic)波动做出动态响应。一个类比可能是试图通过正在发出宽频噪音的扬声器播放音乐。在正常情况下,扬声器传输不同的音乐音调。然而,噪音是一个压倒性的稳定信号,不允许扬声器清晰地传达构成旋律的灵活音调。扬声器类似于玩耍系统,而宽频噪音类似于精神兴奋剂引起的高度且持续的唤起。在这些条件下,导致动态玩耍活动的游戏旋律可能会受阻。但这只是一个想法,目前没有明确的支持。因此,需要考虑所有可能性。另一种可能是,精神兴奋剂只是将动物转移到更强烈的探索性寻求模式,与玩耍动机竞争。
实际上,这可能是许多药物研究中的普遍问题。显然,仅仅应用某些神经活性药物无法模拟相关大脑神经化学系统在正常行为期间的实际运作方式。例如,像安非他明这样的精神兴奋剂药物通常是外周给药—通过注射或口服—这对所有许多大脑多巴胺系统都有大致相同的效果。这会略微增加快乐的50千赫兹鸣叫声。然而,如果将安非他明直接放入大脑的动机区域,特别是多巴胺系统寻求部分的一些主要投射通路(进入伏隔核的一个亚区域—“外壳”,这是中脑边缘多巴胺系统的目标区域,见图3.2),鸣叫会显著增加(Burgdorf等人,2007年;Brudzynski等人,2010年)。相比之下,在伏隔核相邻的”核心”区域效果非常温和,而从其他附近富含多巴胺的区域(如嗅结节和背侧纹状体,后者通常介导习惯性熟练行为)几乎没有任何效果。
除了这些不确定性之外,如前所述,打闹玩耍似乎很容易与攻击性混淆,尤其是对于未经训练的观察者。成年人可能将幼儿的恶作剧视为攻击性,即使孩子们认为这是纯粹的乐趣。尽管玩耍和攻击在表面上可能相似,但仔细审查这两种行为模式会发现许多差异。例如,在真正的打斗中,老鼠经常表现出拳击行为,包括用后腿站立,用前爪互相拍打。有攻击性的老鼠还表现出一种横向的攻击姿态,称为”侧步”,以及竖毛,伴随着许多22千赫兹的”抱怨”声。
动物在玩耍时最初不会发出这样的声音。然而,有时玩耍确实会演变成真正的打斗,导致一些22千赫兹的超声波发声(USVs)。当这种情况发生时,玩耍的迹象—疯狂的跳跃、飞奔和猛扑—立即停止。实际上,在长时间观察期间,玩耍总是系统性地减少,这部分是由于抱怨声(由22千赫兹超声波发声表示)开始增加,而积极的鸣叫相应减少(图10.3)。实际上,如果让小老鼠在测试箱的一侧玩耍前15分钟,在另一个不同的一侧玩耍后15分钟,每只动物都会对它们开始玩耍会话的一侧表现出相当大的偏好。这无疑是因为玩耍会话的后半部分对它们来说不那么愉快。
玩耍与攻击性之间的区别进一步得到以下事实的支持:睾酮促进成年雄性之间的攻击性,而对它们的玩耍冲动影响很小,除了在某些动物中睾酮水平升高会减少玩耍,因为动物更容易陷入真正的打斗(Panksepp,个人观察,1985年)。换句话说,经过几天的高剂量睾酮治疗后,幼年老鼠玩耍较少,显然是因为玩耍回合很快演变成彻底的攻击性,于是社交互动很快失去了无忧无虑的特质。
图 10.3. 这幅图描绘了大鼠在半小时自由玩耍期间的积极和消极情绪发声,类似于男孩和女孩的情况(参见图 10.1)。在这样的时段中,积极的 55-kHz 叫声随着玩耍的减少而系统性地下降。但 22-kHz 超声波发声(可被认为反映消极情绪,即抱怨)则系统性地增加。如果在一个具有明显墙壁的箱子的一侧允许前 15 分钟的玩耍,而在另一侧进行后 15 分钟的玩耍,动物会持续表现出对玩耍开始那一侧的位置偏好,在那一侧观察到大量快乐的超声波发声(数据改编自 Burgdorf 等人,2007)。
人们还观察到,不同的社会规则适用于玩耍和攻击。例如,当成年雄性在其中一只动物的地盘上为争夺统治地位而打斗时,居住方雄性总是获胜。但在玩耍打斗中情况并非如此——居住方”获胜”的次数并不比在中性玩耍室中测试的动物更多(Panksepp 等人,1984)。在玩耍期间,没有持续的防御或攻击姿势,即一只动物长时间仰卧而另一只保持威胁性的上位姿势。但在成年打斗中,这种姿势很常见。在玩耍中,欢快的运动模式会优雅而快速地持续相当长一段时间。然而,正如已经指出的,玩耍并不总是保持情绪上的积极。就像在儿童中一样,可能会出现中断玩耍的争执。然后大鼠会用 22-kHz 痛苦叫声”抱怨”,玩耍随之停止。但通常这只会持续很短的时间,因为大鼠通常会迅速恢复良好的精神状态。然而,在长时间的玩耍过程中,这些抱怨在大鼠中会逐渐增加,因为积极的发声会下降,玩耍的总量会减少,这可能是因为大脑中释放了更多消极的神经化学物质(Burgdorf 等人,2006)。正如我们将看到的,这是在儿童玩耍过程中需要关注的重要问题,特别是如果我们想要促进敏感、具有社交智慧的脑心(BrainMind)的发展。
尽管玩耍可能会以儿童的眼泪和大鼠的发声抱怨告终,但重要的是要强调,对于所有参与者来说,粗暴打闹式的玩耍都是高度有益的;而在真正的打斗中,只有获胜者才能获得暂时的满足。我们如何知道玩耍”打斗”在情感上是积极的?玩耍”打斗”的”获胜者”和”失败者”都能快速学会工具性任务,例如在 T 型迷宫中做出快速而恰当的选择,以获得玩耍的机会(Normansell & Panksepp, 1990; Pellis & McKenna, 1995)。唯一的区别是,获胜者会毫不犹豫地快速冲进”游乐场”,而失败者在进入玩耍场地时会稍微犹豫一些。此外,玩耍场充满了欢快的啁啾声,这些声音已被验证反映了大脑内的积极情感唤起(Burgdorf 等人,2007)。
神经科学家发现,对身体某些区域的挠痒刺激特别容易引起玩耍性——例如,大鼠的后颈和肩部区域。当以恰当的方式触摸这些皮肤部位时,大鼠会发起玩耍。如果这些相同的皮肤部位被麻醉,那么动物的玩耍情绪似乎无法协调,随着皮肤麻醉程度的增加,我们看到的玩耍越来越少(Siviy & Panksepp, 1987a)。然而,麻醉不会减少大鼠相互扑向对方背部的次数——背侧接触的次数是衡量玩耍冲动的良好指标,并没有减少。
然而,对丘脑中处理触觉信息的某些核团的损伤会降低玩耍的冲动。重要的是要注意,大多数感觉系统,包括触觉,在通往大脑高级区域的途中会在丘脑水平分叉,一些”信息”前往新皮质,而其余部分影响大脑较低的网状区域。后者似乎传达感觉输入的情感影响。促进玩耍的触觉成分不是主要前往新皮质,而是前往更古老的中线丘脑区域,如束旁复合体(parafascicular complex)和后背内侧丘脑核。这两者都包含在非特异性丘脑网状核的标题下。对这些大脑区域的双侧损伤,特别是在束旁核中,会减少背侧接触和压制,表明这种大脑损伤确实降低了玩耍的欲望。只有极少量大脑损伤的动物仍然正常参与其他类型的复杂动机行为,如觅食(Siviy & Panksepp, 1985, 1987b)。目前,这些非特异性网状核是我们所拥有的最接近玩耍特异性基质的结构。至关重要的是要记住,我们可以移除整个新皮质,大约是幼年大鼠大脑的四分之一,大鼠仍然会相当正常地玩耍。
显然,游戏动机可能涉及大脑中控制粗暴打闹游戏动作的部分。小脑、基底神经节和前庭系统等大脑区域产生和调节运动。这些区域的损伤会扰乱游戏活动的流程。但这些损伤也会损害几乎所有复杂的运动活动,因此几乎没有机会评估它们的哪些功能是专门用于游戏的。其他大脑损伤会引发抑制游戏的情绪状态。例如,腹内侧下丘脑(VMH)的损伤会导致动物变得病态攻击性,从而减少游戏活动。然而,这并不表明VMH通常促进游戏。它只表明具有攻击性的动物不爱玩耍。
关于杏仁核在游戏中的作用存在一些争议。我们认为,它在游戏动机中的作用是次要的,不是关键的。杏仁核嵌入在颞叶中,当猴子和猫的颞叶被切除时,会出现一种称为克鲁弗-比西综合征(Klüver-Bucy Syndrome)的状况。患有这种综合征的动物往往表现出性欲亢进、口欲亢进(它们会用嘴咬,并且经常吃任何东西),并且几乎不表现出恐惧(Klüver & Bucy, 1939)。然而,这些动物非常渴望玩耍,尽管许多其他社交能力和细微的社交反应已经丧失。然而,Pellis和Pellis(2009)报告称,大脑较大的哺乳动物通常比大脑较小的哺乳动物更爱玩耍,而最爱玩耍的往往有相对较大的杏仁核区域。尽管如此,杏仁核的适度损伤很难发现游戏方面的重大缺陷(Panksepp et al., 1984)。
同样,我们(以及Pellis & Pellis, 2009)注意到,新皮层的参与对于其他六个情绪系统的功能来说都不是必需的,对于游戏系统也不是必需的。去皮层动物大量玩耍,尽管它们被压制的频率低于对照组动物。这并不表明它们不爱玩耍。它们打闹的程度与对照组一样多,只是在嬉戏打闹时不太可能被翻转、被压制。这可能是一种运动副作用,因为去皮层动物总体上似乎更贴近地面。但这也可能是由于这些动物相对缺乏社交敏感性。完整的动物能够敏感地意识到,除非其他动物有机会偶尔获胜,否则它们不会想玩。如上所述,较强的动物会限制自己,以保持游戏的乐趣。可以推测,这种敏感性需要更高级的大脑参与水平,而去皮层动物显然不具备这种能力。尽管如此,如果观察这些动物相互对抗时的游戏主导地位,神经系统完整的大鼠并不总是战胜去皮层大鼠。实际上,结果基本持平,每种动物大约赢一半的时间(Panksepp et al., 1994)。
尽管我们对游戏回路的神经解剖学理解仍处于初级阶段,但我们可以确定这种初级过程情绪是在哺乳动物大脑的皮层下水平组织的。这并不意味着高级大脑区域在游戏中没有功能。它们当然有功能,尤其是在象征性游戏中,这种游戏让我们许多人喜欢演奏乐器、参与戏剧制作和电影表演,最重要的是发明可以为生活增添无尽乐趣和兴奋的游戏。所有这些都深刻依赖于皮层对来自下方的原始信息的专注倾听。因此,越来越多的证据表明,原始的游戏冲动是帮助编程高级大脑区域的重要影响因素——使其成为充满创造力和生活热情的快乐成年大脑。
由于大多数幼年哺乳动物表现出从事粗暴打闹游戏的冲动,我们认为这是最基本的游戏形式。更复杂的人类游戏形式可能是我们与其他动物共有的原始粗暴打闹冲动的次级和三级过程变体。成年人的游戏冲动可以通过多种方式表现出来。随着个体的成熟,人类的大量游戏开始集中在语言交流上。经常表现友好戏弄的持续言语交锋与粗暴打闹游戏中的背部接触和压制有明显的相似之处。一个人试图用某种挑衅来激起另一个人,有时甚至是尖锐和刻薄的评论;然后,如果其他人回应,通常会希望用特别聪明的回应”回击他们”。如果成功,这会在年轻人中引发阵阵笑声,在老年人中引发轻笑。这种类型的交锋可能会重复多次,每个人都试图超越对方——成为最聪明的人——直到明确一方获胜或每个人都满意地认为自己是对方的对手。当后者发生时,这些人可能很有潜力建立特殊的尊重和友谊。
在回到我们游戏冲动的一些更高级功能之前——这使许多社交互动感觉特别积极——让我们暂停重新考虑各种感官如何控制基本的游戏冲动。例如,失明的大鼠会精力充沛地玩耍。尽管大鼠不需要视觉,但失明确实会减少像我们这样视觉导向生物的游戏活动。尽管如此,盲童和能看见的孩子一样喜欢玩耍,并且在玩耍时很容易笑。嗅觉能力受损的大鼠几乎可以正常玩耍。毫无疑问,鼻塞的儿童也能很好地玩耍,尽管没有人正式研究过这一点。简而言之,视觉和嗅觉在游戏冲动中都不起关键作用。
耳聋的老鼠玩耍的时间比听力正常的老鼠略少,这可能是因为它们对超声波老鼠”笑声”不敏感。然而,正如已经指出的,引发和维持玩耍的主要感觉系统是触觉。重申一下,有两条主要的触觉通路。通往新皮层的特定通路携带关于触觉刺激的认知信息(例如,你被触摸的位置),对玩耍并不是至关重要的。然而,通过丘脑非特定网状核运行的通路携带触觉引起的情感体验,它们对打闹玩耍非常重要(Siviy & Panksepp, 1987a, 1987b)。玩耍引发区域被麻醉的动物根本无法协调它们的玩耍冲动。触觉和玩耍冲动重合的事实表明,某些类型的触觉会产生情感体验,这对唤起玩耍的欲望非常重要。我们在挠痒痒游戏中最清楚地看到这一点。
这些发现表明,老鼠拥有专门的皮肤区域,当它们被触摸时会向玩耍系统发送信号。此外,人类更容易在老鼠的颈部和肩部挠痒痒,而不是在后腿部位。换句话说,老鼠似乎有”玩耍皮肤”或”挠痒痒皮肤”,拥有专门的受体将信息发送到大脑的特定部位,这些部位接收动物之间玩耍意图的交流。显然,人类也有挠痒痒皮肤,它位于颈部后面和肋骨周围。当然,这是最容易挠痒痒小孩子的地方之一,会产生玩耍的心情。
显然,玩耍系统也会对不可预测的刺激感知进行调整。例如,人无法给自己挠痒痒。此外,底层神经系统的设计使得儿童不能轻易成为自己打闹玩耍的伙伴(尽管存在独自探索和幻想玩耍的形式)。挠痒痒需要他人参与才能唤起玩耍性。这不仅仅是一种感觉现象,而是非常重要的内部大脑功能。
许多人认为笑是人类现象,它总是与幽默相关,比如笑话的结尾。然而,笑并不需要太多认知复杂性。例如,孩子们喜欢表演短剧和节目,但当他们试图认真表演时,往往最终会高兴地咯咯笑起来。儿童笑声的最高水平出现在孩子们进行身体玩耍时。显然,即使是人类的笑声也根植于古老的玩耍系统,这些系统在其他哺乳动物中产生快乐的社交互动。
现在有强有力的证据表明,除了人类之外,许多其他哺乳动物也会发出类似笑声的声音。最有力的证据来自人类给其他动物挠痒痒。正如身体挠痒痒是引发小孩子笑声的最简单方法之一,事实证明许多其他动物也是如此,从所有大猿到实验室老鼠。人们早就知道,通过挠痒痒黑猩猩和大猩猩,可以诱发类似人类笑声的循环模式的喘息和咕噜声(Provine, 2000)。事实上,最近的一项研究(Ross et al., 2009)直接对比了所有大猿中挠痒痒诱发的声音,作者断言
至少可以得出结论,将”笑声”视为跨物种现象是合适的,因此使用这个术语来描述大猿产生的挠痒痒诱发的声音并不是拟人化。这个术语已经在以前关于几种非人类物种的挠痒痒和玩耍相关声音的研究中使用过……目前的结果为这种用法提供了明确的支持。(第1109页)
对许多人来说,令人惊讶的是笑的概念可以扩展到像老鼠这样低等的哺乳动物物种。许多严肃的神经科学家没有准备好接受这一发现(概述见Panksepp, 2007c, 2010d; Panksepp & Burgdorf, 2003, 2010)。为了评估老鼠是否会笑,我们使用了一种简单的挠痒痒方法—基本上是人类手部玩耍—这诱发了持续的高频啁啾声(约50千赫兹,远超人类听力范围,因此需要特殊的声谱测量)。事实上,通过挠痒痒老鼠通常寻求玩耍的那些身体区域—特别是颈背部—可以最有效地唤起这种啁啾声。当然,全身挠痒痒更有效。就像孩子一样,老鼠喜欢它!因为在老鼠的自然玩耍中这些相同的啁啾声非常丰富,如果我们认为它们与我们自己的笑声没有关系,那就太冷酷无情了。事实上,就像给小孩挠过痒痒之后,仅仅通过”威胁”挠痒痒就可以得到阵阵笑声。在老鼠身上也看到了非常相似的反应:仅仅通过接近的手指动作就会开始产生笑声(Panksepp & Burgdorf, 1999)。
但我们不必仅仅依靠行为研究来识别这种与人类笑声的联系,因为已经对老鼠的笑声进行了严肃的脑研究。通过使用局部脑刺激来唤起笑声型啁啾声,已经在老鼠身上绘制了笑声回路。该回路沿着中脑边缘系统的寻求系统(SEEKING system)运行,并受多巴胺的强烈控制;无论我们在哪里找到笑声点,幼年老鼠都会readily自我刺激——自愿”充电”——这些大脑部位(见图10.4,总结了Burgdorf等人2007年的绘图工作)。虽然我们对产生初级过程人类笑声的皮层下区域仍有很多需要了解的,但大量研究发现表明,与老鼠笑声相关的大脑区域在产生人类笑声方面也起着重要作用(Black, 1982; Chen & Forster, 1973; Poeck, 1969; Sterns, 1972; Wild et al., 2003)。因此,现有证据与人类和老鼠笑声由进化相关的皮层下回路产生的可能性一致。
在上一章中,我们说过痛苦的发声是唤起悲伤(GRIEF)的有用指标。同样,笑声型啁啾声是老鼠唤起玩耍性的主要指标。笑声,就像玩耍本身一样,是一种无条件的本能反应,在适当的社会环境条件下,从哺乳动物大脑的古老区域产生。它不是通过模仿学习的,因为盲人和聋哑儿童很容易笑(Eibl-Eibesfeldt, 1989)。我们很容易培育出老鼠的挠痒诱导啁啾反应(Burgdorf et al., 2005)。根据我们采取的所有措施,表现出丰富啁啾声的动物更快乐,而那些不太啁啾的动物表现出负面情绪和易患抑郁症(Brudzynski et al., 2010; Burgdorf, Panksepp et al., 2008; Harmon et al., 2008)。如果这些发声模式在哺乳动物物种之间确实是同源的,我们最终可能通过研究在老鼠身上产生欣快啁啾声的回路来了解人类快乐的原始本质(Panksepp & Burgdorf, 2003; Panksepp, 2007c)。
当然,这种分析的底线将是我们是否能够识别出有助于构建和作用于特定神经化学回路的主要基因,这些回路使动物能够玩耍。我们在啮齿动物模型上正在取得进展,我们将在本章后面部分讲述这一进展的一些细节。这种工作可以为笑声的进化连续性和社会快乐在不同物种之间的本质提供严格的神经进化证据,无论是支持还是反对(Panksepp, 2007c)。简而言之,对老鼠的研究可能比科学家可用的任何其他策略都更能告诉我们人类玩耍(PLAY)和笑声的原始本质。
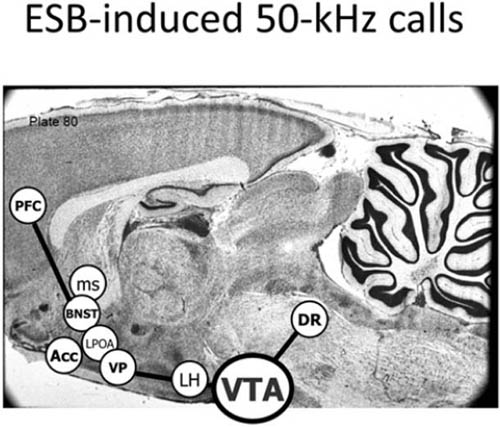
图10.4. 在老鼠的局部电刺激大脑期间,可以从哪些大脑区域唤起丰富的频率调制(FM)型50-kHz超声波发声(USVs)的总结。这些往往遵循寻求系统(SEEKING system)的轨迹。在每个可以唤起这种叫声的地方,动物也会自我刺激电极部位,多巴胺阻断药物选择性地减少这些叫声。从下到上描绘的解剖区域是中缝背核(DR)、腹侧被盖区(VTA)——它与外侧下丘脑(LH)一起可能是最有效的部位——腹侧苍白球(VP)、伏隔核(Acc)、外侧视前区(LPOA)、终纹床核(BNST)、内侧隔区(ms)和内侧前额叶皮层(PFC)(Burgdorf等人2007年报告的数据摘要;我们感谢Jeff Burgdorf分享这个摘要)。
我们现在对老鼠笑声了解得非常多,当然比我们对初级过程人类笑声的了解要多得多。就像小孩子一样,挠痒痒对幼年老鼠来说是一种积极的激励。它们寻求这种刺激,当收到与挠痒痒相关的提示时,会迅速开始啁啾。人们还可以通过直接刺激许多大脑部位来唤起啁啾声。重要的是要强调这样一个事实,即在老鼠身上可以唤起啁啾声的每个大脑位置都支持自我刺激——动物很容易打开大脑刺激,表明与这些声音相关的体验是积极的(Burgdorf et al., 2007)。以这种方式自我刺激的准备程度表明这些部位提供情感愉悦,啁啾是老鼠正在经历愉快体验的信号。简而言之,就像我们自己的孩子一样,幼年老鼠很容易学会享受挠痒痒。这个笑声系统主要遵循寻求系统(SEEKING system)的事实也有助于解释为什么当动物和儿童期待非社交性奖励时会出现这种快乐的声音(Knutson et al., 2002)。
因此,50-kHz叫声在动物对情感上令人愉悦的药物成瘾时大量出现也就不足为奇了(Burgdorf et al., 2001; Knutson et al., 1999)。事实上,这种测量可以用作药物渴望的自我报告(Browning, et al., 2011; Panksepp, Knutson et al., 2002)。同样,在雄性和雌性大鼠的性唤起期间,都会发出50-kHz叫声,特别是在交配前的”求偶-邀请”兴奋阶段(McGinnis & Vakulenko, 2003)。正如我们之前提到的,有些令人惊讶的是,当交配结束时,雄性大鼠开始发出22-kHz警报型叫声,与在真正危险情况下发出的叫声只有轻微差异;也就是说,交配后的叫声不像警报叫声那样单调平坦,而是有频率调制的波纹(Burgdorf, Kroes et al., 2008)。我们之前提到,雄性大鼠可能只是在告诉雌性现在离远一点,因为它已经相当满足并准备梳理自己了。更有趣的是,也许这种通常用作警报的发声,可能有助于让其他雄性保持距离。在像大鼠这样的滥交物种中,精子竞争是决定谁真正繁殖的重要因素。发出伪警报叫声将是一种了不起的进化适应,有助于确保其他雄性大鼠保持距离,从而增加自己的精子实现受精目标的可能性。如果过度拟人化,也许这是一种欺骗,在附近其他雄性眼前”蒙混过关”,而这些雄性本来会迫不及待地成为下一个与性接受雌性交配的对象。
在人类中,人类笑声的阴暗面长期以来一直被认为是对看到他人受伤、羞辱或尴尬的反应(可以说是滑稽幽默)。阴暗的笑声认识到受害者处境的荒谬性,同时感觉自己在心理上比那些不幸的人更幸运,甚至可能更聪明。这种场景经常在戏剧喜剧中被利用。值得注意的是,在人类儿童的竞争性游戏互动中,明显的胜利者总是比失败者表现出更多的笑声,尽管这种效应可能还没有得到科学记录。同样,恶作剧的实施者比受害者更有可能笑。
这些模式表明,笑声经常被竞争性,甚至可能是攻击性的冲动所利用。它可以用来对对手造成情感伤害。也许这种更高形式的窃笑只能发生在像灵长类这样复杂的认知生物身上,它们可以将自己的心智能力用于多种目的。这将接近弗洛伊德对幽默的经典解释,即幽默是原本不可接受的性和攻击冲动的可接受面纱(Freud, 1905a/1968)。但在其他动物身上没有这种过程的证据。神经科学研究表明,服务于攻击的笑声不是初级过程游戏系统(PLAY system)的内在方面。那是一种更高级的心智功能。
通过药理学操作抑制游戏是非常容易的。然而,很难确定药物的抑制作用是否包括对游戏系统的特定改变,还是仅仅由于焦虑增加、认知干扰或镇静等引起的普遍行为破坏。脑成像研究表明,在游戏过程中,神经系统中有广泛的阿片类物质释放,特别是在视前区(POA,它也控制性行为和母性行为;见第7章和第8章)。这些发现表明,阿片类物质释放可能在唤起游戏系统中发挥积极作用(Panksepp & Bishop, 1981; Vanderschuren et al., 1995)。此外,随着大脑中内源性大麻素系统的发现,以及长期以来文化上认识到大麻可以引起咯咯笑,促进大脑中”大麻样”活动确实能促进大鼠的游戏性也就不足为奇了(Trezza & Vanderschuren, 2008, 2009)。
阿片类物质活动在游戏中的作用现在已经在动物研究中得到广泛研究。非常低剂量的吗啡实际上会促进游戏性和社会支配地位(Panksepp, Jalowiec et al., 1985; Vanderschuren, 2010),并且也控制游戏支配地位。众所周知,在预测两只动物之间游戏互动的”赢家”时,动物体重的增加具有明显优势,就像男孩之间的摔跤一样。但大脑神经化学活动也会产生重大影响。研究发现,在两只体力相当的动物之间,如果一只被给予小剂量的阿片受体刺激剂如吗啡,而另一只被给予等量剂量的阿片受体拮抗剂如纳洛酮,接受吗啡的动物总是成为赢家(Panksepp, Jalowiec et al., 1985)。这些结果表明,较高水平的大脑阿片类物质,足以产生社会自信感(即如第9章所讨论的,减少分离痛苦),有助于在游戏竞争中获胜。另一方面,低水平的大脑阿片类物质活动会产生更大的社会需求感,因此产生不安全感。这使动物处于情感劣势,使它们更有可能失败。当然,为了促进游戏和自信,阿片类物质剂量必须很低。高剂量会使动物镇静并减少包括游戏在内的所有社会行为,非常高的剂量会引起紧张性僵直。
正如行为和心理科学中的所有发现一样,这些结果也存在其他解释。例如,阿片受体拮抗剂如纳洛酮可能会降低游戏性,但它们也可能只是减少了通常在各种友好社交互动中产生的积极感受。另一种可能性是阿片类物质可以减轻疼痛,因此接受纳洛酮的动物可能会感到游戏过程中一些较粗野的活动比接受吗啡的动物更不愉快。无论如何解释,阿片操纵对游戏主导地位的影响在那些在相互游戏体验开始时就接受这些药物的动物身上都非常稳定。然而,如果在这种药理学操纵之前,一对游戏伙伴的社会关系中已经建立了主导模式,那么主导模式就不会因这些神经化学变化而轻易改变。这种结果的差异表明,过去的社会学习对游戏行为施加了强大的影响。
社交剥夺是另一个增加游戏欲望的因素,这表明应该可以人为地增加游戏欲望。大脑中可能存在高度特异性的促进游戏的神经化学物质,也许是神经肽(neuropeptides)。然而,尚未确定这类物质,尽管一些候选物质正在从基因研究中出现(如下文所述)。寻找相关证据的部分困难在于,几乎所有神经肽都必须直接注射到大脑中,而我们对游戏回路的了解还不够充分,无法将这些物质放置在适当的区域。不过,我们已经评估了一些神经肽的影响,包括催产素和CRF,我们发现这两者都会减少游戏;我们还发现加压素对游戏的影响不太明显(Panksepp, Crepeau et al., 1987)。我们仍在寻找能够”开启”那些心理上还没准备好游戏的动物的游戏性的神经化学系统。到目前为止,这项努力还不太成功。也许需要的是一系列神经化学变化以正确的模式同时发生。只有当我们理解了动物进行游戏活动时演奏的神经化学旋律,我们才能开始对哺乳动物大脑中的游戏性有深刻的神经理解。其他神经化学系统肯定会被发现,并对游戏产生更特异的影响。
在这方面正在取得一些进展。在研究游戏产生的大脑基因表达模式时,发现胰岛素样生长因子-1(IGF-1)和谷氨酸受体亚型的升高,进而进行的行为研究表明,促进这种增加的分子确实也能促进游戏性(Burgdorf et al., 2010)。事实上,初步数据显示,游戏性可以逆转抑郁症状,包括增强经常被压力损害的大脑区域(Wöhr et al., 2009)。我们正在使用上述策略顺利识别新的抗抑郁分子,一种甘氨酸位点谷氨酸受体调节剂可以刺激(低剂量)和阻断(高剂量),从而温和地增加积极情感并减少消极情感,目前已进入临床试验(Burgdorf et al., 2011)。
总之,非常明确的是,游戏(PLAY)是大脑中一个非常有益的过程。当研究人员研究调节游戏的神经化学时,他们也发展出了关于哪些大脑化学物质在产生社会奖励中很重要的想法。第一批看起来重要的化学物质是内源性阿片类物质,它们在游戏过程中分泌(Panksepp & Bishop, 1981; Vanderschuren et al., 1995)。考虑到寻求(SEEKING)冲动在游戏过程中可能非常活跃,而多巴胺似乎介导大脑中的欣快感,多巴胺也参与游戏的欣快方面的可能性很高。此外,越来越清楚的是,内源性大麻素似乎也促进大脑中其他形式的积极情感,它们是游戏奖励包的重要组成部分(Trezza & Vanderschuren, 2008, 2009)。最后,新的基因发现方法开始产生大脑中其他介导游戏奖励的分子路径(Burgdorf et al., 2010)。
许多研究人员和理论家都考虑过游戏可能具有什么功能。建议分为两大类:社会性和非社会性。可能的社会功能包括学习各种竞争性和非竞争性的社交技能。这些技能范围从促进社会联结和社会合作的行为,到促进社会等级和领导力的行为,以及有效沟通的能力。游戏的潜在非社会功能包括学习或增强诸如体能、认知功能、熟练使用工具以及在意外事件面前进行创新的能力等资产和能力(Spinka et al., 2001)。非社会功能的范围可以从复杂的认知技能(如在各种情况下创造性思考的能力),到非常具体的能力(如幼年捕食者学习狩猎和幼年猎物学习躲避捕食者所获得的技能)。不幸的是,对于这些想法中的任何一个,都没有大量实质性的科学数据库。
当然,玩耍以各种方式提高繁殖适应性,但在大鼠的粗暴打闹游戏过程中,性行为类型的行为非常少见,尽管其他各种动物在玩耍期间表现出相当多的骑跨行为。人们可能期望在幼年发育期间没有玩耍的动物在成年性行为方面会处于劣势。事实上,在整个幼年期(21-45天大)被社交剥夺的雄性大鼠,当置于激素刺激的雌性面前时,表现出相当正常的初级过程性行为。然而,我们在一些未发表的研究中发现,在包括两只雄性和仅一只可接受雌性的竞争情况下,有玩耍经验的动物在阻止那些很少进行幼年玩耍的动物的进攻方面更有效。因此,幼年玩耍经验似乎确实可能在竞争获得繁殖机会方面赋予优势。
与几乎没有玩耍历史的动物相比,那些有丰富玩耍经验历史的动物花更多时间与其他动物在一起,这表明玩耍具有社交联结(友谊)功能。事实上,幼年动物喜欢花更多时间与仍然大量鸣叫的年长动物在一起(例如,我们上面描述的高鸣叫系动物),而不是那些不太鸣叫的动物(见图10.5)。这种功能对于建立未来的社交联盟和合作,甚至可能是共情,都非常有用。在认知方面,一些研究者报告说,有玩耍经验的大鼠在非社交问题解决方面有所提高。但我们在重复这些结果时遇到了反复的困难。另一方面,社交效应更容易记录。被剥夺玩耍的大鼠在各种社交情况下往往更恐惧,当然也更具攻击性(Potegal & Einon, 1989)。尽管在这些问题上需要更多数据,但似乎玩耍很少的动物往往更易怒,社交创造力更低。
PLAY(玩耍)系统似乎有助于磨练广泛的社交和非社交技能——它是一个经验期待过程(experience-expectant process),为动物未来的挑战做准备。关于这如何发生的数据很少。但让我们发挥创造力。PLAY可能与做梦的功能有关吗?玩耍和做梦似乎都是大脑的经验期待功能,旨在评估过去的事件作为创造性和有用的未来行为的来源。也许玩耍的功能与做梦是互补的。两者都可能帮助以促进对未来生活事件的高阶情感反应的方式组织大脑中的信息。换句话说,也许玩耍和做梦都允许动物测试他们在现实生活中遇到的复杂问题的解决方案。如果是这样,我们怀疑玩耍性(playfulness)在心理治疗中应该比目前发挥更大的作用。但让我们首先看看睡眠本身的现象。
图10.5. 幼年大鼠可以选择去T型迷宫的两侧,每一端都有一只成年雄性大鼠,它们的社交气质不同。一侧有一只表现出大量50-kHz鸣叫的动物,另一侧有一只表现出低鸣叫的动物。在半小时的测试期间,幼年动物明显更喜欢与”更快乐”的成年动物在一起,这种效应在整个测试期间变得更大(数据来自Panksepp & Burgdorf,未发表数据)。
神经科学家现在知道,哺乳动物的大脑包含内源性每日节律生成器——昼夜节律时钟(circadian clocks),在一天中产生清醒和睡眠时期。睡眠的主要昼夜节律时钟位于视交叉上核(suprachiasmic nuclei, SCN)的神经元中,位于大脑底部,就在视交叉上方,视神经在那里交叉,为两个大脑半球提供来自每只眼睛的视觉信息。这些SCN神经元对化学物质褪黑素(melatonin)特别敏感,并且对光也很敏感。褪黑素从松果体(pineal gland)分泌,强烈影响视交叉上核:当光线减弱时,这会促进人类和大多数动物的睡眠(大鼠是夜行性的,但在夜幕降临后褪黑素水平也较高,这表明黑暗驱动褪黑素的产生)。
哺乳动物的两种主要睡眠状态是:(1) 慢波睡眠(SWS,也常被称为非快速眼动睡眠,或NREM),通常是无梦的;(2) 快速眼动睡眠(REM),在此期间人和动物会做生动的梦。虽然快速眼动睡眠和做梦确实有不同的大脑机制,但两者通常协调良好。腹外侧视前区的一个非常特定的部位已被确定为重要的慢波睡眠生成器,尽管很明显新皮层也有内在的慢波睡眠生成器——在大脑的较高部分,睡眠在一定程度上是区域性的(Krueger等人,2008)。快速眼动睡眠是由脑干中位于中脑下方较低位置的区域生成的。在快速眼动睡眠期间,会有持续的肌肉松弛,这通常可以防止动物将梦境付诸行动。因此,大的抗重力肌肉在快速眼动期间保持放松,在全身产生肌张力缺失。但在做梦期间也有各种阶段性成分的爆发,这反映在各种小的肌肉抽搐中,其中研究最充分的是快速眼动,这种”矛盾”的睡眠阶段就是因此而得名的。当人和动物处于快速眼动睡眠时,他们还会移动手指、嘴唇、鼻子、脚趾、中耳的各种肌肉等等。关键在于这些外周抽搐不会导致任何协调的全身行为。快速眼动睡眠期间的肌肉抽搐在典型的脑电图读数中表现为巨大的放电峰值,特别是在视觉系统内。
生成慢波睡眠和快速眼动睡眠以及清醒状态的关键大脑结构位于脑干深处。慢波睡眠机制位于脑干的较高位置,基本的清醒机制位于脑干网状结构的较低位置,而快速眼动生成器则位于更低的位置。因此,哺乳动物有一种不寻常的大脑排列,特别是考虑到大脑的较高区域通常比较低区域进化得更晚。生成慢波睡眠最有影响力的机制位于大脑中比让我们保持清醒的基本神经系统更高的区域。快速眼动的执行机制位于三者中最低、也许是最古老的位置。如果我们接受位于中枢神经系统较低位置的结构通常比位于较高位置的结构更原始,那么我们需要理解这样一个事实:哺乳动物大脑的主要清醒机制比基本的快速眼动-梦境生成器进化得更晚。然而,我们必须对这种明显的倒置持保留态度,因为越来越多的证据表明,做梦的体验来自比快速眼动睡眠更高的大脑区域(Solms,2000)。特别是,现在的证据表明,多巴胺介导的SEEKING唤醒可能对梦境生成非常重要(Léger等人,2010;Léna等人,2005)。
尽管存在这些模糊性,我们仍然需要面对我们自己大脑中看似颠倒的状况:快速眼动唤醒网络显然比我们的脑干清醒系统更古老。为了理解这个悖论,我们可以考虑一种原始形式的情绪清醒可能在我们与新皮层功能相关的那种清醒(伴随所有感官意识和思维)之前就进化出来了。换句话说,在古老的进化历史中,原始的初级过程意识最初可能仅作为一种做梦式的清醒而存在——一种充满情绪唤醒的状态。这种简单的情感清醒可能已被更具认知性的心智框架所取代。但情绪唤醒可能仍在快速眼动睡眠期间占主导地位。因此,人类和动物心智的梦境内容可能会随着其主导的情绪唤醒而变化。为了让大脑的较高认知区域在学习和思考中变得最有用,在做梦睡眠的安全环境中锻炼与情绪相关的认知可能性可能很重要,从而也许能更好地帮助整合认知和情感问题。在睡眠期间,这种情绪唤醒隐藏在称为快速眼动-肌张力缺失的运动麻痹之后,但它们并未完全消除,在肢体的许多抽搐中仍然明显。也许我们情感丰富的梦境生活是这种双重心智逐步进化的残余;一开始这种唤醒可能主要是情感性的,但随着大脑的扩展,它们在古老的情感过程和更现代的认知过程之间达到了平衡。这可能有助于复杂的问题解决(Levin等人,2008)。
古老的情感意识形式在大脑进化早期占主导地位这一观点得到了以下事实的支持:当肌张力缺失的大脑机制被选择性破坏时,动物仍然表现出有规律的做梦型睡眠期。这些动物会将梦境付诸行动,因为它们的大抗重力肌肉不再变得如此松弛以至于无法移动。这些奇怪的”梦境”时期为观察它们的、也许还有我们自己的古老情绪心智提供了一个窗口。例如,那些内眼睑(瞬膜)在这些付诸行动的梦中保持闭合的猫(使它们基本上处于失明状态),表现出四种主要的行为模式:捕食性跟踪、恐惧、明显愤怒的猛击,以及周期性的梳理行为。
我们的解释是,当高度情感性的、非反思性的、梦境类型的意识首次进化出来时,伴随着丰富的情感活动,这些情感活动的自由表达在进化过程中逐渐被抑制和调节。这是因为这种简单的生存解决方案不再像以前那样具有适应性。哺乳动物中更高级、更认知的大脑区域的大规模扩张,可能需要大脑中新的觉醒机制的进化,以帮助维持新皮质区域的清醒状态,从而让更高级、更认知的意识形式得以出现。因此,那些控制简单情感觉醒的更古老的大脑觉醒机制(可能是爬行动物所需要的全部),将逐渐被抑制和重塑,成为控制快速眼动睡眠觉醒的机制。通过这种方式,梦境可能仍然主要由古老的情感觉醒状态控制,但在更现代的动物中,这些状态允许认知信息与情感压力性觉醒更好地整合。这可能是一种方式,让古老的心智框架仍然能够调节更近期进化动物的更高级信息处理。
除了它在脑干中的深层位置之外,还有另一个理由认为快速眼动睡眠可能是一种原始的清醒状态类型,因为它不再能最优地促进生存而被抑制:快速眼动睡眠存在于哺乳动物中,但不存在于爬行动物或鱼类中。鸟类仅表现出适度的快速眼动睡眠期,每次只持续几秒钟。似乎不太可能产生快速眼动睡眠的大脑机制是在哺乳动物中独特进化出来的,而不是从预先存在的古老大脑功能中出现的。在大脑进化的早期阶段,动物肯定有更简单的意识形式,随着大脑的更高级进化,这些古老的解决方案必须与最近进化出的大脑功能整合。有可能在清醒时,游戏是以类似于睡眠期间做梦的方式促进这种整合的大脑系统。
总之,进化中的哺乳动物心脑向认知复杂性的投射需要一个重大的进化步骤:它需要构建新的觉醒系统来调节丘脑和新皮质的清醒状态,以及一个新系统来抑制简单的情绪性(现在表现为快速眼动活动)。我们知道这两个系统都存在于哺乳动物的大脑中。唤醒皮质的系统包括生物胺(多巴胺、血清素和去甲肾上腺素)以及产生乙酰胆碱的细胞群,位于上行网状激活系统中,该系统位于脑桥的上部区域。此外,各种神经肽神经元,如集中在下丘脑外侧等更高脑区的食欲素(orexin,也称为下丘脑分泌素hypocretin)神经元,对于从慢波睡眠平稳过渡到快速眼动睡眠是必需的;没有它们,动物和人类会表现出发作性睡病(narcolepsy),即从清醒状态突然崩溃进入快速眼动状态。在快速眼动睡眠期间产生肌肉无力的大脑部分也不同于产生快速眼动睡眠中充满情感的阶段性活动的部分。它位于蓝斑核正下方——蓝斑核是大脑中最大的去甲肾上腺素细胞群,它在整个皮质中促进觉醒,在情绪状态期间尤其强烈(见图1.1)。
为什么我们在游戏章节的背景下考虑这些深奥的问题?我们设想游戏和快速眼动睡眠之间可能存在联系:如果我们认为快速眼动睡眠在哺乳动物大脑中的一个关键功能是促进复杂情感信息的整合是正确的,那么游戏系统可能在清醒生活中执行类似的功能。我们提出这种可能性是因为,在游戏中,许多类型的情感行为在非严肃的互动背景下表现出来。支持这一观点的事实是,快速眼动睡眠和游戏都受到乙酰胆碱、多巴胺、去甲肾上腺素和血清素等神经递质的严格控制。相似的化学介质可能表明相似的功能。游戏可能在社会快乐感的影响下,巩固不同情绪的多样化行为成分,让儿童逐渐发展出对其物理和社会环境做出反应的习惯性创造性和积极方式。游戏冲动可能对复杂社会大脑的文化和表观遗传构建至关重要,这种大脑能够理解他人的情感状态和动机,为复杂的社会合作以及对他人的友情、同情、共情和团结的同伴情感打开大门。游戏促进社会智能。
快速眼动梦境可能对侵入并赋予每个生命意义的多样化初级过程情感成分发挥非常相似的功能。换句话说,做梦和游戏在心智生活的表观遗传创造中可能具有协同功能。正如我们之前所指出的,现在很清楚,只有少数更高级的心智功能是由我们心脑的更高认知区域中的进化遗产赋予的。大多数是在强烈的文化影响下学习的。我们在这里关注的基本情绪系统都参与构建我们的认知优势和劣势,以及将我们每个人巩固为独特的人格。我们的情感潜力与认知能力的整合是由每个人独特的发展景观的魔力创造的。
表观遗传原则(关于这个概念的讨论,请参见[第6章]和[9])在考虑PLAY系统时特别适用。尽管新皮层不是产生PLAY的必要参与者,但玩耍性对新皮层产生特别强的影响,导致基因表达谱的许多变化。当儿童玩耍时,他们的活动促进了这个器官的表观遗传变化。神经元代谢的脑成像显示,当动物玩耍时,新皮层和许多皮层下区域都有高水平的活动(见Panksepp, 1998a, Fig. 15.7; Gordon et al., 2002)。研究表明,社交游戏还激活了大脑某些区域的神经生长因子(如脑源性神经营养因子[BDNF]),最明显的是在额叶皮层和杏仁核(Gordon et al., 2003)。但BDNF遍布整个大脑,因此大脑效应非常广泛,显然在某些回路中促进积极情感,可能在其他回路中促进消极情感。
图10.6. BDNF基因表达图片(通过原位杂交),冠状鼠脑切片(从前到后;A–E),在有和没有半小时社交游戏的动物中。如直方图所示,这种广泛的神经元生长因子在额叶皮层和杏仁核中升高(数据来自Gordon et al., 2004)。
最近一项更全面的大脑基因表达分析表明,我们在额叶皮层区域评估的约1,200个大脑基因中,大约三分之一的活动会被游戏迅速改变(Burgdorf et al., 2010)。可以暂时合理地假设,游戏引起的动态大脑变化促进大脑生长和成熟,可能通过表观遗传创建大脑的亲社会回路,可能部分通过完善额叶执行功能(见[图10.7])。我们最近确定了促进玩耍性和积极情感的分子途径,大脑”肥料”IGF-1(胰岛素样生长因子)在游戏期间高度激活。IGF-1已被证明是大脑中的积极享乐分子(Burgdorf et al., 2010)。因此,PLAY活动的效应之一越来越可能是通过表观遗传在新皮层中创建新的亲社会神经通路—作为经验函数的基因表达模式的长期修改。
PLAY如何帮助编程新皮层?游戏的困境之一是它将儿童带到他们情感知识的边缘。当这种情况发生时,不可避免地会出现冲突的情感感受,需要在关爱成人的帮助下立即解决。自由游戏期间会发生不好的事情,如果没有监督,游戏很容易导致一个孩子欺负另一个孩子以获得社会主导地位。然而,在成人的关注下,每一个这样的冲突时刻都成为积极的亲社会学习的绝佳机会。因此,在我们这个幼儿活动比以往任何时候都更受控制的时代,我们确实需要有爱心的人监督游乐场,以便他们可以在这些关键时刻温和地教育。
图10.7. 额叶功能的概要总览,这些功能在被诊断为注意力缺陷多动障碍(Attention-Deficit Hyperactivity Disorder)的儿童中可能成熟缓慢(改编自Panksepp, 2007)。
在我们对人类社交游戏进行首次良好控制的行为学分析(Scott & Panksepp, 2003)之后,我们努力在半小时的学前游戏课程中非正式地评估这个命题。当亲社会期望被温和但坚定地传达,而奖励是立即继续游戏时,幼儿理解并迅速内化了”己所不欲,勿施于人”的社会规则,以便继续享受乐趣(Scott, 2001)。这些考虑强调了PLAY在儿童社会发展中的重要性。
PLAY激活、心智发展和神经通路(尤其是社交通路)表观遗传变化之间的联系,肯定对给幼儿服用精神药物的做法有影响。例如,正如我们上面看到的,神经科学研究表明精神兴奋剂抑制PLAY系统:给予这些药物的动物游戏明显减少。如果PLAY激活是社会诱导的表观遗传和亲社会大脑逐步创建的重要促进因素,那么长期服用像哌醋甲酯(Ritalin)这样的精神兴奋剂可能对儿童人格发展产生有害影响。我们已经知道PLAY激活对新皮层产生强大影响。因此,服用精神兴奋剂也可能改变PLAY编程新皮层的方式。
服用精神兴奋剂显著减少儿童的游戏和多动症状这一事实,表明PLAY系统与注意力缺陷多动障碍(ADHD)之间可能存在联系。多动儿童的父母经常抱怨精神兴奋剂的一个不良副作用是减少了他们孩子的玩耍性。也许儿童的ADHD有时是游戏匮乏或特别强大的PLAY系统的指示,而不是精神病理学的迹象。尽管我们现在知道ADHD儿童在解剖学上(因此在功能上)其额叶执行功能有点缺陷(约5%)(Castellanos & Tannock, 2002),但这通常只有在孩子进入学校时才成为社会问题。他们不如那些具有更好的大脑-心智调节功能的儿童成熟和合作。
目前,治疗ADHD儿童的首选方法是哌醋甲酯(methylphenidate)和相关的精神兴奋剂,这些药物在化学上对大脑的作用与可卡因相似,只是效力和起效速度较低。长期以来,一个令人困惑的现象是,精神兴奋剂——这种促进运动性唤起的药物——往往能够平静问题儿童。现在,使用精神兴奋剂的一个新兴理论依据是,假设ADHD儿童在前额叶皮层结构和活动方面存在缺陷;因此,大脑的这一部分需要被刺激,以便促进注意力,从而更好地抑制过度的情绪性。这应该能增强儿童集中注意力和学习的能力,但对学习益处的证据相当薄弱。我们有争议的结论是,尽管少数ADHD儿童确实存在可证实的大脑问题,但大多数被诊断为ADHD的儿童并没有临床相关的大脑障碍(Panksepp, 2007b)。这些儿童中的许多人仅仅是在他们的玩耍冲动受挫时出现社交顺从性问题。
如果ADHD至少部分反映了对冲动性、玩耍活动的过度渴望(或饥渴),那么因为这些特质而给儿童服用药物是否符合伦理,就成为一个深刻的社会问题。显然,在课堂上保持对学业的注意力是必要的,但通过给儿童服用减少其玩耍性的精神活性药物来诱导顺从性是否恰当?至少,应该首先尝试更温和的干预措施,例如在上课前的清晨提供大量的打闹游戏活动,在乐于玩耍的年轻成年人的监督下,他们准备好促进社交学习,并在不好的事情发生时温和地进行干预。
我们过去使用ADHD动物模型的研究表明,长期每天充足的玩耍有助于减少幼年大鼠成熟时的冲动行为(Panksepp et al., 2003)。早期玩耍还可以使成年动物不那么具有攻击性和防御性(Potegal & Einon; 1989; Einon & Potegal, 1991)。除了来自动物界的证据外,还有人注意到,人类的病理性攻击行为往往源于缺乏玩耍性的童年,尽管肯定还需要其他促成因素(Brown, 1998)。尽管在玩耍过程中学到的一些技能最终可能有助于成年后的支配行为,但目前没有明确的证据表明充足的打闹游戏与成年形式的攻击行为之间存在联系。非常清楚的是,PLAY回路在很大程度上独立于攻击回路,玩耍通常教会人和动物如何更好地相处。
因此,我们相信,如果在我们的教育系统中充分利用PLAY的力量,特别是在学前教育阶段,我们将能够减少过于频繁的ADHD诊断。鉴于像利他林这样的精神兴奋剂潜在的长期有害影响,我们建议,如果孩子们以半小时的积极玩耍开始一天,他们可能会更好地学会在课堂上控制自己并吸收学业材料。
有一点是确定的:在玩耍过程中,动物特别倾向于以灵活和创造性的方式行事。毫不奇怪,玩耍干预已在教育和治疗环境(即游戏治疗)中成功使用,以促进新信息的有效获取和行为矫正(例如,Power, 2000)。然而,由于玩耍是有趣的,它也可以作为期望的行为改变的奖励。如果额外的打闹游戏的可获得性取决于良好的学业表现,儿童愿意在多大程度上通过学业任务来约束自己?如果将体育玩耍的可获得性用于系统地奖励学业成就,那么对课堂纪律和教育进步的益处可能会得到增强。这些考虑意味着我们必须将这一古老的进化大脑功能视为一种潜在的理想活动,而不是将其视为一种破坏性力量,其能量需要在认真的教育事业完成后被压制或在操场上消散。
还有一个担忧是,儿童服用精神兴奋剂可能会诱导对可卡因或甲基苯丙胺等滥用药物的敏感性和渴望增加。这种潜在影响从未在儿童中测量过,但已在其他动物中进行了评估。临床前研究提供了关于精神兴奋剂暴露长期后果的良好对照数据。成年动物通常会对所有精神兴奋剂的周期性给药产生敏感化。简而言之,它们的神经系统对各种滥用药物变得长期超敏反应,这种增加的敏感性反映在药物渴望的增加(Berridge & Robinson, 1998)以及对追求所有类型享乐性奖励的热切程度增加。用通俗的话说,这种动机强度的增加反映了正常欲望的转变:可以说,从”我想要它”到”我要它,而且我现在就要”。精神兴奋剂敏感化使动物更迫切地”消费主义”,更渴望所有外部奖励,从味觉享受到性(Nocjar & Panksepp, 2002)。相比之下,如果说在ADHD儿童的大脑中我们应该希望敏感化什么,那就是对亲社会活动的冲动。
尽管目前还没有研究尝试评估接受药物治疗的儿童与未接受药物治疗的儿童之间对药物渴望的强化程度,但早该评估精神兴奋剂诱导的”敏化(sensitization)“是否已经在因ADHD接受药物治疗的儿童中发生了。这可以通过对比儿童首次服药后与过去长期服药后精神兴奋剂的急性生理效应来实现。如果事实证明这些药物确实会产生持久的变化,我们应该担心这些效应对儿童并无益处。尽管我们知道非常年幼的动物不像年长动物那样容易敏化(Solanto, 2000),但我们确实知道它们会表现出一些敏化现象(Laviola et al., 1999; Panksepp, Burgdorf et al., 2002)。众所周知,这种大脑变化会促进寻求药物倾向的增强。
同样值得考虑的是,图雷特综合征(Tourette’s syndrome)及其奇特的神经冲动——导致抽搐和突然的言语爆发,通常包括”禁忌”表达如咒骂和诽谤(Chase & Friedhoff, 1982; Comings et al., 1991)——可能代表着异常的游戏冲动,或游戏冲动的组成部分,在神经系统中不受约束地循环。药理学证据为这一假设提供了一些支持。多巴胺阻断剂目前在控制图雷特综合征症状方面最为有效,同时在减少动物的游戏性方面也非常有效(Beatty et al., 1982, 1984; Panksepp, Normansell et al., 1987)。尽管这些联系具有高度推测性,但如果我们对这些可能性保持开放的态度,我们可能会更好地理解游戏的本质以及一些令人困惑的儿童期障碍。
当儿童一起玩耍时,他们会建立友谊,这使他们在一定程度上获得对父母的情感独立。游戏(PLAY)帮助培养感到快乐和自主的能力。这使儿童感到成熟、自立和有能力。这对他们的自尊和对他人的友好感是无法估量的恩惠。即使是宠物动物,特别是忠诚和爱玩的狗,在人类陪伴稀缺时也能帮助发展这种潜力。因为游戏在巩固友谊方面如此重要,它是允许儿童成熟的核心要素。经过精心磨练的游戏本能,被提炼得对他人的情感需求和愿望敏感,使儿童能够在家庭领域之外有效地发挥作用。这对某些人来说可能显得矛盾。毕竟,与工作相比,游戏有时被视为微不足道的追求。尽管如此,这项活动有助于培养满足和自我实现的成年人,因为它促进情感成长和社会敏感性。游戏有助于预防抑郁障碍,并促进大脑海马体(hippocampus)等区域的神经生长,这些区域在抑郁症患者中常常显示出压力诱导损伤的迹象(Wöhr et al., 2009)。
当儿童在游戏能力方面出现缺陷时,他们常常表现出抑郁并嫉妒其他儿童(Power, 2000; Powers et al., 2009; Ross et al., 2010)。这并不奇怪。如果他们难以参与游戏,看到其他人一起玩得开心就会产生怨恨。无论如何,儿童,即使是没有朋友的儿童,通常也会找到一种游戏的方式。有些人会发明想象中的朋友。毫无疑问,这些幻想(有时是妄想的)伙伴也是为了减少悲伤(GRIEF)的感觉而被创造出来的,但知道游戏系统的存在,以及所有儿童都有游戏的冲动,使我们能够理解为什么孤独的儿童会尽力寻找或发明快乐的伙伴。
如果得到良好培养,与其他儿童游戏的冲动自然会导致社会能力和对核心家庭的情感独立,这为青春期的成功成熟铺平了道路。它也可能为心智巩固一个情感上积极的基础,甚至深入到神经化学层面(Burgdorf et al., 2010),促进儿童在整个成年期顺利过渡到实现令人满意的亲社会目标。如果幼儿没有固定的玩伴,父母明智的做法是确保在孩子的日常社交饮食中有一些打闹游戏活动。显然,宠物往往可以作为儿童有益的游戏伙伴,这再次突出了这是一个可以在不同哺乳动物之间共享的心智-大脑(MindBrain)过程。
直到最近,神经科学家和心理治疗师往往忽视了所有年幼哺乳动物,包括我们的儿童,都有游戏的基本冲动——参与快乐竞争性互动的可能性。也许游戏被视为”幼稚”因而不重要。相反,严格的科学方法表明,所有哺乳动物共有的一个基本大脑系统解释了这种普遍倾向。目前的研究表明,游戏系统在新皮层的表观遗传发育和成熟中可能特别重要。进一步理解这个系统可能是解决某些儿童期情绪问题的关键。幼儿教育的目标应该是让孩子”五岁前茁壮成长”(这是华盛顿州目前儿童发展的口号)。要实现这一目标,游戏性必须成为整体方程式的一部分。对每个儿童游戏需求的普遍认识可能有助于在未来塑造明智的社会和教育政策。
总的来说,缺乏游戏的儿童不仅被诊断为ADHD的概率可能高于正常水平,而且成年后成为孤僻者和社会潜在威胁的可能性也更高。当然,人类人格的发展是一个多因素过程,成年后的生活状况很少能归因于单一原因。不良的养育通常伴随着许多其他参与因素,从营养不良到家庭环境中的攻击行为。然而,缺乏安全的婴儿依恋和缺乏早期游戏,无疑是促进成年后易怒和攻击性的促成因素(Brown, 1998)。
对社会而言,帮助创造条件让所有儿童在整个童年期间能够真正地游戏可能是明智的。现代社会中越来越多的儿童在能够充分进行身体游戏方面遇到的困难,目前可能正在以尚未测量的方式影响文化品质。我们认为,许多社会已经远离了我们人科动物过去的社会生态需求,为了防止社会人际品质的下降,也许我们需要建立更多的”游戏庇护所”——让儿童能够安全地沉浸于他们自己发起的游戏活动中的场所。
这既不是新颖的想法。早在科学家意识到大脑功能及其基因组成之前,柏拉图就在他的论著《法律篇》[VII, 794]中赞扬了自由童年游戏的益处:
到三岁阶段,以及四岁、五岁、六岁之后,游戏将是必需的。这些是大自然在那个年龄自然提示的游戏;当儿童被留在彼此的陪伴下时,他们很容易自己发明这些游戏。所有三到六岁特定年龄的儿童,应该首先在当地的庇护所集合——每个村庄的所有儿童都在同一个地方集会。此外,保育员要关注他们行为的得体或不得体。[强调为后加]
柏拉图的基本信息是,我们的儿童没有游戏就无法完全成为人类。今天也没有什么不同。但今天我们确实对游戏在促进儿童福祉和心理健康方面的作用有非常积极的讨论(例如,Schaefer & Kaduson, 2006)。
丰富的早期游戏机会可能在文化和表观遗传学上有益于儿童快乐和共情的脑心智发展,并持续终生。它也可能有助于缓解已经成为我们常规商业经济环境特征的自私贪婪。社交游戏可能有助于打开更好理解他人的门户,从而产生亲社会倾向,从增强的社交性到直接的共情。但这些只是想法,像许多其他想法一样,仍有待神经科学家和临床医生以严格的方式进行实证评估。在第12章中,Panksepp将考虑游戏性能量如何通过心理治疗互动中新的情感平衡方法促进快速的治疗改变。
再次深入那古老的知识源泉
深入我们超过40亿年的创造深处
深入那将地球聚合在一起的创世烈焰
在古人的混沌和开端中
在静谧中追溯造就我们今日之貌的生命不断之线
思考所有那些逝去的生物
我们佩戴着每一个曾经漂浮、蠕动、游泳或奔跑者的面容
我们行走在它们开辟的古老道路上
——Sandy Hartman,《再次深入内心(精神的生物学)》(2011)
本章提供了一个关于如何处理神经科学中最棘手问题的愿景。正如Robert Holt在1989年所说的,“主观体验存在于物理化学世界这一事实是一个难以穿透的谜团。”对我们来说,这个问题归结为神经系统究竟如何产生主观情感体验的问题。我们已经提供了证据,表明初级过程情感在很大程度上来自产生连贯情绪行为的相同皮层下回路。换句话说,无论在何处通过局部皮层下脑刺激引发连贯的情绪行为,这些脑心智状态都作为学习控制中的”奖励”和”惩罚”。要进一步理解情感体验的深层神经本质——现象意识的基本形式——可能需要对一些人称之为”核心意识”的基本大脑过程进行实证阐明。也许体验的原始本质如果没有对拥有”核心自我”意味着什么的现实”具身”愿景就无法得到澄清——甚至可以带点诗意地将其称为我们的动物性”灵魂”。我们再次将这个大脑功能的术语大写,因为它被认为是心智的一个初级过程——一个”简单自我型生命形式(SELF)“——一个内部有机体内脏-情感和外部感觉-运动表征的连贯重心。
人类”灵魂”的问题——反映了我们每个人都是独特”我”的这种难以言喻的感觉——有着曲折的历史。它导致了形而上学的二元论——心智与身体的分离——以及现代对”自我”本质的无穷变化的思考(见Gallagher & Shear, 1999; Panksepp & Northoff, 2009)。正如普及二元论的勒内·笛卡尔(René Descartes)(1596-1650)所说:“我知道我存在;问题是,’我’所知道的这个’我’是什么?” 他的思考最终引出了他个人的”解答”:“我思故我在。” 笛卡尔和许多其他哲学家都认识到我们个人记忆的重要性,这些记忆是我们思想的基础,也是我们人类独特的自我概念的基础。这句格言催生了大量变体。我们更喜欢”我感故我在”(Panksepp, 1998a, pp. 308, 420)。这突出了我们基础情感体验的连贯性。
这些理解核心自我的替代观点试图认识到,原始的情感体验形式(初级过程或”核心”意识)——以基本感受(非反思性、非理性、情感性的存在于世界的方式)为特征——肯定在地球上出现得比认知觉知等更高形式的意识要早。这在哲学中并非主流观点,但它与一些人的观点一致,例如大卫·休谟(David Hume)(1711-1776),他在《人类理解研究》中将其哲学建立在我们拥有情感体验的能力之上。但休谟也认为,我们的记忆就像串在线上的珍珠,将个体生命汇聚成一个连贯的”我性”——自我。
伊曼努尔·康德(Immanuel Kant)(1724-1804)欣赏休谟论证的说服力,但在他的《纯粹理性批判》中,康德继续发展他的”理性主义”论点,即心智拥有先验的认知能力,这些能力提供先于经验的内在知识。许多其他理性主义-认知主义哲学家发现很难将他们对更高概念性心智的愿景与可能由情感核心-自我构成的存在基础联系起来。虽然这可能不是一个”非此即彼”的问题,但我们将在本章中探讨这样一种可能性:一个具身的核心-自我过程——身体(尤其是内脏身体)在大脑中的原初表征——可能是情感”存在”和更高心智装置出现的基础。核心自我在这里被设想为那些产生有机体连贯性的深层皮层下过程的标签——一个具有多样化情感感受的活跃有机体的统一存在。(关于这些观点如何能够为无教派宗教体验提供普遍基础的深刻历史-哲学分析,见Thandeka, 2005, 2009。)
本章的目标是探讨这样一种可能性:对”具身自我”的神经科学理解——一个植根于身体及其神经表征的自我——可能会澄清体验如何在心脑进化中首次出现的困境。这个困境尚无公认的解决方案,也没有明确的答案。但从自下而上的神经进化观点来看,可实验检验的想法正从理解初级过程情绪性的情感神经科学策略中产生。
原始情感体验是如何在人类大脑中产生的?我们对这个问题没有最终的实证答案,但本书已经论证了这样一个观点:我们现在知道该往哪里看——我们应该在哺乳动物大脑古老的内侧皮层下网络的祖传、产生情感的本能机制中寻找。如果是这样,上述问题的详细答案只能通过因果性脑研究来获得,而这项工作在人类身上显然在伦理上是不可能进行的。许多人会说这也适用于其他动物。无论如何,除了研究其他动物的相应过程外,我们没有有效的策略。要做到这一点,动物显然必须具有初级过程的情感体验。否则,就这一主题对动物大脑的研究从定义上讲就是愚蠢的行为;基于大量证据,我们可以确信事实并非如此。
本书概述了其他动物具有古老情感感受的证据。为什么这些证据的分量仍未被大多数神经科学家接受,这是一个文化-历史问题,而不是科学问题。一个主要的历史原因是,该领域的领导者长期以来一直告诫学生要认识到,既然”主观现象在动物身上无法被客观观察到,那么声称或否认它们的存在都是徒劳的”(Tinbergen, 1951, p. 5)。事实上,在动物神经科学领域并不罕见的不可知论和唯我论世界观导致这样的结论:其他动物甚至人类的行为永远无法提供确凿证据证明它们体验到了什么。
然而,对这个问题的科学方法是一个可观察预测和由此产生的证据趋同的问题,而不仅仅是争论。现在有压倒性的证据表明,当所有哺乳动物情绪脑的古老网络被直接操纵时,它们都会有强烈的体验。因此,产生连贯情绪反应的大脑网络也会产生感受。但这仍然留下一个重大的科学问题,神经科学家才刚刚开始讨论。这种从大脑活动到心理体验的”魔法”转变实际上是如何发生的?大脑的物质过程如何产生心智,一个”我”?没有人知道答案。目前,只能有假设……希望是可检验的假设。本章的目标是提出这样一种可能性:核心自我(core-SELF)概念对于取得实证进展将是非常宝贵的,尽管在很长一段时间内,怀疑肯定会超过确定性。尽管如此,大量证据表明,情感性感受(affective feelings)在大脑进化中非常古老,但我们现在必须接受关于它们如何实际从神经活动中构建出来的新想法。我们可以相当有信心地认为它们源于脑干内侧区域(Panksepp, 1998a, 1998b),但无论是精确的神经机制还是产生明确理解的策略都不是很清楚。然而,核心自我似乎明显与脑心(BrainMind)的初级过程情绪和其他情感过程有关。我们的主要希望是,通过将一些可检验的想法摆在桌面上,我们将激励年轻学者开展必要的研究,以实证方式澄清潜在的可能性。
我们使用大写字母的术语”核心自我(core SELF)“,原因与我们用大写字母来指定七种基本情绪系统相同。这些大脑系统在哺乳动物物种之间是同源的(homologous),可能在其他脊椎动物之间也是如此。通过为多样化的情感体验提供一个共享的神经平台,核心自我可以被视为一种”通则性的(nomothetic)“(普遍的)大脑功能。当核心自我以及它所精心制作的许多原始感受与高级认知三级过程相互作用时,它促进了各种”个异性的(idiographic)“(个体独特的、经验精炼的)”扩展”自我的出现,这发生在发育性大脑成熟过程中(Northoff & Panksepp, 2008; Panksepp & Northoff, 2009)。
当然,核心自我结构在不同哺乳动物物种之间的细节上有所不同。不同物种身体的显著差异肯定会反映在脊椎动物物种之间这些心理存在基础网络的自然变化中。然而,由于大脑是一个进化上分层的器官,最古老的生存功能被最深刻地保存下来,我们只是认为存在显著的进化相似性(同源性),超过了差异。所有哺乳动物的情感进化”生存工具”都非常相似。与普遍的(通则性的)核心自我相反,依赖经验的个异性自我不是同源的。这是因为新皮层的生长和由此产生的认知能力在不同物种之间差异巨大,导致反思性意识的巨大差异。(我们使用小写术语来讨论这类高阶过程。)个异性自我在每个生命周期中通过每个人和每个动物独特的经验景观而出现(Panksepp & Northoff, 2009)。
在本章中,我们分享一个关于自我初级过程本质的工作假设,基于对神经科学证据的跨物种解读。关于皮层下大脑区域初级过程情感产生的趋同证据鼓励我们谈论一个非反思性的(无意识的,anoetic)自我和纯粹的情感意识形式。在这种观点中,核心自我和各种初级过程情绪系统提供的各种先天生存工具是同时产生有机体情绪-行为连贯性以及相关情感状态的必要成分。这种观点还提出,核心自我和七种情绪系统与高级大脑功能(如工作记忆)相互作用,允许更高层次的反思性”认知”(意识性,noetic意识)以及多层次存在性自我意识的出现,这是人类心智的一种发育性的、也许是独特的品质。在感知到的世界事件中体验自己作为一个特定和个体的主动行为者的难以言喻的感觉,肯定反映了脑心最近出现的能力,构成了一种认知的、甚至是理性的意识形式。
正如前面已经提到的,假设皮层下结构产生了意识的原始形式,这并没有什么不寻常或不现实的地方。事实上,在上个世纪中期,神经科学家发现网状结构(reticular formation)——一种位于核心脑干中松散结合的细胞体和神经纤维的聚集体——允许大脑皮层产生觉醒状态(Watt & Pincus, 2004)。新皮层本身无法维持意识。因此,我们过去记忆的储存库——可以被调入主动计划模式或”工作记忆”功能——集中在大脑的背外侧额叶区域。我们对所有这些如何与个人关注点相关的评估集中在内侧额叶区域(Northoff et al., 2006),而这种思考深受皮层下初级过程情绪功能的影响(Panksepp & Northoff, 2009)。可能还存在中间脑区,如眶额皮层,专门负责情感工作记忆。
从心理学角度看,核心自我主要由情感性感受所支配,并伴随着对世界和身体内部稳态的一些初步知觉。更高形式的自我意识是通过这些初级情感能力与编码动物生态、社会和文化环境的次级/三级心智能力的相互交织而形成的。下一章将关注这些皮层下情感系统的功能与它们对心理健康或疾病(以及相关心理治疗问题)影响之间的一些可能联系。
我们在这里考虑的问题如此复杂且难以理解,因此我们可能需要从稍微不同的概念轨迹再次考虑一些之前涉及的问题。当然,从哲学角度来看,如果不假设自我作为情境化体验的实体,就很难思考意识。从神经生理学角度来看,我们必须设想有机体的基本连贯性(coherence)——它们在世界中内在感受到的统一存在——是如何由大脑古老的皮层下中线系统创造的。我们有什么进化理由来论证这些古老神经复杂性中存在一个自我?这就是我们现在要探讨的问题。
如果我们仅仅考虑生物与非生物有机体之间的根本差异,从进化角度娱乐核心自我的概念就很有意义。生物执行新陈代谢功能,使它们作为离散实体保持活力。例如,它们利用能量供应并排除废物。非生物不执行新陈代谢功能。非生物由于其化学组成而作为单元保持连贯性。它们没有明确和独特的生化过程来主动维持自己作为个体实体。因此,在身体层面,新陈代谢(metabolism)区分了生物和非生物。在大脑层面,内源性产生的运动性(motility)(自发的身体活动)以及在高等动物中预测未来事件的能力区分了生物和非生物。新陈代谢和运动性的基本能力,虽然最初是无意识的,但为意识的出现和进化提供了必要的基础。
我们与其他神经科学同事(Damasio, 2010)一起设想,在大脑进化早期,出现了身体的原始神经映射(map),以促进许多不同功能的整体连贯性,从行动倾向到伴随行动的自主神经变化(Northoff & Panksepp, 2008; Panksepp, 1998b)。我们与达马西奥(Damasio, 1999)一起,将这种身体映射称为原始的”原自我”(proto-self),它随着初级过程情绪和动机系统的出现而进化成为一个更复杂的心智器官——核心自我。我们假设这种类型的大脑组织整合了原始体验,如原始感觉、稳态和情绪性情感。然而,这还不是”觉知”一词通常所指的含义。
为了理解核心意识,我们还需要设想更高层次的心智是如何从更基本的形式进化而来的。因此,我们必须区分原始的现象性意识形式——它提供了纯粹体验的能力,但尚未具有反思体验的能力(即具有自我意识觉知(self-conscious awareness),定义为能够将自己设想为世界舞台上的体验行动者的能力)。这是相当复杂的东西,远远超出了仅仅在世界中体验自己的能力。
我们假设原始现象意识有两种形式。首先是体验各种积极和消极情感的能力——各种形式的”好”和”坏”——特别是本书关注的原始情感体验。其次是以体验方式感知世界的能力(即”头脑中的电影”),这是认知意识的基础。我们很容易说不知道哪种在大脑进化中更古老,或者这些类型的现象体验是如何耦合的。但如果必须做出选择,我们会认为情感形式的主观体验在心脑进化中比认知形式更古老,因为它们在大脑更内侧和尾侧(朝向尾部)的区域得到发展,因此也是更古老的区域。一个突出的例子是导水管周围灰质(PAG,也称为”中央灰质”)的位置,它位于中脑的核心。相比之下,离散的感觉-知觉功能位于更外侧,这表明它们起源较晚。
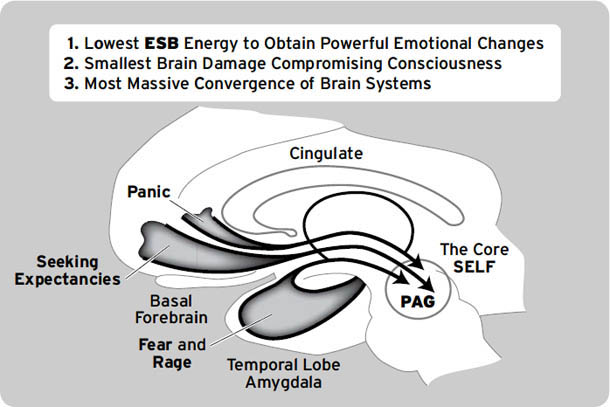
图11.1. 前脑区域概览,这些区域致力于阐述基本情绪过程的高级表现。每个情绪系统都有更高的影响范围,恐惧和愤怒集中在外侧和内侧颞叶,寻求集中在腹内侧额叶,各种社会情绪过程如分离痛苦或恐慌/悲伤集中在前扣带回。所有这些系统都汇聚到中脑的情绪和自我表征区域。图中突出显示了PAG的三个特性,这表明它对大脑核心情绪处理的关键重要性,与高级脑区相比(摘自Panksepp, 1998a;经牛津大学出版社许可重新发表)。
在早期脑心进化过程中,原始情感和感觉现象体验最初是紧密联系的,还是从一开始就是大脑两种根本不同的原始意识形式?我们不知道。但可以从这样的假设中构建新的理论视角:有意识的视觉和听觉的体验最初在很大程度上是情感性的(Panksepp, 1998b)。突然的视觉或听觉刺激能让我们立即受到惊吓和恐惧,特别是当这些刺激源非常接近我们的身体时,这表明这些感觉系统与我们一些最基本的情感生存机制之间存在深层的原始整合。还要考虑我们倾向于将特定颜色与感觉联系起来——红色与激情唤起,黄色与快乐,蓝色与凉爽或放松状态,绿色和棕色与对生命土地的安全热爱,黑色与死亡。同样,想想声音如何轻易唤起我们的情感,从某人的语调、大自然中鸟儿的歌声,到人类创造的音乐奇迹(Malloch & Trevarthen, 2009; Panksepp & Trevarthen, 2009)。音乐的动态似乎能直接触及我们核心意识的情感结构(Blood & Zatorre, 2001)。
在这种背景下,应该强调的是,集中在中线结构中的古老情感过程如果不与感觉过程相联系就不会有用,而它们确实是相联系的。然而,这些感觉输入不是那些上升到丘脑然后进入新皮层的输入。它们是直接进入自我相关信息处理的中线网状区域的古老支流,与情绪-情感处理直接相关。再次强调,这并不意味着通往大脑更高区域的较新信息流不能影响情绪。它们当然可以,但这主要需要学习。情绪的高级认知调节不是大脑的内在精细功能;它主要通过情绪教育和由此产生的社会智能而出现(Goleman, 2006; Keltner, 2009)。这使得情感敏感的心理治疗成为”成长”的有用帮助(见第12章)。但也存在自上而下、高级脑调节情感强度的进化倾向。新皮层可以内在地抑制初级过程情绪,倾向于将其保持在”潜意识”领域,直到需要处理重大生活挑战情境。然而,在精神病理学中,这种调节控制常常失效,个体被他们无法再控制的情感淹没。许多心理治疗包括促进对自己情感的认知调节。
无论如何,非常清楚的是,情绪情感的初级加工机制位于大脑更深层的中线区域,这表明情绪行为和伴随的情感在脑心智进化中具有某种优先级——它们在精密的感觉-知觉新皮层能力(如我们清晰而精细的视觉和听觉能力)之前就已进化出来。然而,皮层下结构能够以复杂的方式处理感觉信息,因此在新皮层之下也存在一些知觉-现象体验(见第6章)。例如,上丘(superior colliculus)能够在某种心理分辨水平上感知视觉信息,下丘(inferior colliculus)能够感知听觉信息(Merker, 2007)。类似的皮层下能力也存在于对触觉的感受中。这些皮层下结构不会产生清晰的看、听和体表刺激的意识体验,但它们确实以独特的方式给予事物的感知。例如,这些皮层下系统能够确定视觉和听觉刺激在空间中的位置,但在儿童早期的体验水平上比成年期更为有效。
显然,我们体验为五大感官的皮层能力,通常具有显著的知觉清晰度,其进化远晚于皮层下的知觉敏感性。因此,随着个体成熟,高级脑功能的”接管”如此显著,以至于大多数研究者认为皮层下系统一直都是无意识的。(这解释了经典的神经学现象,如”盲视”(blindsight)——即无意识地看见——它几乎是无意识的,但并非完全如此,因为人们能够正确定位空间中移动的物体,而没有任何清晰的视觉感知;他们只是模糊地感觉到某些东西,如纯粹的运动,正在空间的特定位置发生。)同样可能的是,皮层下知觉功能在发展上变得潜意识,因为更高级的体验机制逐渐在行为控制和对意识注意资源的需求中占据主导地位(全面讨论见Merker, 2007)。
特别值得注意的是,皮层下知觉加工继续与核心自我(SELF)的情感网络相互作用。这有助于解释我们在第5章和第6章中关注的情绪唤醒的”低级通路”。此外,值得考虑的是,在心智-脑进化中,生物体在拥有能够代表重要生存需求的神经系统——即初级加工情感——之前,可能不需要精细的知觉。从这个角度来看,我们可以理解,在为生存而奋斗和竞争的无数代中,拥有原始情感系统的生物如果能被日益精密的知觉和认知-学习机制更准确地引导,就会具有巨大优势。在心智脑进化的早期阶段,情感核心-自我网络与外部世界之间相当简单的感觉连接可能就足够了(例如,恐惧条件反射的”低级通路”)。然而,越来越精密的距离感受器(如皮层听觉和视觉)的效用,以及利用这些信息进行策略思考的能力,在后期的脑心智进化中获得了丰厚回报。所有这些让我们认识到,为什么某种代表原始身体的神经符号矩阵,反映一个自发活跃、情感反应性的有机体,在脑进化中比精密距离感受器及其新皮层分析器的出现更早确立。这也让我们理解为什么需要区分初级情感机制和高级认知机制。
直到今天,最古老的感觉系统如嗅觉、味觉和触觉,对人类来说仍然比视觉或听觉具有更直接的情感性。也许原始的知觉敏锐度最初与这些最优服务于情感性身体和大脑需求的信息通道紧密相连。内稳态情感与代表身体状态的情感过程仍然特别紧密地交织在一起。因此,饥饿感仍然刺激食欲,也放大了味觉和嗅觉的愉悦。我们仍然赋予颜色知觉以情感特征——从热情激昂的红色和欢快的黄色,到清凉放松的蓝色,以及舒缓的棕色和绿色。因此,从进化角度来看,我们明智的做法是保持开放的可能性,即情感可能引导了许多感觉-知觉能力的构建(Panksepp, 1998b)。如果这一情景是正确的,我们将永远无法完全理解更高形式的意识,除非首先破译更原始的情感形式。
这可能再次有助于强调为什么我们将这个知之甚少、几乎未被研究的情感自我表征的神经基础称为核心自我(core SELF)。它照顾即时的身体关切——产生(i)寻求(SEEKING),首先照顾内稳态需求,如水、能量和热平衡(Denton, 2006),然后是更微妙的情绪需求;(ii)愤怒(RAGE)和恐惧(FEAR)以避免身体毁灭并有效竞争许多对(iii)原始欲望(LUST)至关重要的资源,后者促进物种生存。这些爬行动物情绪逐渐被更微妙的社会原则所补充。心智进化的下一阶段,大概在鸟类和哺乳动物分化之前存在的物种中,增加了独特的社会-情感系统:关爱(CARE)、悲伤(GRIEF)和玩耍(PLAY),所有这些都建立在先前存在的爬行动物情绪之上,尤其是寻求(SEEKING)。
让我们用另一种(稍有不同的)方式来表述这一切:核心自我(core SELF)在神经科学界既缺乏充分理解,也鲜少被讨论。然而,我们将其视为创造所有情感体验的神经基础。它具备恰当的要素,尤其是一个庞大的神经符号表征场域,涵盖从心脏到肠道的各种身体器官,这些都是情感体验的组成部分。如果情感不被主观的”我”所体验,那它又是什么呢?它首先将各种稳态状态表征为心智大脑(MindBrain)的情感状态,通过大脑内身体的神经表征所发生的变化来体验(这些神经网络当然仍与外周身体中实际发生的情况相互关联)。例如,低体液水平和高血液溶质水平被体验为口渴,血糖的快速下降唤起饥饿感,等等。最重要的感觉体验,如食物的气味和各种类型的触觉,都被体验为情感上的愉快或不愉快。这些相关联的情感往往可能是习得的,这凸显了我们如何发展出习得的偏好和厌恶。
随着复杂情绪网络的出现,核心自我还能够预期各种环境变化。例如,捕食者的气味会唤起恐惧系统(FEAR system),核心自我会转变为一种独特的神经动力学状态,这种状态仍被体验为恐惧感。同样,母亲乳房的气味,尤其是当饥饿信号强烈时,会唤起婴儿的寻求冲动(SEEKING urges),他们会接近并更靠近母亲的身体进食,并同时通过母性关爱(maternal CARE)获得情感滋养。从进化的角度来看,值得注意的是,稳态的身体需求系统集中在下丘脑的中线位置,处于比稍微更外侧的寻求系统更古老的进化位置,而寻求系统,正如我们反复强调的,是所有情绪系统中最大、最普遍的。性欲系统(LUST)和关爱系统(CARE)位于身体需求系统的更前方位置,这再次清楚地表明了稍晚的进化起源。
这是一个重要观点。所有情绪系统在大脑心智进化中的地位并不相同。可以假设,在原始的”爬行动物”情绪中,通用的寻求系统在早期就被设计用于获取大量资源,包括躲避危险的安全(例如逃跑)。因此,寻求系统可能作为平台(预适应),促进了我们讨论过的其他社会情绪系统——性欲、关爱、玩耍(PLAY)——的进化出现,同时也促进了古老的恐惧和愤怒系统(RAGE)。
这是寻求系统首先在这里被讨论的一个原因。它也是最有证据介导一种非常特殊的积极情感的系统——不是感觉的愉悦,而是积极的振奋、欣快的兴奋,源于与世界的富有成效的互动。幼儿看似无限的热情无疑部分源于这个系统,鼓励玩耍。正如第三章所强调的,寻求冲动不仅仅是”大脑的奖励系统”,而是一个追求有机体幸福的系统。寻求系统的唤起无疑是有奖励性的,但这些奖励不是典型的感觉愉悦,而是追求各种奖励的渴望,这一观点也以更侧重感觉的方式被强调(“激励显著性”(incentive-salience)——一种条件性次级过程;见Berridge等人,2009)。
核心自我建立了一个明确的框架来解释情感意识的存在,并摆脱”读出”机制的无限回归(见第二章)。原始的基本情感要被体验,不需要被更高级的心智大脑机制”读出”,尽管许多当代情绪理论坚持这样认为。当然,更高级的皮层功能可能会增添其他类型的感受,尤其是通过允许原始感受渗透并与认知交织——更高的大脑功能可能”倾听”较低层次的功能,并为体验增添额外的认知解析的情感色彩。通过这种方式,次级和三级心理情感过程可能会创造出各种更微妙的高阶感受——如勇气、嫉妒、内疚、妒忌、骄傲、羞耻和社会厌恶/轻蔑,仅举几例(关于妒忌的完整讨论,见Hart & Legerstee, 2010)。核心自我的连贯性可能使人类和动物拥有一种拥有其情感体验的基本感觉:这些情感是他们心理上不可分割的一部分。
从历史的角度来看,重要的是要注意,这些是行为主义者通过教条式地回避大脑活动的心理维度而决定称之为”强化”行为改变的”奖励”和”惩罚”的大脑-心智基质。由于这些概念选择的结果,奖励和惩罚可以被定义为动物皮肤之外世界中的可观察事件。因为科学家可以使用关注世界中物体的操作性定义,而不是内部过程,所以关于动物情感体验的讨论被视为没有意义的。所有必要的概念(除了”强化”,它在系统中构成某种”粘合剂”)都方便地用视觉上明显的环境和学习术语来定义,而不需要沉溺于模糊的神经心理学概念。“奖励”和”惩罚”只是世界中的事件,仅此而已。然而,神经科学很快开启了这样一种可能性,即它们实际上是神经功能或大脑的体验过程。但这些想法受到了阻碍,甚至受到了积极的压制。当科学对话停止时,教条而不是知识就开始统治时代。我们对心智-大脑过程真实本质的理解相应地减少了。
目前的证据表明,在最原始的层面上,原始的情感体验emanate(源自)皮层下中线系统(SCMS),该系统位于上脑干(中脑、下丘脑和丘脑),并与更多头端的内侧扣带回、岛叶和额叶以及眶额皮层区域密切连接。我们在独特的个体化自我中情境化普遍的、初级过程情感的能力需要更高的皮层中线结构(CMS),以及许多其他更高级的大脑组织(Northoff et al., 2006; Northoff & Panksepp, 2008; Panksepp & Northoff, 2009)。然而,这种体验的个体化情境化如果没有从这个神经连续体的下层出现的有机体一致性是不可能发生的,SCMS在内脏身体表征方面特别丰富,这里被设想为核心自我的中心。
尽管我们还不能确定核心自我的精确解剖学组成部分,但任何可行的大脑候选区域都应该满足以下标准:(i)基础设施应该是古老的,位于大脑的祖先中线区域;(ii)关键系统应该是多模态的,能够在神经轴的许多层面上被表征;(iii)回路应该具有特征性的、先天的静息状态,表明与稳态偏离的某种设定点;(iv)自我的共享基础设施应该在初级过程情感状态期间以不同的方式被唤醒。总之,自我结构应该与大脑的其他部分非常密切地连接。
SCMS/CMS连续体的初级过程情绪网络充分满足这些标准。这些古老的深层中线系统”评估”身体和世界的状态,并对典型的生命挑战事件产生情绪反应。SCMS的原始感受随后在CMS内被重新表征,最终在相关的情感记忆领域内被表征为自我的个体化呈现。许多研究人员现在已经观察到这些中线系统中特征性的静息神经活动,当人们在脑扫描期间什么都不做(自我反思和/或沉思)时,这些活动比他们受到各种认知任务轰炸时更强烈(Damoiseaux, et al., 2006; Fox and Raichle, 2007; Fransson, 2006; Raichle et al., 2001; Rilling et al., 2007)。这些系统在抑郁并沉思自己生活状况的个体中也异常活跃(Alcaro et al., 2010; Grimm et al., 2008, 2009; Northoff et al., 2010)。至少有六个这样的皮层下中线结构与大脑的其他部分特别密切地连接。这些包括,从下往上,(i)小脑深核和第四脑室周围底部(臂旁区和迷走神经背运动核)的大部分;(ii)PAG和紧邻的中脑区域;(iii)上丘和下丘,特别是更深层的运动成分;(iv)腹侧被盖区(VTA);(v)下丘脑;以及(vi)一系列基底神经节核,最突出的是杏仁核和伏隔核。这些部分中哪一个对核心意识最重要?
整个小脑的移除不会严重损害意识。因此,我们显然排除它作为核心自我的基质,尽管它肯定调节和控制情绪行为。大面积小脑损伤确实会显著损害各种肌肉协调,而这当然是所有行动所需要的,尤其是复杂的情绪反应。事实上,这可能就是为什么深层小脑系统在人类大脑情绪唤起成像中经常”亮起”的原因:所有情绪行动都需要许多身体部位的复杂协调。第四脑室底部的损伤很可能会导致动物死亡,因为许多自主神经功能受到损害。在顶部,所有各种基底神经节核团都可以被破坏,产生许多行为缺陷,但不会损害核心意识;它们只是损害基本学习机制,如经典条件反射。这意味着上丘的深层运动层和导水管周围灰质(PAG),以及相关的腹侧被盖区(VTA)和下丘脑回路,可能是支持核心自我的最重要结构。我们已经注意到,PAG是大脑中最古老、最高度集中的情绪汇聚区;这就是我们在此重点关注它的原因(有关出色总结,见Watt, 2000)。PAG对于产生原始情绪感受比杏仁核重要得多,尽管杏仁核因为在各种情绪唤起(通常是认知-情感任务)期间如此经常地”亮起”,继续在大众媒体中被过度宣传为大脑最重要的情绪中枢。实际上,它是一个认知-情绪接口,而不是主要过程情感体验的产生器。在专门处理外部信息的心智认知方面与告知我们大脑和身体状态的情感心智功能之间,还有许多其他这样的接口核团。
PAG与其他脑区的关系也暗示了它在整体情绪生活中的某种核心作用。上丘的深层构成了身体的基本运动映射系统,当向中脑外侧移动时,它与一系列感觉系统(触觉、听觉和视觉)相互作用,当向中脑核心内侧移动时,也与PAG的许多情绪系统相互作用。与PAG相邻的是中脑运动区(MLR),它能够启动神经模式,这对于建立各种连贯的情绪行动倾向至关重要,例如朝向或远离世界事件奔跑。因为情绪表达由这样的情感行动倾向组成,MLR也可能是核心自我的一部分。VTA是一个可行的核心自我候选者,因为VTA和其他邻近区域的多巴胺能场,通过它们向内侧皮层下和皮层前脑区域的丰富投射,阐述了寻求冲动的本能和习得成分,这些冲动在大脑较高中线区域促进与自我相关的信息处理。
最后,尽管我们没有强调它,但维持皮层清醒活动的上行网状激活系统(ARAS)就位于PAG和MLR旁边。该系统主要由上行乙酰胆碱、组胺、去甲肾上腺素和血清素系统构成——所有这些都调节注意力和清醒唤起(Pfaff, 2006)。它与丘脑网状核相互作用,促进整个大脑较高知觉区域——控制视觉、听觉、触觉以及所有这些感官相互作用的新皮层区域——的感觉刺激处理,这些相互作用使我们能够用语言和思想重新象征世界。ARAS是历史上首次被认为参与清醒调节的大脑系统,因此它是意识控制,尤其是认知意识控制的主要参与者。目前尚不清楚这些系统如何参与特定形式的情绪唤起,但它们肯定在许多脑区的一般唤起中发挥作用,尤其是情绪状态期间的新皮层区域。例如,大脑去甲肾上腺素促进传入感觉信号处理,使信号被更强烈地感知(也就是说,它们在脑心智中具有更多的显著性)。该系统可能也增加情感显著性的强度,这可能主要被体验为一般情绪唤起。相比之下,PAG的更内侧区域似乎阐述更具体的情绪行为和相关的独特感受。
这些古老中线回路的复杂性正在被积极探索(Holstege & Saper, 2005)。通过监测脑干的特定运动核团以观察情绪的可观察行动,以及通过监测自主神经核团以观察情绪的内脏成分,将更容易理解PAG内整合系统下行”输出”的功能组织,包括进入ARAS的输入。重要的是,这些情绪输出受到来自PAG的连贯”管弦乐”控制,PAG是情绪唤起的中心,但对于核心自我的实例化也至关重要。
核心自我所体验到的意识从根本上是情感性的,不包含任何命题内容。我们认为,每个情绪系统都能在皮层下中线结构——即核心自我——内产生不同的全局神经动态,从而产生人类和其他哺乳动物所体验到的不同的原始过程情绪。试图用文字描述这种大脑机制是困难的,我们只能想象每种情绪的特征性动态。我们认为,在数天或数周内持续思考这些问题可能有助于理解。此外,以不同形式重读同一观点应该有助于整合(Austin, 1998; Panksepp, 1998b, 2009b)。
我们设想以下类型的过程。七种基本情绪系统的唤起可能导致每种情绪产生特征性的大规模神经放电模式;这些模式是构成原始过程情感的神经心理特征的特征性振荡,它们在动态上类似于同时释放的情绪行为(理解的一个困境,尤其是对最聪明的人来说,可能是人类可以通过高级皮层意志力抑制这些表达性的低级脑功能)。无论如何,各种原始过程情感动态可能反映了核心自我结构内神经放电的速率和模式,导致大脑古老的内脏-躯体身体地图内全局振荡的独特模式。证据表明,正是在这些神经振荡中,不同的情感唤起出现,产生神经系统的各种积极(“奖赏性”)和消极(“惩罚性”)状态。换句话说,情感的神经动力学(大规模神经元活动)类似于各种原始过程本能情绪行为,即愤怒(RAGE)的猛烈冲击、恐惧(FEAR)的僵硬紧张、关怀(CARE)的抚慰,等等。因此,核心自我中情感网络放电动态与本能情绪行为之间的生物物理相似性使我们能够通过研究动物客观的本能情绪表达来研究它们的主观情感感受。
这种大脑情绪唤起的独特全局动态尚未在大脑内部得到客观测量。事实上,脑科学家目前没有有效的方法来研究全局神经网络动态。这使得对人类情绪动态体验的精神分析研究尤为重要。然而,在传统神经科学文献中,对于大脑内某些类型的相关同步振荡有一些提示性线索,例如海马体中的4-7赫兹节律,称为θ节律(theta rhythm),它帮助动物探索世界(例如,大鼠的嗅探)并由此在海马体中创建记忆。θ节律是海马体在积极处理信息时高度特征性的神经特征。当人工唤起大鼠的探寻系统(SEEKING system)——一个主要的信息收集情绪系统时,当动物嗅探和探查周围环境时,这种节律尤为明显(Vertes & Kocsis, 1997)。换句话说,嗅探节律通常对应于海马θ节律的持续频率。重要的是要回忆(见第3章),这种嗅探节律仅通过在固定间隔时间表上电刺激探寻系统就会以预期的方式自发地条件化(例如,每20秒给予一次脉冲奖赏性脑刺激)。这可能突出了认知知识如何从情感过程的模式化唤起中产生,或许使我们能够理解为什么康德推断大脑具有内在知识。这些数据支持这样的观点:情绪系统帮助在大脑的高级区域创建知识。同样,正如我们在第5章和第6章中讨论的,恐惧的非条件情绪反应可能对大脑高级区域的简单恐惧条件化的产生至关重要。
因此,每种情感的特质可能源于每个情绪系统的特征振荡在核心自我的全局神经空间内促进动态变化的方式,所有原始过程情绪网络都嵌入其中。例如,恐惧系统的唤起可能产生快速振荡,将核心自我推入一种”紧张”的颤抖紧张状态。愤怒可能将全局神经动力学推入充满活力的强力循环中,向外界发起攻击,等等。核心自我与大脑的许多其他部分大量连接——与各种感觉触发和调节反馈、运动功能、自主神经整合反应以及许多高级认知过程,尤其是皮层中线系统连接。
在这里,非线性动力学的概念,如混沌理论的”吸引子景观”(attractor landscapes)变得重要(Lewis, 2005)。例如,在人类中,原始过程情绪反应的强烈全身运动模式,如笑和哭,可以有效地促进喜悦和悲伤的情感变化(Panksepp & Gordon, 2003)。此外,由于不同物种的情绪基质细节必然存在大量的进化变异,我们也可以开始设想不同物种的感受变异,这取决于在大脑的心智景观中详细阐述的大规模本能吸引子景观的变异和复杂性。
重申一下,我们假设当核心自我(SELF)内的情绪系统以某种方式被唤起时,该情绪网络和核心自我参与了一个共同的功能,即创造原始情感和适应性行为。我们提出,当核心自我呈现出被唤起的情绪系统的神经特征(尚待测量,但已有尝试:Panksepp, 2000)时,核心自我就成为了情绪本能行为和情感意识在不同初级过程情绪中的共享神经基础。此外,由于位于皮层下中线位置的核心自我网络与皮层中线结构紧密相连,每个情绪系统被唤起时的神经特征可能会向上传递到大脑的许多认知区域,从而创造出各种高阶的第三级法则性(在人类中,也是高度个体化的)情绪过程,这些过程在人类事务中尤为重要(产生如羞愧、内疚、嫉妒、同情、共情等情感)。这些高阶的社会衍生情感可能反映了发展精细的全脑动态,仍然与基本情绪紧密相连;它们不太可能只是新皮层工作记忆区域内”信息处理”的变体,正如许多当代研究者似乎认为的那样(见第2章)。
再次强调,这只是一个假设性描述,使我们能够想象那些仍需详细研究才能理解其底层神经机制实际如何运作的全脑系统的活动。事实上,如前所述,神经科学目前缺乏研究大规模网络功能的工具。目前,在没有直接脑部操作(如局部脑刺激)能在动物中产生评估性”奖励”和”惩罚”效应的情况下,本能情绪模式的释放仍然是在动物大脑中唤起和监测情绪-情感过程的最佳方式。这一论断得到了大量数据的支持,但在情绪研究(或整个心理学)中尚未得到广泛认可。动物通常不被认为有任何情感感受,只有情绪行为,因为据说高级脑区以某种方式构建了使情绪对人类如此强大和有趣的强烈感受。
无论如何,根据现有证据的权重,原始情感是一个本能/自动过程而非认知/意志过程。例如,健康的大鼠不可能不害怕猫的气味,尽管不同动物在这种情绪反应上存在大量变异。此外,大鼠会很快自动学会对猫的气味产生联想,通过条件反射,它们在这些相关感知出现时也会变得恐惧。正如我们之前指出的,大鼠会学会害怕猫铃铛的声音、猫的视觉形象、它的叫声、猫气味持续存在的环境等等。这些都是基于大脑自动学习机制的非自愿次级过程反应。此外,像我们这样聪明的物种可以使用其强大的高级脑区来抑制或增强情感反应。例如,一个害怕的人可能通过想象快乐的场景来让自己振作起来。然而,这可能不会消除恐惧情感。如果初级过程情绪反应——恐惧(FEAR)——足够强烈,它将赢得这场战斗。例如,在得知孩子死亡的消息后的几个小时内,无论如何试图想起更快乐的时光,都几乎不可能感到平静。
我们提出,随着动物的进化和对各种生态条件的适应,它们需要比感官情感和源于身体内稳态失衡的情感更复杂的自动量规来指导它们的生存预测。虽然低等动物可能仅通过过滤水环境就能获得营养,但进化很快塑造出需要寻求(SEEKING)系统来驱使它们主动寻找食物和其他资源的动物。随着进化——生存斗争中的有效筛选器——为日益复杂的动物推进了额外的生存策略,其他情绪也随之出现。例如,雌性爬行动物几乎没有表现出母性关怀(CARE)。它们产下数百个卵(成为许多其他动物的食物,甚至可能是同一物种的动物),尽管大多数后代无法存活,但庞大的孵化数量往往能确保物种的生存。鲑鱼在产卵后不久可能死亡的一个原因是,它们贪婪的食欲可能对幼鱼的生存有害,进而影响物种生存;事实上,没有关怀(CARE)本能,它们很可能会吞食太多自己的小鱼苗。然而,当哺乳动物进化时,它们的身体产生的后代要少得多,这些后代需要养育照顾才能生存。如果哺乳动物的大脑没有进化出驱使它们养育幼崽的关怀(CARE)系统,每个物种都会灭绝。
然而,我们注意到,这个CARE系统有时会对我们所说的”残忍”视而不见。斑点鬣狗和黑鹰通常会生育超过它们能够有效养育的后代数量。兄弟姐妹天生彼此不容,它们通常会打斗直到其中一个死亡或从巢中跌落。父母很少干预。这不仅仅是适者生存;这更多的是运气的抽签。在鹰类中,几乎无一例外地是第一个孵化的雏鸟”赢得”这场比赛。它在生理上大概并不比第二个出生的更”适合”——它只是在发育上占有优势——稍微更胖一些,运动能力上更成熟一些。为什么这些生物要制造一个”备用品”——一个可消耗的婴儿呢?大概是因为其中一个无论如何都很可能会死亡的概率很高。如果第一个出生的死了,这个备用品只是”廉价的保险”。当然这只是一个理论,因为科学很少能以令人信服和严格的方式回答”为什么”的问题。当它处理”如何”的问题时,要令人信服得多。
尽管如此,情感出现的原因可能是因为它们在预测并经常保护生物体免受未来不利事件方面非常有效。如第2章所述,饥饿并不意味着一个人的身体能量危险地低;它只是强调在储存显著下降之前”补充”能量在进化上是明智的。通过这种方式,稳态情感(如饥饿)预测未来,并提供超越即时代谢考虑的动机。情绪情感提供类似的预期功能。例如,我们在面对可能的危险时会感到害怕,并在受伤或被杀之前采取预防措施。情绪情感采取现象体验的心理状态——带有价值的主观状态——这种形式的原因,大概是因为这是一种在不可预测的环境中以相对精确的方式激励和引导动物的有效方法。事实证明,情感是引导(“强化”和”惩罚”?)行为的理想工具,从而促进学习。我们倾向于重复那些之后带来情感愉悦的行为,并避免那些之后带来情感不适的行为。
我们提出,随着七种基本情绪系统的进化,无意识的原始自我也进化成了一个能够进行原始形式意识体验的初级核心自我(SELF)。我们说”原始”并不意味着它在体验上是微弱的。事实上,我们认为这些感受在心理显著性上是”巨大的”,后来可能随着皮层认知能力的发展而减弱,这些能力允许精确的行为调节,部分是通过抑制低级脑功能来实现的。认知决策虽然通常受到情感的引导(Damasio, 1999),但如果情感过于强烈也会被打乱。换句话说,我们认为正是核心自我,与各种基本情绪和动机系统交织在一起,产生了强烈体验的情感,随着额外的大脑复杂性——最特别是新皮层——的进化,这些情感继续提供至关重要的生存信息,从而允许更高认知功能的出现。如上所述,在这方面也许最令人信服的证据是,刺激这些古老的神经基质仍然会在人类中产生强烈的情感体验(Panksepp, 1985)。对所有情绪系统汇聚的大脑区域(如中脑的PAG)的广泛损伤会损害人类意识的所有形式(Schiff, 2007)。
我们倾向于这一观点的另一个原因是,最近的脑神经成像实验表明,当人们参与关注自己的活动时,在对各种形式信息的自我相关处理中,中线脑系统会被激活(Northoff et al., 2006)。确实,参与自我相关信息处理的中线脑系统形成了一个从内侧脑干到内侧额叶和扣带皮层的连续体,位于大脑皮层更古老的区域内。这个中线连续体的皮层下部分,即SCMS(皮层下中线系统),在所有哺乳动物物种中都具有显著的同源性,在其他脊椎动物的大脑中也能发现,包括爬行动物和鸟类(Northoff & Panksepp, 2008)。中线系统的皮层部分,即CMS(皮层中线系统),包括新皮层中一些与情感更相关的区域,包括岛叶、内侧额叶和眶额皮层。皮层中线系统在物种间的同源性不像皮层下系统那样明确,因为新皮层的大小和复杂性在不同物种之间差异如此之大。这再次表明,这个自我相关处理连续体的皮层下范围构成了一个规律性的情感核心自我,而更高的范围则贡献于日益个性化的认知自我。
虽然从同源性角度区分SCMS和CMS很有用,但它们在功能上是相互关联的,CMS依赖于SCMS来维持其完整性和存在。这些脑区的损伤会显著损害意识,而且损伤位置在这个自我相关处理的神经连续体中越靠下,缺陷就越严重(Merker, 2007; Panksepp & Trevarthen, 2008; Watt & Pincus, 2004)。例如,位于SCMS核心的PAG(导水管周围灰质)的完全破坏,会导致所有与自我相关的环境事件处理能力的丧失。当PAG完全受损时,所有指向外界的活动都会受到损害。动物勉强保持清醒状态,但它们似乎无法以任何有意义的方式感知事物(Bailey & Davis, 1942, 1943)。相比之下,较高级的皮质中线系统的损伤不会造成同样程度的意识障碍(Watt & Pincus, 2004)。
当损伤从大脑的内侧延伸到外侧区域时,严重程度也存在梯度差异。如果某人遭受外侧脑损伤导致失明、失聪或丧失语言能力,这个人仍然体验到自己是一直以来的那个人。他的基本自我感和情感体验没有受损,尽管他失去了一些宝贵的认知能力。相比之下,内侧脑区的损伤,特别是额叶区域,会更严重地损害自我体验(Northoff, 2004; Northoff & Bermpohl, 2004)。
接受过”裂脑”手术的患者也提供了相关发现。这是一种主要用于治疗严重癫痫的手术,通过切断胼胝体来实现。胼胝体是一组大量的神经轴突纤维,提供了新皮质两半球之间的大部分通讯。它本质上是包含高级脑两半球之间绝大多数连接的通道。新皮质的每一半处理不同类型的信息。左半球通常更具语言性、社交性和快乐,而右半球语言能力较弱,更情绪化和不快乐(Davidson et al., 2003; Tucker & Williamson, 1984)。这对我们人类历代构建社会的方式有着深远的影响(McGilchrist, 2009)。两个半球的功能不同,但通常它们处于某种协调状态。人们可能预期裂脑手术后,两半球之间缺乏交流可能导致功能不连贯。然而,并没有发生这种情况。例如,当裂脑个体跳入游泳池时,没有出现不连贯的行为迹象,比如身体一侧乱动,这表明半个大脑措手不及。
不过,在各种实验情境中,以及偶尔在现实生活中,那些分离的半球会表现出相互矛盾的行为。由言语性左半球控制的右手可能拿起一张纸,而由右半球控制的左手则把纸折叠起来放好。普遍认为右半球通常更情绪化(感受复杂问题),而会说话的左半球更具认知命题性(对复杂问题发表意见)。在认知层面,两个半球可能有不同的目标和意图。然而,裂脑患者的整体表现表明,他们继续表现出连贯的情绪、运动和全局性的全身意向行为,这表明自我和意识存在连贯性,因此必定植根于仍然以正常方式相互连接的皮质下系统(Panksepp, 1998b)。
在生命早期被去除皮质的动物行为中也可以观察到其他相关现象:正如我们反复强调的,它们保持了相当强的行为连贯性和自发性。它们甚至保留了在打闹游戏中与正常动物有效竞争的能力。因此,显然最先进化出来的皮质下结构,为自我提供了基础性的神经结构基石,而发育中出现的高级脑功能所产生的更多变的认知版本的自我则依赖于这一基础。由于皮质下中线功能在大脑进化的祖先熔炉中出现得更早,我们可以确信这些结构在维持心智连贯性方面具有优先地位。这种内在连贯性需要某种神经解释,而核心-自我(core-SELF)概念仅仅突出了我们可以在哪里以及如何寻求这个未解问题的解决方案。
我们承认我们的假设存在争议。许多认知和行为神经科学家可能不接受皮质下中线区域在产生情感方面比帮助介导人类思想和其他认知的高级皮质区域更重要(例如,Damasio在《笛卡尔的错误》中的观点,但在他最近的著作中可能不是这样:Damasio et al., 2000; Damasio, 2010)。LeDoux(例如,正如他在1996年首次阐述的那样)似乎仍然反对”低等”动物将情绪体验为情感的观点。事实上,大多数关于人类意识的讨论倾向于关注大脑心智(BrainMind)最近的进化赋予,认为它们对意识的产生最为关键。然而,让我们再次重申为什么我们必须得出结论(正如其他人也强调的那样),大脑的低级区域对于情感状态(Denton, 2006)以及原始知觉状态(Merker, 2007)实际上比高级区域更为关键。
本质上,三条有力的证据线索指向了皮层下深部区域对情感生成的关键重要性。首先,当人们电刺激大脑的特定内侧区域并唤起与大多数初级过程情绪相关的情感状态时,在刺激网络最低部位时总是需要更低的电流水平。例如,所有情绪系统都汇聚于PAG(导水管周围灰质),并且可以用最小的电流水平从这个结构获得最强烈的情绪反应。换句话说,这些环路在较低的脑区比在较高的脑区更加敏感或更加集中。其次,当人们电刺激下丘脑的局部区域并获得与RAGE(愤怒)和SEEKING(寻求)相关的情感状态时,通常的情况是行为和情感状态被这些环路较低部位的损伤削弱的程度大于较高投射区域(Valenstein, 1966),尽管较高的基底神经节,如扩展杏仁核,在向更高脑区传递奖赏价值方面很重要(Waraczynski, 2006)。第三,来自脑成像的最新证据表明,皮层下神经元唤醒信号与体验到的情感程度呈正相关,而较高脑区通常与认知性情绪体验呈负相关(Liotti & Panksepp, 2004a; Northoff et al., 2009)。这强烈表明较低区域主动生成原始情感状态,而较高区域可能在调节、再加工和抑制它们。同样,这种相互作用有助于解释较低脑区的唤醒如何可能干扰认知处理。
关于初级过程情感的皮层下控制位点的大部分相关因果证据来自动物研究。这在多大程度上适用于人类?当然,人类的脑刺激研究与我们的论点是相当一致的(Heath, 1996)。一个特别巧妙的相关证据来自在富含捕食者的恐怖模拟环境中的人脑成像(Mobbs et al., 2007):当捕食者在远处时,较高的FEAR(恐惧)区域(例如杏仁核)被唤醒,而当捕食者即将咬人时,较低区域(PAG)变得更加兴奋。(对手指的电击用来模拟捕食者。)换句话说,当你仍然相对安全时——当捕食者从远处潜行时只有轻微焦虑——只有较高的FEAR环路明显参与。毕竟,你只是在思考捕食者,而且你仍然有很好的逃脱机会。然而,随着电子”捕食者”逼近,PAG表现出越来越多的唤醒。而且从因果性脑刺激研究中得知,正是背侧PAG能够精心制造出心智血肉所能产生的最强烈的恐惧感受。同时,显然这个脑区对于生成积极情感心理状态并不那么重要。
因此,如果我们要向一位知识渊博的脑科学家提出一个深刻的神经存在主义问题,那会是这样的:出于你避免强烈负面情绪的自然愿望,你永远不会允许哪个脑区被局部脑刺激人为激活?如果我们的神经科学同事了解这个领域的跨物种证据以及足够的人类数据,他或她肯定永远不会选择新皮层甚至杏仁核的区域。相反,科学家会选择PAG的背侧部分,那里集中了FEAR(恐惧)、RAGE(愤怒)和GRIEF(悲伤)的心理痛苦环路。大脑中没有已知的更令人情绪厌恶的部位。例如,在这些区域受到刺激的患者可能会突然惊呼:“我快被吓死了!”(Nashold et al., 1969)。相比之下,腹侧区域没有那么消极,并且通常是积极的,突出包括LUST(欲望)和SEEKING(寻求)冲动。然而,PAG并不是唯一参与情感生成的皮层下中线结构。但它只是最重要的之一,可能也是最强大的(图11.1)。
尽管较高脑区对情绪调节至关重要,这是一个庞大而复杂的话题,我们在本书中不会详述(但可参见Harmon-Jones & Winkielman, 2007),但似乎很清楚的是,原始情感感受与相同皮层下情绪系统的唤醒密切相关,这些系统生成我们可以直接看到的本能情绪行为,以及相关的身体生理变化,如用复杂技术监测时的自主神经和激素唤醒。如果我们想要科学地理解生成强烈情感感受的因果机制,我们别无选择,只能实证性地研究大脑的某些较低中线区域。
综上所述,有两个主要理由让我们相信,现象性意识的核心自我体验可能由情感意识主导。首先,如果聚焦于皮层下中线结构的解剖学,可以看到导水管周围灰质(PAG)位于其基础的中心位置。PAG是一个所有情绪系统在某种程度上都会汇聚的结构,尤其是负性情感系统(Panksepp, 1998a, 1998b; Watt, 2000)。因此,我们可以将其视为一个为产生情绪性的神经系统提供大规模汇聚点的结构。如果我们假设皮层下中线结构是核心自我的基础这一点是正确的,那么PAG是这个系统的一部分这一事实表明,情绪可能在核心自我的功能中发挥重要作用。其次,皮层下情绪系统的唤醒会产生带有效价的好与坏的情感体验。这些体验已经通过一系列评估任务进行了监测,从寻求或回避脑刺激到条件性场所偏好/回避。如果情感由SCMS阐发,且SCMS是核心自我的基础,这表明核心自我是情感体验的所在地。
核心自我的神经进化基础是什么?有进化论的理由认为,核心自我可能由运动装置支撑,而原始核心情感意识如果没有这种与行动相关的核心自我脚手架就无法存在。首先,精密的运动能力可能在动物发展出让我们都能很好地看和听的复杂感觉-知觉装置之前就已经存在了。其次,归根结底,是身体行为决定了生存;感觉信息和内部情感变化的目的是引导运动系统。如果没有内部产生行动的能力,那么感觉信息就没有用处——例如,视觉让寄居蟹能够比被咬后才缩进壳里更早、更有效地缩进壳里。如果核心自我基础由这样的本能运动-行动系统支撑,而本能行动系统包含情感特性,人们也可以开始设想某些类型的感觉信息如何能够轻易地产生情感反应。
我们提出,自我相关的信息处理与大脑的本能行动倾向是整体相连的。当然,这并不是说感觉功能不参与其中,而是说核心自我是以行动坐标铺设的。这为自我表征提供了一个稳定的神经矩阵。当一个人在大脑中向上进行到更加个人化的自我时,感觉-知觉装置变得越来越有影响力。我们在上面提到,大多数人觉得原始意识植根于行动系统这一想法违反直觉。几乎每个人都认为意识是一种感觉功能,因为意识的内容往往是这样或那样的感觉印象。(我们的视觉、嗅觉、触觉、味觉和听觉能力是产生我们所思考的大部分内容的感觉功能。)然而,仅仅因为意识的内容主要是感觉性的,并不意味着意识的祖先基础必然与感觉-知觉装置紧密相连。毕竟,最重要的”感觉”是内感受性的,因此与自我相关。
也许大多数感觉-知觉能力应该被设想为对意识的高阶”觉知”功能的影响,而不是意识本身。显然,如果我们失明,我们不会在任何有意义的方式上变得更少意识。意识的整体质量更多地是被额叶损伤所改变,而运动功能集中在额叶。新皮层的额叶运动-行动区域大量投入到执行功能中,如注意力集中、运动规划、想象,以及更高级的社会情绪如内疚、羞耻和共情。显然,大脑的这些执行区域需要有知觉输入才能知道如何规划。这样,人们可以看到,意识的表面内容可能是感觉性的,而有意识的基本能力可能明确依赖于大脑的行动产生装置。
大多数研究者可能甚至没有考虑过初级过程意识与情绪-本能行动坐标整体相连这一想法,因为他们假设运动反应只是大脑的”输出”,因此运动装置只是支配像膝跳反射这样的无意识运动反射(这种思维方式在恐惧学习等领域尤其盛行,如第6章所述)。这样的大脑”纯粹”输出怎么可能构成核心整合原则?如果我们考虑得到充分研究的视觉功能,也许本能行动作为意识基础的这种关系会更容易想象。
众所周知,如果视觉感知不依托于探索性眼动(eye movements),往往会退化(Yarbus, 1967)。换句话说,如果不移动眼部肌肉,视觉能力就会下降。而且,正如我们将要展示的,上丘(superior colliculi, SC)中低水平视觉感知的稳定性依赖于稳定的眼动坐标。因此,眼动映射决定了上丘在何处处理传入的视觉感官信息,可能也决定了如何处理(Sparks, 1988)。上丘除了在视觉中的作用(即上丘最表层)之外,还在下方的连续神经层中与听觉和触觉整合——这是一个非常有趣的进化进程。触觉先于听觉,听觉先于视觉。这个区域可能是自我的核心结构(Strehler, 1991)。无论如何,视觉定向所需的动作系统位于上丘的最底层,正好位于导水管周围灰质(PAG)的上方。换句话说,上丘的最底层神经元构成了一个基本的运动定向系统,它在自我参照坐标中稳定并可能允许视觉感知,这也可能与PAG下方的初级过程情绪系统相互作用。
因此,有充分的神经生理学理由认为,与自我相关的视觉加工主要依赖于内在的视觉运动功能,而不仅仅是视觉感知装置。同样,在中脑中紧邻PAG的位置,上丘的最后一个神经元层控制着支配探索性眼动的运动能力。这些运动区域对世界的反应方式比上层感觉区域更稳定。例如,顶盖(tectum,上丘的外层)表面的神经网络在收集视觉刺激位置信息时表现得相当灵活。然而,视觉定向到刺激的确切方式决定了上丘的哪些区域处理传入信息。换句话说,视觉场景可以根据眼睛的定向在视觉感觉上丘上”滑动”。相比之下,产生适当视觉定向和搜索运动的底层运动系统使用一组非常稳定的动作坐标,无论处理什么类型的视觉场景,这些坐标都保持在相同位置(Sparks, 1988)。
这值得重复。当从上丘的视觉表面记录动物对视觉刺激做出定向眼动和头部运动时,刺激在上丘上的神经记录会改变位置;可以说,它们会”浮动”,这取决于动物刚刚如何定向。换句话说,视觉空间中相同的刺激会根据动物移动眼睛的位置而在上丘表面改变位置,从而改变其焦点注意力。相比之下,底层的运动映射始终保持稳定和可预测,当动物做出特定定向运动时表现完全相同。有充分理由认为,情绪动作网络的功能是通过进化以类似稳定的方式建立的。这些发现促使我们认为,核心自我(SELF)的情绪情感功能从根本上基于我们可以在动物行为中观察到的本能情绪动作系统。
从所有外在表现来看,所有哺乳动物都有这种稳定的”基础”,原始情绪感受本质上存在于它们古老的情绪动作系统中,无疑得到来自身体的反馈支持。我们可以确信的是,在更深处,在中脑最古老的区域,就在脑干上部的正中央,我们发现了PAG,所有情绪动作系统,尤其是负性情感系统,连同它们强大的情感能量,都汇聚在这里。这无疑有力地证明了情绪动作系统在心智进化中的首要地位。
如果我们把以上所有数据综合起来,似乎连贯的情绪动作系统在意识的产生中占据首要地位。显然,大脑需要稳定的机制来产生心理行为的连贯性。如果我们关于核心自我的概念是正确的,那么可以合理地假设,所有多样的”个性化”自我都依赖于深度情感性的核心自我的完整性。因此,情感性核心自我可能为所有更高级的自我功能提供了坚实的神经进化支架。
上丘的深层运动区域及其与之相互作用的PAG下层区域与额叶皮层运动区域的连接比与后部皮层感觉区域的连接更丰富。事实上,新皮层的额叶运动区域是产生计划和意图的地方,这表明大脑的这些心理特征与运动功能有关。在其他研究大脑活动变化的人类实验中,“拥有”活动的感觉与各种额叶皮层脑区的激活增加有关(Ehrsson et al., 2004)。此外,激活的幅度与主观所有权的程度相称。
新皮层的额叶运动区域是个体自我开始”孵化”的大脑区域,这是作为生活经历的函数,因为这些高级网络帮助在时间上建立行为优先级,基于感觉信息。长期以来人们就知道,当额叶/运动区域受损时,相比后部感觉区域的损伤,会导致更显著的人格变化(Elsinger et al., 1992; Passingham, 1993; Perecman, 1987)。因此,个体自我的构建是由额叶皮层的高级运动行为导向执行装置所精细化的。高阶情绪行动能力可能对产生情感色彩的个体自我功能(我们称之为人格)特别重要。
一个有趣的问题依然存在:高级皮层区域在多大程度上主动参与情感状态的产生?显然存在大量的自上而下认知功能可以激发和调节情绪(Gross, 2008)。正如每个明智的人都知道的,当情绪唤醒相对较低时,更容易在情感负荷的事项上做出合理的认知决策。相反,皮层下情绪唤醒倾向于在强迫性自我服务的思维轨道中激发和维持思考。因此,额叶的内侧情绪区域和外侧更偏认知的区域处于跷跷板平衡状态也就不足为奇了(Liotti & Panksepp, 2004b)。当人们以情感方式看待世界中的事件时,内侧额叶区域比外侧(认知)区域更加活跃。如果以更认知、更少情绪的方式看待相同的刺激,平衡就会逆转,外侧区域相比更偏情绪的内侧区域变得更加活跃(Northoff et al., 2009)。显然,强烈的情绪和理性不能共存。同时,也很明显,当需要做出选择时,感受对于下定决心非常重要(Damasio, 1994)。
我们必须注意到,最近一些复杂的情绪行为反应,如恐惧的做鬼脸,已经通过微刺激顶叶体感皮层区域的某些区域而被激发(Stepniewska, et al., 2005)。我们还不知道这些刺激是否能在学习任务中介导情感性奖励或惩罚。在这些问题得到解决之前,合理的看法是,它们是通过生活经验而非进化遗产在这些大脑区域中出现的情绪行为的习得性基底。
综合来看,核心-自我系统由本能回路的复杂网状结构构成,不仅产生情绪行为和相关的身体变化,还产生原始情感。基于这些理由,我们支持双面向一元论理论。一元论,与二元论相对,提出心智中发生的一切最终根植于单一实体——在这种情况下,是物理大脑,显然最根本地位于皮层下和皮层中线系统。双面向指的是这些中线系统同时产生情绪的两个看似不同的方面——连贯的情绪行为倾向和相关的初级过程心理状态(情感)。事实上,两者都是大脑中同一整合过程的反映,这使我们能够使用可以客观测量的事件(本能情绪行为)作为必须间接推断的主观体验过程(即本能情绪感受)的代理。因此,动物的本能愤怒行为反映了内在的愤怒情感。这使我们能够根据动物的外部行为直接解读其情感问题。我们知道产生愤怒的回路感觉并不好;如果有机会,动物会试图逃避这种刺激(Panksepp, 1991)。
我们并不是说这些系统对身体的许多状态、大脑其余部分的活动以及世界事件不敏感。它们显然是敏感的,但情感体验之谜的主要部分存在于所概述的内侧脑系统的动力学中。至少现在我们知道,如果想更详细地理解原始情绪感受,应该将努力集中在哪里。我们遗憾的是,从情绪感受的角度对这些神经系统进行的详细研究相对较少,这应该对理解许多心理健康问题和精神疾病的本质产生重要影响(参见接下来的两章讨论)。
人们还可以看到,核心自我是大脑中产生基本学习-认知的部分,可能主要是无意识的,因为它将情绪情感和行为倾向与基本感觉印象相结合。正如我们所讨论的,这些感觉印象(视觉、声音等)不必来自新皮层。它们可以完全通过皮层下感觉系统控制情绪学习([第6章]和[第7章])。核心自我还接收来自身体内部的信息。例如,内感受神经元可以检测到水、能量、热量和其他失衡,在核心自我中被情感性地表征为口渴、饥饿或寒冷。情绪系统也直接受到自主神经信息的调节,如血压和身体的各种激素状态。一些外部刺激,如捕食者的气味和疼痛,可以直接唤起恐惧系统——导致情感和行为倾向与简单知觉相结合而产生。但与世界事件的大多数联系最终是通过学习建立的。
将外部刺激与核心情绪自我(SELF)联系起来的过程,以及各种个人化自我(idiographic selves)的逐渐出现,肯定不是孤立的认知过程。就个人化自我的出现而言,复杂的认知过程——我们的想象力、计划能力以及对因果关系的理解——无疑参与其中。但它们总是伴随着对世界的情感评价(affective valuation),这使得高级脑系统在心理成熟过程中,能够与各种感知的效用(评价)保持联系,以便促进对未来行动方案的评估,从而增强每个个体的生存能力,并从长远来看,增强物种的生存能力。
在这一部分,我们将提出两个可能影响心理健康领域的观点。首先,作为人类,我们都有高度发展的个人化自我,因此很难理解以最基本形式体验自我相关处理会是什么感觉。然而,我们认为这可能会让人和动物感到他们在某种程度上拥有(Blakemore et al., 2000; Blakemore, 2003; Ehrsson et al., 2004; Jeannerod, 2003)他们生活的世界,他们是导致事情发生的主动行为者(Gallagher, 2000; Gallagher & Frith, 2003)。他们可能获得一种关于自己经历的我属性感(mineness)(van Gulick, 2004),并且他们可能将自己的情感价值投射到世界中的物体上。相反,他们也会将有价值的物体内摄到他们的内在生活中。这种拥有感可能让人和动物感到他们在某种程度上属于自己的世界——他们在情感上嵌入到他们的生活环境中(Izdebski, 2008; Northoff, 2004; Schore, 1994; Zinken et al., 2008)。
正如我们已经指出的,生命早期严重的依恋障碍与大脑皮层和大脑情绪网络之间神经交流线路的衰减有关(Schore, 1994)。这种衰减可能限制大脑皮层适当抑制和调节情绪表达的能力。我们知道大脑皮层的一个重要功能是抑制情绪性,因为这种强烈的情感唤起可能会破坏更高级认知处理的微妙形式。早期依恋障碍通常会导致成年后的人格障碍,患有人格障碍的人通常难以调节他们的情绪。出于这些原因,许多人认为神经连接的缺乏(这些连接本应允许大脑的一个区域有效调节其他区域的活动)可能是人格障碍的原因。简单来说,较高的中线区域调节较低的中线区域;额叶皮层的内侧和外侧区域也以跷跷板的方式工作。也许额叶执行区域和后部感知区域的皮层具有某些相互依赖性,这对于健康和快乐的上层心智发展是必需的。
虽然我们认同这一论点,但我们提供了一种理解人格障碍和相关精神状况的辅助方式。我们注意到,对皮层下中线系统的重大损伤,例如完全破坏导水管周围灰质(PAG),会导致自我相关信息处理的严重减少。遭受这种损伤的人和动物不会有意义地保持意识。然而,对皮层下中线系统、对核心自我(SELF)的更轻微或更微妙的损伤,可能会对人或动物的情感定向感产生有害影响。嵌入到自己世界中的感觉可能会受损,从而削弱成为主动行为者的能力,这种行为者能够对重要的个人经历主张我属性感/拥有感。这些正是人格障碍患者抱怨的心理特征。因此,除了皮层抑制失败之外,皮层下中线系统内的神经失衡也可能导致人格障碍以及许多其他精神问题(Koenigsberg, 2010; Stein, 2009)。
与心理健康相关的第二点关注这样一个事实:皮层以及可能也包括皮层下中线系统具有高度活跃的静息状态,或者通常所说的高度活跃的默认模式网络(default mode network)(Damoiseaux et al., 2006; Fox and Raichle, 2007; Fransson, 2006; Raichle et al., 2001)。这种高静息活动可能反映了内部沉思(ruminations),当一个人专注于外部需求时会减少。因为大脑的这些中线系统介导自我相关处理,我们假设默认模式网络的高唤起不应该被外部认知刺激进一步增加,而只应该被内部产生的情绪材料增加。事实上,这在容易沉思自己问题的抑郁症患者中已被发现(Northoff, 2007)。这些发现有助于解释为什么大脑在高水平情绪唤起期间可能无法轻易整合传入信息,以及为什么认知处理在这种时候会特别受到损害。从心理学角度来说,这并不令人惊讶。我们都从个人经验中知道,当我们处于情感唤起状态时,我们无法高效思考。然而,上述发现为此提供了一个已确立的大脑机制,允许对潜在的大脑过程进行更详细的分析。
更有趣的是,高静息状态表明自我相关处理可能是一个持续的过程/活动。这种持续的自我相关处理可能使我们能够维持一种连续的、在时间上延伸的关联感。这样,正常的高静息状态活动可以被视为一种”生理基线”,它指示着心理健康(Northoff and Bermpohl, 2004; Northoff et al., 2006)。因此,更深入地研究这种静息状态如何被各种类型的强烈内部情绪唤醒所调节,以及当一个人以不同方式情绪激动时,这些CMS结构中是否会出现不同的神经动力学模式,将是非常有趣的。如前所述,这些大脑功能在抑郁症中明显过度激活,这可能突出了抑郁症患者具有极端的、负面情感性自我关注,可能通常由GRIEF系统的持续过度激活所引发,从而放大了心理痛苦。我们预期,在情绪激活期间,这种增强的神经活动也会在皮层下中线区域表现出来。
开发更直接的此类心智大脑变化的神经测量方法将非常重要。因为fMRI无法直接监测神经活动,所以拥有大脑激活的直接电信号测量而非血流变化等次级效应也是可取的。唯一适用的技术仍然是有史以来发现的第一种脑成像类型:脑电图,以及最近它的高级版本,脑磁图。当使用位置良好的电极从CMS和SCMS记录这类研究时,我们预期它们将被证明是可以从人脑获得情感特异性神经元特征的最佳大脑区域(Bekkedal et al., 2011; Northoff et al., 2009)。
对意识感兴趣的哲学家无法逃避考虑自我的本质。例如,尽管笛卡尔在探索意识现象时没有明确谈到自我,但他仍然假设它的存在。他的”我思故我在”宣言只使他有权确认思考的存在,这是一种产生对自己经验的觉知并因此思考世界事件和个人在生命流动中位置的、脱离身体的高级意识形式。但他假设有某种东西,即他的自我,在进行思考(Copleston, 1962b)。因此,笛卡尔含蓄地接受了意识的存在,连同一套连贯稳定的自传体记忆,意味着自我的存在。当然,他并未将其设想为严格的神经生物学过程,而是与宗教概念化的灵魂的非物质方面更相关的过程。因此,他选择在物质和精神领域之间做出明确的界限,将精神(灵性)领域仅保留给人类。这一选择从那时起就损害了心智大脑科学的进步。
在本章中,我们提出了关于自我本质的另一种假设,这可能有助于解释哺乳动物大脑中如何产生情感。我们假设的核心是断言大脑中的中线系统,它赋予所有哺乳动物一个普遍的(规律性的)核心SELF,可以在与更高信息处理相关的大脑其他区域中支持自我的各种表现(个体性形式)。越来越明显的是,中线系统介导各种自我相关的信息处理(Northoff et al., 2006)。我们设想自我最初进化为一个同源的规律性核心SELF,它帮助大脑的其余部分发展出更多的个体性自我形式,特别是在像人类这样高度大脑化的物种中明显的巨大多样性,但在低等哺乳动物中并非不存在(尽管如果动物是基因克隆,多样性会大大减少)。额外的解剖学和实验数据使我们能够假设,初级过程情绪系统在核心SELF的功能中发挥关键作用,以及对世界的习得性评价,最终随着更高皮层的参与,导致多样化的个体性自我。在此基础上,我们提出当各种情绪网络被唤醒时,中线系统呈现出不同类型的神经元放电模式时,就会产生情感。
情感系统持续评估外部和内部世界。此外,由于其进化设计特征,原始过程情感总是相对于个体和物种的生存来评估内部和外部世界。因此,产生和调节情绪性的中线系统持续参与对外部信息的自我相关(“这对我有什么好处?”类型)处理。通过这种方式,我们可以再次设想所有哺乳动物如何成为”主动”寻求信息的生物,而不仅仅是”被动”整合信息的生物。这就是为什么将中脑边缘多巴胺网络概念化为参与阐述寻求系统(SEEKING system)的行动冲动,而不仅仅是产生诸如”想要”和”激励显著性(incentive salience)“之类的高阶过程或成为”大脑奖赏或强化系统”,这样做要明智得多的原因之一。寻求概念可以解释这样一个事实:在大量不同的情绪情境中,感觉刺激变得更能吸引接近行为和集中注意力。它还有助于解释为什么人工激活寻求系统是”有益的”,但不是在促进愉悦感觉的典型意义上。它促进对世界的热情参与以及对世界的食欲式渴望,特别关注世界上那些预示欣快兴奋或满足和愉悦的刺激。
我们回顾了一系列数据,表明这些支持自我相关情绪处理的中线系统从根本上基于内在的行动-运动过程,而不仅仅是感觉输入到某种认知分析器中(如所有读出理论所假设的那样)。这意味着情绪唤起同时导致情感的产生和行动倾向的产生:肌肉张力的改变、自主反应以及相对刻板的情绪行动冲动,如接近或逃跑、以各种方式进行社交参与或退缩。因此,我们支持双面一元论(dual-aspect monism)理论,该理论指出,皮层下中线情绪系统通过一个连贯的整合系统同时产生各种行为、生理和情感性的情绪表现,用于自我(SELF)表征(Panksepp, 2005b, 2009b)。
最后,我们注意到,当清醒的动物没有发生其他事情时,中线系统以高静息状态运行。这使我们能够考虑这样一种可能性:情绪健康的动物和人类通常进行持续的自我相关信息处理,这在抑郁症等疾病中可能变得极端——充满负面情绪和自我厌恶的沉思。通过大脑中情绪系统的存在,最特别的是古老的寻求系统(所有积极社会情绪系统的鼻祖)参与了许多其他情绪反应(例如,由恐惧系统(FEAR system)促进的”寻求安全”),动物成为其世界中的主动代理者而不是被动的僵尸。随着更高的心智脑(MindBrain)进化,这意味着用情感性的心灵之眼看世界,用耳朵倾听情绪细微差别,并拥有最大多样性的感受,最强烈的时候是当一个人与自己的同类以及具有相似情绪系统的进化相关动物处于各种形式的社会情绪接触中时。人与动物的联结很容易与伴侣动物形成,这强烈基于我们共享进化相关的社会情绪系统这一事实。
大脑中主观性的存在也意味着我们不能简单地用传统的科学第三人称工具来研究大脑功能。心智不是像田野中的石头那样的中性事物。它有一个观点,一个我性(I-ness)。每一个真正重要的观点都强烈带有情感色彩。要看到脑心智(BrainMind)的真实面貌,我们必须在神经科学术语中越来越有效地处理主观性。当我们对心智进行准确的考古学研究时,我们在心智的基础上发现了情感体验。令人惊讶的是,情感仍然是大脑功能中最不被理解、研究不足和讨论不足的功能之一。情感是大脑处理外部”奖赏”和”惩罚”时所做事情的核心。情感变化可能是”强化”在神经系统中创建学习和记忆的方式。正如第一作者将在下一章中讨论的那样,我们未能以这种方式看待脑心智,对现代生物精神病学的成熟以及对心理治疗如何能够帮助恢复情绪平衡的更好、更连贯的理解产生了深远的负面影响。
我们越深入外部宇宙,就越接近我们的起源,回溯到时间的开端。我们越深入无意识的暗物质,就越了解我们的起源和现在……我们就能更好地预测未来的问题,无论是焦虑、药物成瘾还是性功能障碍。
——Arthur Janov (2007, 第17页)
本章是倒数第二章,目标是整合迄今为止涵盖的各种思路,以便从我们对基本情绪系统日益增长的认识中提炼出一些心理治疗的启示。在这个层面上,需要处理一种尚未与临床思想整合的对脑心的新理解。这里的目标不是给出建议或做出确定性声明,而是探索我们对心智情绪基础的新理解为治疗工作提供的多方面维度——无论是在咨询室的人际动力学中(Siegel & Solomon, 2012),还是在生活本身的创伤领域(Belenky, et al., 1996)。
我的观点是,通过心理、躯体和生理方法直接操控初级过程情绪回路,可以在情感障碍中取得显著的治疗效果。更传统的观点认为,持久的改变只能通过语言的途径来处理情绪动力学——通过三级认知过程的中介来处理个人生活事件。当然,所有心理治疗师都接受这一观点,但也有一些人相信,对于许多情绪疾病,特别是那些源于早期发展问题的疾病,还必须更直接地处理潜在的情绪动力学,有时甚至要在非语言的初级过程层面上工作(例如,Janov, 2007)。虽然所有人都认同心理治疗按照传统概念必须通过人类心智的语言途径进行,但有理由相信,心理治疗学的下一次革命将来自新的神经精神分析视角,以及使用多模式方法更直接地操控情感脑心功能。为了结束这个叙述,最后的”尾声”章节将在历史相关的哲学视角中框定本书的目标。
本书的一个前提是,我们越深入情感基础的深处——我们的内在宇宙——就越接近我们的心智起源。在某个时刻,当我们下降到大脑更古老的深处时,可能除了创造纯粹行为的无意识神经网络什么都没有——生物体四处游走却没有感觉——也许就像起伏的水母随着潮汐漂浮,带着最微弱的前意识形式。也许我们脊髓中的神经网络就是这样:深度无意识的。我们只是不知道,也没有好办法找出答案。但我们终于可以理解原始情绪感受的本质——它们产生于与无条件情绪行为和反应相同的脑区。理解这些过程,为进一步的进步提供了坚实的基础。理解情感动力学如何在哺乳动物大脑中产生,可能是精神病学和意识研究以及试图恢复情绪平衡的心理治疗师面临的最重要的科学问题。说得委婉一点,这个领域的历史是混乱的,有许多突出的死胡同,例如詹姆斯-兰格新皮层解读情绪理论(James-Lange neocortical readout theory),它仍然处于许多心理学家理解的前沿,尽管没有持续的批判性研究线来支持它。充其量,它目前应该成为本书讨论的大脑内主要原因背后的一个次要主题。许多其他人相信情绪,确实是情感感受,可以是动态无意识的。也许吧,但这可能只在感受被过度认知活动否认或压抑时才会发生,这是人类心智的常见倾向,确实可以在某种程度上抑制皮层下的情绪动荡。但这些心理压力会以意想不到的方式渗出,并在人们的生活中制造混乱。
对一些心理治疗师来说,根据目前对心智祖先的分析,传统的无意识概念(由西格蒙德·弗洛伊德引入)并非完全”无意识”——它并非完全没有体验,这可能会令人惊讶。真正深度无意识的是大脑的自动学习和记忆过程。弗洛伊德的动态无意识或前意识(他含糊地使用这些术语)据说部分由本书描述的情绪状态构成。但这些状态,当足够强烈时,会被情感性地体验,尽管不是反思性地(认知性地)体验,不仅人类如此,其他许多动物肯定也是如此。我们现在可以确信,其他哺乳动物确实体验到它们的情绪唤醒——尽管大多数像新生儿一样,可能没有反思性地意识到它们正在拥有这样的体验。这就是证据现在表明的,值得记住的是,弗洛伊德也经常声称情感从来都不是无意识的。处于原始情绪状态中是有感觉的。它们是原始的情感体验——特殊的心理现象状态,一种独特的感质类别,产生于意识心智的根基。
尽管在哺乳动物中发生的强烈原始过程情绪唤起可能永远不会未被体验到——永远不会是无意识的——但学习的次级过程机制,即控制的下一个层级,是深度无意识的。这一点值得重复。据我们所知,学习和记忆反映了神经机制以确定性方式运转,将我们的原始情感与世界事件连接起来。因此,在更高的三级过程脑区——新皮质层,会出现复杂的意识形式——具有非常不同的心智模式(更纯粹的自我和世界的认知表征)——不同于皮质下区域的祖先性、原始性的注意、情绪和动机领域,在那里情感状态占主导地位。它们是在没有预先思考的情况下内在地预测未来的进化解决方案,这源于它们对新皮质编程的深远影响。
我们更高级的心智活动是深度认知的,因为新皮质脑区(总是与较低的脑心功能协同工作)从不同的感觉通道构建世界的图像,使我们能够与外部事件保持联系。一个神经科学和遗传学上站得住脚的观点是,新皮质在出生时基本上是白板(tabula rasa)——一种随机存取存储器类型的空白状态——大多数伴随成熟而来的高度可预测的功能特化是皮质下特化通过表观遗传的发育魔力编织出可预测类型的皮质”模块化”的结果,以及大量文化引导的学习。因此,我们的自传式知识和记忆库出现了,其中大部分是在SEEKING系统的指导和动机影响下形成的。
如果没有清晰的视角了解低级情感心智和高级认知心智如何相互连接(主要在内侧皮质和皮质下基底神经节区域:见[图1.1]、[1.2]和[3.1]),我们就无法清楚地讨论心理装置出现精神病学意义上的问题意味着什么,以及如何开发新的脑心疗法。很可能大多数行为和认知疗法之所以有效,主要是因为它们能够调节情感并更好地协调认知观点与积极情感。直到最近才出现了关于更直接的动态情感疗法潜在效用的丰富讨论,在这种疗法中,个体的情感生活被置于核心位置。
未来我们必须认识到情感过程皮质下定位的压倒性证据,这已经通过总结的局部脑刺激工作得到了如此戏剧性的展示,以及Damasio及其同事(2000;见[图12.1])的出色脑成像工作。在人类从自己的自传式记忆库中诱发情绪唤起期间看到的所有许多大脑变化中,当人类体验我们所说的RAGE(愤怒)、FEAR(恐惧)、GRIEF(悲伤)和joyfulness(快乐)(PLAY(玩耍)?)时,绝大部分唤起是皮质下的(见[图12.2])。如果说有什么不同的话,皮质区域在情绪唤起期间往往会关闭。显然,要理解情感大脑,我们必须理解神经发育的进化分层和整合,当然还有各个控制层级之间广泛的相互交织(我在[第2章]中讨论的嵌套层级,见[图2.1])。
这种对脑心的进化多层次视角对精神疾病及其疗法(神经化学和神经心理学)都有影响。在这里,我将简要探讨这些知识对理解人类情绪问题以及开发旨在帮助重建情绪稳态的新临床干预措施的一些意义,当生活的变迁和脑心的情感失衡对人们变得极其困扰时。
在这样做时,重要的是要记住,动物研究几乎没有告诉我们任何关于伴随我们情绪唤起的记忆和思想的短暂流动。但与此同时,对哺乳动物心智的情感神经科学方法在因果层面上告诉我们关于大脑如何实际产生情感感受以及哺乳动物大脑的深度无意识学习和记忆过程如何实际运作的大部分我们所知道的。然而,获取其他动物的高级心智体验仍然是一个科学上无法处理的问题。巨大的多层次脑心互动在我们试图理解精神障碍时产生了大量的复杂性层次——构建清晰的词语图像、精神疾病的概念符号描述以及用于改善人们生活中不受调节的情绪的破坏性的疗法的影响。
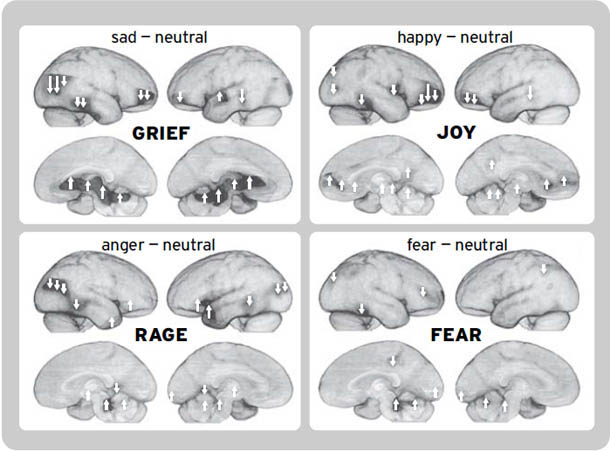
图12.1. 当人类在PET扫描期间体验四种基本情绪:悲伤、快乐、愤怒和恐惧时,大脑唤起和抑制的概览(基于Damasio等人,2000)。在这些情绪的每一种中,不同的低级皮质下脑区表现出明显的唤起,而各个皮质区域存在皮质抑制(血流减少)([图12.2]中的定量数据)。由于色码变化在这些黑白版本上难以看到,向上箭头表示皮质下唤起增加,向下箭头表示区域新皮质唤起减少(数据由Antonio Damasio慷慨分享;彩色图见Panksepp,2011b)。
本章的有限目标是探讨情感神经科学知识与特定心理治疗问题的相关性,同时不时插入到目前为止所阐述的情感心智(emotional mind)情感观点的概要片段。我还希望强调可能预防和促进未来情绪问题的发展性视角——即积极情感如何成功抵消消极情感。越来越清楚的是,情绪韧性可以通过育儿实践、持续的积极人际关注,以及贯穿整个生命周期的各种经过时间考验的(例如精神分析)和新兴的临床干预来提升。当然,早期经历长期以来一直被认为对长期心理健康问题具有决定性的重要性,现在神经机制正在被阐明,最显著的是通过临床前(动物)研究。
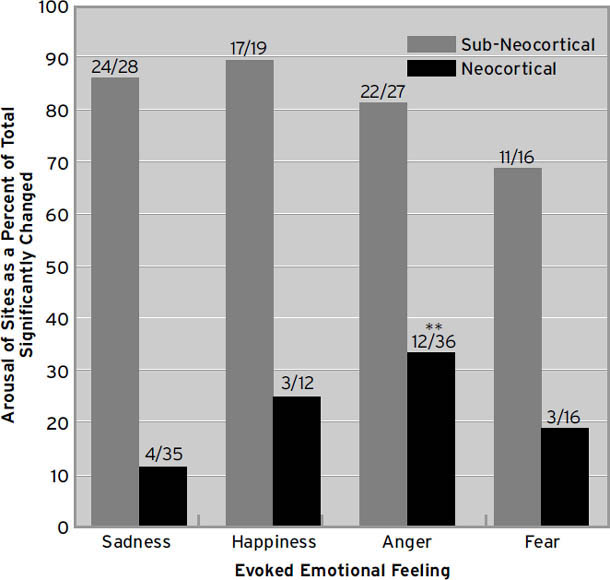
图12.2. 对图12.1中提供的数据的总体总结。显然,当人类体验基本情绪时,皮层下唤醒优于表现出抑制的区域。愤怒显示出最大的新皮层唤醒总体百分比,但每种情绪仍保持一般的不对称模式(数据摘自Damasio等人,2000年)。
因此,情感神经科学对所有心理治疗师和父母都可能具有相当大的用处,特别是那些关注健康儿童发展的人。人们应该理解,儿童天生具有某些情感能力,这些能力对他们的生活质量至关重要(见Sunderland,2006)。这样的知识将促进更好的育儿实践,其中(i)儿童的情感生活成为帮助父母了解婴儿茁壮成长所需条件的核心问题(Narvaez等人,2012; Worthman等人,2010);(ii)家庭中的积极依恋动力成为帮助儿童茁壮成长的关键(Code,2009; Hughes,2007);(iii)对我们情感生活的现实愿景,伴随着正念的实现潜力,促进积极的个人转变(Siegel,2007,2010)。
在前11章中,我们概述了情感神经科学中出现的关键科学问题,并提出了一些临床思考。我现在将讨论(i)如何将对初级过程情绪的理解纳入精神疾病的进化动物模型中,(ii)如何理解初级情感过程为精神病学和心理科学提供新的基础,以及(iii)这些新的理解如何为开发比以往任何时候都更直接针对情感的生物学和心理治疗干预提供新方法的基础。
我将在这里讨论的关键问题包括:大脑内产生的原始情感体验与情绪障碍有何关系?这些知识对实现情绪稳态、增强幸福感和对生活持更积极态度有什么意义?当然,对其他动物的情感神经科学研究如何能让我们更好地了解人类的情感生活?
有许多新策略等待在临床和临床前(即在动物模型中)进行评估。我们如何利用对积极情感系统日益增长的知识来对抗以消极情感为特征的障碍(例如抑郁症)?例如,我们可以设想许多新方法来使用SEEKING(寻求)和PLAY(游戏)的积极情感来对抗抑郁症和焦虑障碍的消极情感。随着我们可以通过动物的情感发声和本能情绪倾向相当直接地监测它们的情感感受的发现,在开发新的药物和心理治疗干预方面,动物研究和精神病学实践之间的智力交流可以得到丰富。然而,我们不应低估跨学科整合所面临的挑战。在继续讨论临床问题之前,让我们考虑一下阻止临床和基础科学问题关于我们原始情感心智本质更充分相互渗透的困难。简而言之,相关神经科学(从行为到分子)目前需要但没有对脑心(BrainMind)的初级过程情感基础设施的清晰愿景。事实上,出于愚蠢的历史原因,很少讨论动物情感感受及其对精神病学的影响的话题。在心理学中,大多数讨论都在三级过程层面,在那里认知和情绪不可分割地混合在一起,导致非常困难、有时混乱的讨论,其中很少有概念有神经科学依据。如果我们尊重控制脑心的分层循环因果影响,首先是自下而上,然后是自上而下,就可以获得更多的清晰度(见图2.3)。
在过去的许多著作中,我论证了心智的原始情感基础是深刻的生物性和皮层下的。指导原则是,原始情感源于产生本能情绪行为的大规模神经网络动力学,而非介导认知意识的更高级、与自我相关的感知脑区。原始情绪感受与情绪行动动力学密切相关,构成了不同类型的心理体验。由此产生的情感动力学也具有特征性的全身感觉:它们是初级过程的现象-体验状态,主要依赖于大脑内的内脏身体表征(核心自我),产生原始的”情感意识”,其次依赖于自主神经唤醒,这些唤醒可以被更高级的心理装置间接体验。因为这些皮层下动力学——大规模模拟网络功能——是情绪生活的原始源泉,心理治疗师需要清晰地设想这些心理能量的本质,以便在心理治疗中更直接、从而更有效地处理复杂的人类情绪问题。
分享和讨论我们对这些系统的新兴知识,这些系统存在于所有人类大脑中,其本身可能对那些处于情绪困扰中的人来说是一种有价值的治疗性洞察,他们有时对正在发生的事情感到束手无策。仅仅告诉陷入困境的个体,他们看似自由漂浮的情绪困扰是”真实的”,即使带着同理心这样做,也可能仅仅被理解为模糊和缺乏实质的安慰。解释每个人都有一组独立的情绪感受系统,这些系统是所有人共享的独特大脑过程,并且这些系统在所有哺乳动物大脑中都存在,有着充分而重要的理由,这是可靠的知识。而且,作为一般规则,许多人更愿意在获得同理心安慰的同时,得到建设性的信息。在更高级的心脑中,情感和认知可以富有成效地协同工作。它们也可以发动全面战争。它们是我们意识的两个截然不同的方面。最终,所有精神障碍都在这两个层面上表现出来。如果改变情感,认知往往会随之改变,特别是在良好的建议下。改变认知也可以奏效,但如果情感不随之改变就不行。认知和情感的相互作用使这变得困难(见图12.3)。
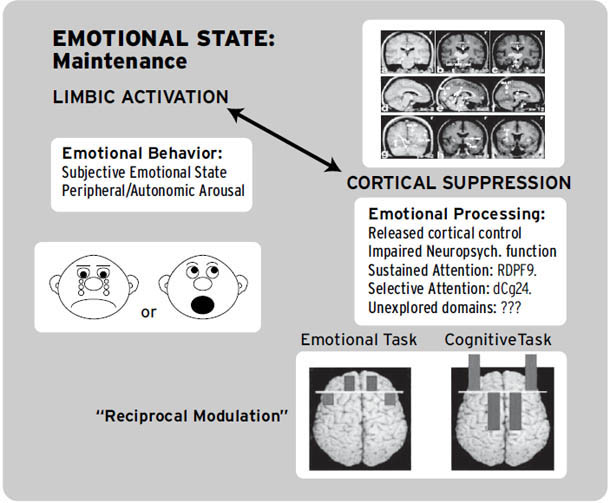
图12.3. 人类大脑中新皮层和皮层下唤醒变化的一般模式总结,作为情绪状态激活的函数。总体而言,每当情绪被唤起时,皮层下边缘系统唤醒具有一致性,而新皮层唤醒降低(总结图来自Liotti & Panksepp, 2004)。右上角插图突出显示了男性大脑在性高潮期间表现出的大脑唤醒示例。实际上所有唤醒都在已知调节基本男性性行为的皮层下区域,如图7.2所总结。(PET扫描由Janniko Georgiadis及其同事慷慨提供。)右下角插图是人们在以情绪感受心态(左)和认知分析心态(右)观看相同情绪图片时的fMRI数据总结;在情感观看期间,存在中线额叶唤醒和侧向工作记忆抑制,而在以更认知分析的方式观看相同材料时,模式发生了戏剧性的逆转(fMRI脑扫描总结由Georg Northoff慷慨提供)。
恕我重复这一点,了解本书描述的七个基本情绪系统产生的初级过程状态,对于情绪障碍临床思考的神经科学和进化基础至关重要。尽管这些系统的大多数细节仍有待探索,但已经建立了一个情感神经科学基础设施,允许基本情绪及其强大的情感指导临床思考。在这里,我将重点关注这些知识对特定情感治疗问题的影响,特别是从发展视角出发。
现在可以设计新的情感平衡疗法(ABT),用于重新平衡”心”而不仅仅是”头”,用隐喻来说——新的ABT的持久目标应该是在个体的原始情感生活中,追求更直接、更精确的有益干预。这显然应该包括认知重构,但通常仅靠这一点不足以最佳地重建情感稳态。简单地说,心理治疗师希望减轻甚至”治愈”心理生活中引起痛苦的不愉快问题。如果出现问题的是近期和特定的生活问题组,并且有明确的认知诱因,那么认知行为咨询是理想的进行方式。另一方面,大多数严重的情绪问题并不仅仅源于近期事件。它们的病因可以追溯到持续的过去压力源和创伤性变故,通常可以追溯到生命早期,如此之早,以至于几乎没有明确的记忆痕迹留存——只有失衡的情绪状态和相关的认知偏见。
这些早期印记可能会持续相当长的时间,因为极度紧张的生活经历会使情绪系统变得敏感或迟钝,产生永久性的、表观遗传诱导的高压力反应性和过度的初级过程消极情绪。我们还必须认识到,对年幼心智来说极度紧张和美好的事件,对成熟的成年人来说可能并非如此。不同年龄的相似压力源会产生不同的影响,这取决于关键的成熟期问题,以及每个人的遗传”易感性背景”——原始气质(primal temperament)。经典的气质类型——胆汁质、多血质、抑郁质和黏液质——并没有偏离太远,尽管我们新的情感神经科学人格量表提供了对这些情绪优势和脆弱性更客观的评估(Davis等,2003;Davis & Panksepp,2011)。需要为青少年和更年幼的儿童开发类似的测试,因为它们可能对教师和辅导员非常有帮助。例如,年轻人渴望在生活中拥有充足的真实游戏的强烈愿望常常被低估。强烈的愤怒和不切实际的恐惧需要被了解。它们需要被表现出来。它们需要被小心地讨论。
下面,我将重点讨论对初级过程跨哺乳动物情绪系统的理解如何指导我们理解情感体验在精神疾病发生中的作用,以及指导开发新疗法来减轻人类痛苦。但这些问题在优化治疗关系方面也是最重要的关注点。许多研究发现,来访者-治疗师关系的情感质量,而不是采取的具体治疗方法,可能是心理治疗结果中最重要的整体变量(Lambert & Barley,2001)。这主要是因为情感协调的治疗师与处于心理痛苦中的来访者分享真正积极的社交情感的需求至关重要。更好地理解我们内在的亲社会情绪也可能有助于阐明心理治疗过程的情感维度如何运作。心理治疗可能是放大的关爱系统(CARE)。有效”治疗联盟”的关键可能在于高阶共情共鸣(empathic resonances),其基础可能是我们大脑的关爱回路,它能很好地适应悲伤系统(GRIEF)和游戏系统(PLAY)的细微差别。
这引出了一些有趣的问题,比如许多积极情绪对心理和身体健康的影响。治疗师是否需要找到那些罕见但珍贵的时刻,在那时痛苦可以转化为快乐?人们能否通过简单地增加积极情绪的机会来对抗脑心(BrainMind)中的消极情感?到目前为止,这种策略在动物身上确实有效(Burgdorf等,2011),我们自己的一些最佳想法现在正在临床测试中,这些药物是通过认真对待动物的情绪而开发出来的(Moskal等,2011)。随着我们更好地理解这些大脑系统,我们可能能够设计出更有效的临床干预措施,让各种积极的社交情绪增强心理资源,并帮助重塑困扰的记忆——在积极情感的慰藉中重新巩固(reconsolidate)心理痛苦。
从理解进化赋予我们促进生存的情绪工具中,可能会产生各种新的治疗见解。情绪障碍总是与一个或多个基本情绪系统联系在一起。显然,消极情绪(愤怒系统RAGE、恐惧系统FEAR、悲伤系统GRIEF和枯竭的寻求系统SEEKING资源)在精神问题中很常见。过度的积极情绪也可能导致问题,在构建新的治疗方法方面同样重要。尽管目前关于情绪原型与精神问题关系的证据是推论性的,以下是一些可能的临床意义的简要概述,直截了当地陈述,没有广泛分析,针对本书中描述的每个系统。在这个叙述中不可能保持任何水晶般的清晰度,因为对于关键问题没有普遍接受的科学术语,而且所有系统都相互作用,并与更高级的脑心功能相互作用。因此,关于潜在功能细节的许多内容仍有待科学记录。实际上,整个章节都是作为思考的素材提供的:
寻求(SEEKING)冲动参与所有情绪唤起,激发并引导我们寻找资源。这个系统与高级脑功能一起,为人类寻求自我认同和生命意义的毕生发展提供能量。这个系统过度和失衡的唤起可能促进各种成瘾行为,并导致妄想性思维和偏执倾向——一种”过度”状态,这通常是躁狂症的特征。当被唤起时,以尚未完全理解的方式,它可以促进创造力;而当过度时,则会导致明显的精神分裂症和自大狂。当这个系统耗尽资源时,就会出现快感缺失和抑郁。调动和积极利用这种心理能量可以在一段时间内帮助缓解抑郁。因此,这个系统在许多积极和消极的心理结果中都起着重要作用。从积极方面来说,它促进与世界的成长性互动;从消极方面来说,当过度活跃时,它会促进成瘾、强迫症、精神分裂症和躁狂症,而当系统长期失调或活动不足时,则会导致空虚、疲惫的抑郁。例如,抑郁症的部分心理迟缓是由于抑制寻求(SEEKING)驱动而产生的过量强啡肽诱导的烦躁不安。相比之下,阿片类药物或内源性阿片肽的轻度刺激可以温和地刺激这个系统,产生愉悦和满足的感觉,并抑制各种负面情绪。因此,能够拮抗强啡肽同时促进μ阿片活性的新药,如丁丙诺啡,应该是非常有效的抗抑郁药,特别是当精神痛苦突出时(Watt & Panksepp, 2009; Panksepp & Watt, 2011;参见第9章)。
当群体或个人试图限制他人的愿望和抱负时,愤怒(RAGE)在社会问题中占据重要地位。这个系统持续唤起会导致慢性易怒和爆发性攻击障碍。当个人与更大的群体之间相互冲突的信念和目标发生碰撞时,愤怒也是人类互动中日常挫折的一部分,甚至会促进精神分裂妄想。愤怒(RAGE)像所有其他情绪一样,需要寻求(SEEKING)系统的参与。精神病性愤怒是最难治疗的情绪问题之一。与所有形式的破坏性愤怒一样,在所有哺乳动物物种中,雄性造成的破坏最大,鬣狗可能是例外(因为雌性通常有更多睾酮)。在人类中,几乎所有大规模谋杀都是由男性实施的。由于P物质是动物模型中防御性愤怒(RAGE)的明确促进剂,药理学阻断这个系统可能被证明是一种强大的抗愤怒药物,特别是如果同时减少患者社会环境中引发愤怒(RAGE)的挫折。
恐惧(FEAR)(及其产生的各种焦虑)是许多人永恒的伴侣。这个系统通过学习促进特定恐惧症,当潜在的大脑基质变得长期敏化时,则促进广泛性焦虑障碍。这个系统旨在对抗捕食——一种防御伤害和过早死亡的机制(可能像愤怒(RAGE)一样,因此导致了历史上所谓的”战斗或逃跑”反应的混淆)。文明进步的一个方面是建立了针对此类事件的安全网(从警察部队、军队到医疗保健系统)。然而,现在随着某些国家对个性和自由主义独立性的推动,我们面临着越来越多的新型、非致命性认知掠夺性行为,这些行为撕裂了社会结构,侵蚀了支撑人们生活的安全基础。因此,恐惧(FEAR)再次上升,那些试图从中获利的人群也在增加。临床焦虑不仅在面临日益艰难日常生活不安全感的人群中大量存在,而且在那些经历过人类生活中最强烈创伤的人中也大量存在。不幸的是,多年来在我们的退伍军人医疗中心治疗创伤后应激障碍(PTSD)一直是一个增长的行业。我们对学习和记忆的神经机制的理解现在正在为直接药理学干预这些过程开辟道路,有可能加速心理治疗的进程。目前,心理治疗的效果可以通过认知促进剂如D-环丝氨酸来促进,它实际上加速了治疗性再学习,如暴露疗法。在过去十年中,已经进行了成功的人体试验,使用D-环丝氨酸来促进焦虑障碍(如创伤后应激障碍(PTSD))的心理治疗(Ganasen et al., 2010; Heresco-Levy et al., 2002; Hofmann et al., 2006)。动物模型的研究也表明,可以通过药理学方法抵消恐惧或创伤性记忆,使它们在心理上不那么令人困扰(Adamec & Young, 2000; Adamec et al., 2005)。
欲望(LUST)是年轻成年人和青春期少年常见的情感”伴侣”,它在人际关系中既可以是积极力量,也可以是消极力量。这个系统不受调节的唤醒可能参与各种反社会行为,从不受欢迎的性骚扰到对欲望对象的掠夺性追求和跟踪。伴随欲望(LUST)而来的持续寻求(SEEKING)行动冲动也应被认识到对支配关系和决定谁的意志在社会互动中占上风至关重要。值得注意的是,青春期和成年期社会-性行为的质量可能由早期社交游戏关系的质量塑造。挑战在于以优化健康和幸福结果的方式管理这些能量,而不是让它们成为生活挫折的主要来源。在撰写本章时,新闻报道了一位中年计算机程序员,他极度孤独,因社会-性失败而充满欲望的愤怒,在匹兹堡郊区的一家健身俱乐部谋杀了三名女性并射伤其他九人后自杀。在一次针对女性的精心策划因而是蓄意的暴力行为中,他发泄了二十年来无法吸引女性性伴侣的挫败感。也许这个迷失的灵魂小时候很少参与社交游戏,没有学会如何以有趣、友好和非威胁性的方式接近他人。欲望(LUST)和积极的寻求(SEEKING)的演化能为关爱(CARE)和游戏(PLAY)的演化提供一个初级加工平台,从这种不人道的行为来看很难相信。然而,每种原始情绪都有助于产生独特的高阶人格结构,这些结构并非情绪原型所固有的,凸显了学习和文化在每种情绪如何表现中的重要性。
关爱(CARE)是大自然母亲的伟大馈赠,有助于促进终生的韧性(resilience),并增加终生幸福的可能性。没有关爱(CARE),人类无法在人际关系中茁壮成长。当这种动机缺乏时,反社会和精神病态倾向可能会滋长。没有关爱(CARE),心理治疗几乎注定会失败。内源性阿片类物质和催产素(oxytocin)在所有社会情绪中都起着重要作用,最佳治疗环境可能需要募集这些神经化学物质。促进大脑中阿片类物质活性可以迅速缓解许多最严重的抑郁症形式(Bodkin等人,1995)。相当多的抑郁症可能是由大脑中愉悦化学物质减少引起的(Watt & Panksepp, 2009)。当高级脑心(BrainMind)区域被亲社会关切和观点塑造时,关爱(CARE)无疑在共情(empathy)的出现中占据重要地位。
悲伤(GRIEF)发出社会需求信号。那些得不到足够关爱的人将在当下和日后遭受更高的悲伤(GRIEF)和心理痛苦——他们的生活更可能充满分离痛苦,导致长期的不安全感、悲伤感和无法体验快乐。边缘性人格障碍和社交焦虑障碍的慢性不安全感可能源于这种过于常见的负面情感。悲伤(GRIEF)系统的过度活跃可以促进从抑郁症到社交恐惧症和惊恐发作等一系列其他精神问题;该系统的长期活跃不足可能促进适应不良的依恋风格以及自闭症和精神病态的冷漠。但最重要的是,这个系统促进悲伤和哀痛的感觉,这些感觉可能变成慢性心理痛苦(Panksepp, 2011a)。许多常见形式的抑郁症是由这些感觉以及随之而来的脑心(BrainMind)内绝望引发的神经化学级联反应所促发的(Watt & Panksepp, 2009)。如前所述,安全的阿片类药物如丁丙诺啡(buprenorphine)在治疗对常规抗抑郁药反应不佳的抑郁症方面利用不足。但有充分理由相信,这些药物只有在伴随改善的社会依恋,包括与心理治疗师的临时依恋(他们的任务包括促进对生活肯定性社会可能性的新的现实世界观点)时,才会产生最佳的长期治疗效果。
PLAY(玩耍)网络可能为我们提供了进化上最新的基本过程情绪冲动:创造性且愉快地参与他人心智世界的冲动,建立友谊并通过热切的友好竞争(伴随着反复的、充分平衡的互动——从某种意义上说,有输有赢)来学习。当这一过程在充分接纳的情况下进行时,往往伴随着欣快的积极情感,让人在社会秩序中感受到安全的归属感。令人遗憾的是,幼儿这种天性活泼的行为常常被父母和其他成年人视为问题,这可能导致在儿童冲动行为最令人不安的情况下,在生命的某个阶段被诊断为ADHD(注意力缺陷多动障碍),在另一个阶段被诊断为躁狂症。但PLAY对于社交技能的发展也是一种福祉。像所有基本情绪一样,PLAY是一个特别丰富的经验期待过程,它激发了大量的学习,最终与GRIEF(悲伤)、CARE(关爱)和LUST(欲望)等社会情绪一起,成为世界上许多艺术创作的驱动力。在最理想的状态下,PLAY充满了生命中最伟大的快乐之一:笑的能力,这是大自然最美好的情感馈赠之一。而且这种福祉并非人类独有。即使是老鼠在彼此玩耍或被人类好玩的手挠痒痒时,也会发出一种快乐的类似笑声的声音,这会产生可测量的大脑益处(Burgdorf等人,2010;Wöhr等人,2009;Yamamuro等人,2010)。基本过程的社交快乐可以通过敏锐的乐趣感和幽默感,被塑造成独特的治疗干预措施,这可能可以抵消诸如压抑的RAGE(愤怒)等慢性负面情感,而且通过卓越的临床技能,甚至可能平息深刻的FEAR(恐惧)和GRIEF的挫败性影响。也许玩耍性的快乐在心理治疗中被大大低估了,尤其是在年轻人中。确实,有充分的理由相信,它可以作为幼儿的日常心理补品,帮助减少我们文化中不断增长的ADHD诊断/发病率。我们对这一情绪系统日益增长的理解应该在讨论各种情绪问题(包括抑郁症)时发挥更重要的作用,因为促进PLAY冲动应该成为未来治疗干预的关键目标。
总之,这些系统需要在各种心理健康问题中加以考虑,这里只是简要提及,而对它们的详细神经科学理解可能对开发新的精神病学系统学和更加特定的脑-心智药物至关重要。它们都通过一个祖先的核心自我结构汇聚成一个协调一致的身体动作和情感可能性的交响乐,这一结构在所有哺乳动物中同源共享。在更高的脑区域,这种统一性可能会丧失,因为各种独特的第三过程自我被产生,为分离性身份(或多重人格)障碍提供了神经基础。
我们必须记住,在心智的基础上,所有这些系统都是能动的、行动网络。情绪困扰的人应该被允许在适当的时刻释放他们的能量,治疗师应该接受训练,以识别何时情绪是真实的,以及如何通过理解人类情感的深度,帮助将负面情感转化为正面情感。
虽然心理治疗显然必须继续现实地处理对个体人类生活的整体性、多层次的认识,但我们终于进入了一个时代,对心智-大脑的规律性(普遍性)部分的理解——基于证据的哺乳动物大脑基本过程情感观点——可以提供对所有人类共享的心智-大脑普遍情感-情绪基础的理解。反过来,这一知识应该为新的以情感为导向的疗法提供科学基础,这些疗法同时考虑身体、心智和大脑的互动动力学。未来几代心理治疗师将通过在所有这些层面发展技能和理解而受益良多。
本章的其余部分将深入探讨这些知识的含义,不仅涉及我们对积极情绪生活基础的理解,还涉及促进心理治疗康复和肯定生命的文化倡议。对积极情绪的理解是”美好生活”意义的核心(Sheldon等人,2011)。随着我们对大脑情感过程的神经化学编码了解得越来越多,必然会出现治疗过度愤怒、焦虑、抑郁、恐惧症和创伤,甚至可能是精神病性妄想的新想法(Panksepp & Harro, 2004; Watt & Panksepp, 2009)。我不声称拥有心理治疗专业知识,但我觉得有责任探讨深思熟虑的神经科学可以为治疗带来的各种可能性。我鼓励临床医生进一步发展这些想法。
近几十年来最引人注目且高度可重复的再发现之一,是早期童年经历对未来心理健康的非常强大的影响(Heim等人,2010)。这方面的研究发现太多,无法在此总结,但它们的范围从诸如早产和药物暴露等物理风险因素(例如,Johnson等人,2010;Stone等人,2010)到母婴和儿童关系的质量(Fearon等人,2010;Lahey等人,2008)。受虐待的儿童可能会发展出慢性愤怒和各种心身症状。但就像在老鼠身上一样,充足的母爱关注会促进韧性,并在儿童成长为成年人时产生更好调节的压力反应(Lester等人,2007;Propper等人,2008)。
关于是否可以仅通过处理当前生活环境来”治愈”此类患者,长期以来一直存在争议。有些人认为,更直接地处理过去的情感创伤(emotional traumas)是至关重要的。例如,Art Janov(2007)的观点是,有方法可以引导经历早期情感创伤的人回到其内隐创伤记忆(implicit traumatic memories)的源头。他认为,在适当的支持性和理解性治疗情境中,特殊的重新体验技术可以永久性地改善那些持久的大脑变化的影响。这将是一个新型ABT的例子,但它仍有待在标准化治疗试验中得到充分验证。
其他新策略的发展潜力巨大,包括心理-行为策略以及躯体和神经药理学联合干预。这些方法的发展可以部分地受到我们对古老的初级加工情绪系统(primary-process emotional systems)所学知识的指导。我现在将考虑这些知识如何对新疗法的发展产生有用的影响,以及对精神病学分类(psychiatric taxonomy)演变的影响,例如直接基于潜在内表型(endophenotypic)情绪问题的精神病综合征编码(Panksepp, 2006a)。
对动物大脑的研究使我们能够以神经科学细节聚焦于初级加工或核心情感问题——这在人类研究中是完全不可能的,即使使用现代脑成像技术(该技术更适合研究大多数心理学家感兴趣的高级认知区域)。动物研究使我们能够深入情感核心。跨物种临床前研究使我们能够对情绪疾病的本质和心理健康的情感必需条件形成越来越连贯的理解。目前,我们主要依赖试图根据一般症状(即DSM型”综合征”)对精神疾病进行分类的描述。让我们简要考虑一下精神病学主要手册中编码的这些现有诊断类别——美国精神病学协会在过去半个世纪中发布的《诊断与统计手册》的各个连续版本。当前的精神病学诊断基于由Eugen Bleuler、Sigmund Freud和Emil Kraepelin等先驱传承给我们的概念。
这些概念性精神病综合征从未基于对大脑或其情绪系统的理解。这就是为什么过时的概念从DSM-I到DSM-IV造成了越来越多问题的原因之一。许多人担心当前DSM-V的构建不会免受这些缺陷的影响(Hyman, 2007)。如果我们能用对心智中真实的与精神病学相关实体(例如,情绪大脑的各种内表型)的更好认识来替代(或至少补充)旧的概念结构,这些问题可能会得到缓解。
内表型是可以在神经科学水平上研究的大脑功能的自然方面;例如,研究人员可以检查像眨眼、瞳孔扩张或收缩以及惊吓反射这样简单的反应(Gottesman & Gould, 2003)。我们知道,巨大的噪音总是会使人和动物受到惊吓。然而,当第二次暴露于相同的噪音时,惊吓反应就不那么明显了。这种现象,当使用温和的听觉刺激来让动物为更大的声音做准备时进行研究时,被称为”前脉冲抑制(pre-pulse inhibition)“,在精神分裂症患者中往往减弱或缺失。我们可以确信,这些准备性、调节性反应是由大脑内连贯的、可分析的回路(特别是基底神经节)控制的,尽管我们对精神病学综合征不能有这样的信心,因为那些是由人类洞察力和创造力创建的概念。
通过情感神经科学,内表型思维(几乎被普遍认可为精神病学科学的一种重要新方法)可以包括初级加工情绪领域。大脑的自然情绪网络可能提供最相关的内表型,因为它们深入精神病学问题的情感核心。沿着这些方向的进一步发展可能有助于我们切开由以往时代曾经革命性的综合征思维所产生的戈尔迪之结(Gordian knot)(Panksepp, 2006a)。
当前的问题最好通过最普遍的精神疾病来说明,包括各种类型的抑郁症和其他情绪障碍。正如《生物精神病学教科书》引言中所指出的(Panksepp, 2004, 第18页):“DSM-II只有8种”(情绪障碍)类型,“但到了DSM-III(修订版)就有97种,根据Paul McHugh(2001)的说法,如果考虑DSM-IV中的所有子类别和详细说明,可以分类出2665种亚型。”这种复杂性源于MindBrain组织的更高层次,这些层次在个体之间可能存在巨大差异。放大诊断细节并不能为可靠的大脑研究或治疗实践提供任何清晰的联系,许多人已经开始渴望一种不同的方法来分类和处理精神问题。围绕情绪内表型(endophenotype)建立诊断分类的可能性只是其中之一,但目前是精神病学未来最可靠、最具科学依据的愿景。目前其他流行的方法包括基于遗传学基础的方法,但这些联系尚未产生太多清晰度——只有易感因素,以及许多意义不明的”联系”。无论如何,从业者应该清楚地设想哺乳动物大脑的自然原始情绪系统,并且至少部分地从这些角度来概念化人类情绪问题。
这可以导致新型的精神病学系统分类以及实验性心理治疗的新思路(已经提到了一些,稍后还会提到更多)。但要实现这样的进展,我们还必须开发具有明确精神病学相关性的临床前(动物)研究新方法,关注可以进行实证评估的原始情感的多样性。跨物种的进化视角在这些追求中将是非常宝贵的。例如,贯穿本书的一个整合性观点是,悲伤系统(GRIEF)和寻求系统(SEEKING)的失衡可能是抑郁症发生的主要影响因素(Watt & Panksepp, 2009; Panksepp & Watt, 2011)。悲伤系统促进了抑郁症特有的心理痛苦。当寻求能量减少时,就像在所有持续的负面情绪状态下似乎会发生的那样,就会出现慢性烦躁不安以及深度心理疲劳和空虚,这可能反映了对寻求冲动的主动抑制,或者只是这些充满活力的欣快资源的耗竭。
提出这些思路是因为它们允许对精神障碍的临床前建模采用全新的方法。例如,在抑郁症建模中,我们可能不再需要对动物施加大规模的负面压力源,例如常用的社会失败的持续压力、连续可变压力或重复的不可预测压力源。相反,我们目前已经了解到足以尝试诱导特定潜在情绪网络的失衡(Wright & Panksepp, 2011)。同样,我们可以更明智地投资于通过监测对系统性应用刺激(如挠痒程序)的情感性声音反应来直接测量内在情感状态,这些刺激探测相关积极情感系统的状态(例如,大鼠的快乐50-kHz超声波),同时还监测相关的负面情感反应(例如,痛苦的22-kHz”抱怨”),这些反应可以通过向大鼠颈后吹一口气来轻柔地诱导,而不是使用非常笼统和非特异性的抑郁情感测量方法,例如在强迫游泳任务中测量绝望(放弃),或者小鼠被尾巴悬挂时挣扎的减少等。真正令人惊讶的是,目前我们有大量的精神障碍动物模型,却没有对其大脑内的情感过程进行认真的讨论或评估。这无疑正在减缓我们对许多与人类精神病学关切直接相关的基本问题的理解进展。我们可以通过更认真地对待初级过程情感回路来做得更好。
基础情感神经科学、精神病学诊断和临床实践的连贯融合才刚刚开始。尽管对新的综合充满热情,但各种可能性之间的坚实桥梁仍有待建立。由于历史原因,传统的临床前研究仍然固守于行为模型,在这种模型中,可测量的行为和身体化学物质很重要,但情绪感受并不重要。
精神病学研究的行为方法将视觉上明确的症状(例如,强迫游泳测试中的持续不动)视为终点,即精神疾病的主要指标。国家精神卫生研究所前所长Steven Hyman(2007)特别批评用于模拟人类精神综合征的现有动物模型的不足。但他没有注意到,它们可能”不令人满意”主要是因为基础神经科学模型的研究人员很少使用情感概念来指导他们的思考。由于令人不安和失衡的情感体验是许多精神疾病的触发因素,因此更明确地认识产生情感的哺乳动物大脑系统应该是有帮助的。如果我们投入更多精力研究情感变化,那么我们的动物模型可能会为精神疾病的主要调节者和来源的大脑系统中正在发生的事情提供更好的见解。
对于更好的模型,我们的首要目标应该是通过使用神经行为学方法——自然情绪行为的神经学——来描述其他动物无条件的原始过程情绪性的大脑解剖结构、生理学和神经化学特征,然后利用这些知识来理解人类和动物心智的情绪失衡。通过直接操纵特定的情绪回路,人们有可能剖析对长期心理健康结果最重要的影响因素。这种理解的沟壑之所以没有被更积极地开拓,原因之一是动物的情绪感受被那些最有能力在这些至关重要的主题上做实质性工作的科学家们边缘化了。也许再次考虑相关的真理图表(图1.5)可能有助于阐明我们目前在思考其他哺乳动物的原始情感感受时应该处于什么位置。有压倒性的证据表明,所有其他哺乳动物都强烈地感受到原始过程的情感状态,但目前,在临床前模型中描述这些情感系统的工作相对较少。
在人类中,伴随情绪唤醒的情感充沛的思想在情绪唤醒消退后很快变得无意识并不罕见。换句话说,当一个人情绪强烈时在心智的高级区域搅动起来的反思性想法,一旦情感风暴过去,可以迅速变得认知上无意识,通常保持休眠状态,直到原始情感再次被唤起。一旦激情消退,就很难将与情绪唤醒相关的想法和意象流明确地带回现象经验中。这就是为什么在治疗环境中重建原始情感可以成为改变的快速途径。因为情感激发和引导认知装置,临床医生可以直接处理随之涌现的相关想法和沉思,使治疗师能够直接理解行动中的适应不良情绪模式,这为重塑它们对每个患者心智装置的影响力提供了理想时刻。
在这样可行的时刻,可以应用新的治疗干预措施,使临床医生能够通过我们对记忆”再巩固”的新兴理解来”软化”过去经历的痛苦边缘(见第六章)。当记忆被检索时,它们可以被修改、重新处理,并希望以不那么令人不安的形式再次存储。事实上,有理由相信,如果在情绪危机时刻能够唤起对抗性的积极情绪,那么麻烦记忆的长期影响可能会减少。
然而,在继续之前,让我们简要承认该领域遵循类似路径的一些历史线索。已经提到的阿瑟·贾诺夫的原始疗法(不是流行情景喜剧神话中的”原始尖叫疗法”),以及莱斯利·格林伯格及其同事更为保守但高度有效且以情绪为导向的过程-体验方法(见Elliott等,2004),还有哈比卜·达万卢(2005)和大卫·马兰(1979,1999)等人的短期体验动力疗法,其主要目标是让人们足够强烈地体验他们的”真实感受”,以实现持续的修正。
越来越清楚的是,动态情绪聚焦方法在促进持久治疗改变方面通常非常有效(Abbass等,2006),通常比对高级心智功能的认知行为重新解释和重构产生更持久的结果。情感体验方法通常基于这样的观点:麻烦的情绪可以比认知激发相关感受更快地激发相关思想,这可以有益地使用,至少在支持性治疗环境中。情绪事件的强烈再体验开辟了新的治疗可能性,因为它为治疗师提供了情感上的”亲密”,特别是在安全的治疗联盟中,这对治疗改变是最佳的。通过用情感积极的视角重新框架,可以减少情感上麻烦的记忆的影响。
以情绪为导向的疗法似乎非常有效,因为它们立即处理相关的原始情感——从而从心智的高级区域带来最相关的联想认知材料。临床医生听到这样的话并不罕见:“我母亲总是对我要求太多,对待我好像我永远不够好”;“我比其他孩子小,我今天仍然感到被贬低和不安全”;“我一生都在与此抗争,我仍然无法原谅自己。”从这些临床故事线中可能出现的人格形象很容易辨认:怨恨地履行义务的孝顺孩子;忙碌、自我怀疑的成就过度者;悲伤、沮丧、自我破坏的无用之人。当然,这些原型故事线几乎与自己的人生故事一样熟悉。它们反映的情感对长期情绪健康往往是适得其反的,因为它们触及人们心智生活的失衡情感基础。但这正是治疗工作需要应用的地方,希望能够修改那些使消极存在状态成为习惯的持续且情感强大的学习模式。
幸运的是,情感问题目前正处于科学化治疗思维的最前沿。当前,心理治疗中一些最有趣的讨论正从新的跨学科前沿领域涌现:(i)发展社会神经科学(Schore, 2003a, 2003b; Siegel, 2010; Stern, 2004);(ii)新兴的神经精神分析学(neuropsychoanalysis)(Solms & Turnbull, 2002);(iii)人类和跨物种情感神经科学(Davidson, 2004; Panksepp, 1998a);以及(iv)关于自主神经系统调节过程的前瞻性视角(Porges, 2009a)。这些方法最终开始认真应对人类心智的情感本质,有时也涉及哺乳动物大脑的深层情感本质。
Dan Siegel在Louis Cozolino(2002)对人类心智的临床和神经科学方法的综合研究前言中说得很好:他说,临床医生”沉浸在前来寻求帮助以改善感受的个体的故事中……无论采用何种方法,治疗中持久的改变都是人类心智改变的结果……这涉及大脑功能的变化。心智在治疗过程中究竟如何改变,这是神经科学与心理治疗综合研究所要解决的根本难题。“也许这个难题中最关键的神经科学部分是情感感受如何从大脑中产生,以及慢性情感感受如何改变。提供对这一知识的介绍一直是本书的目标。
心理治疗师对情感的神经本质、它们在行为中的体现以及它们如何与认知过程互动的兴趣日益增长。人们热切关注如何更好地利用情感状态来重塑困境中人们的情感幸福感(例如,Fosha等,2009a)。尽管心理治疗传统上更多地寻求处理个体生活情感迷宫的认知方面,但少数革新者正在将讨论转向关键的情感问题(例如,Fosha, 2000; Greenberg, 2002; Greenberg & Watson, 2005; Hughes, 2006, 2007; Ogden等,2006; Schore, 1994, 2003a, 2003b; Siegel, 2007, 2010; Stern, 2004)。
早期社会依恋不安全的作用是脱轨情感生活中一个特别突出的因素(Heim等,2010)。因此,一些革命性的治疗师渴望找回已铭刻在发育中的神经矩阵中的早期内隐情感”记忆”,这些记忆控制着婴儿的原始心智状态,并通过使用非语言形式的”原始疗愈”直接处理它们,以修复延续到成年期的残留心理”创伤”(Janov, 2007)。这些直接处理最早期童年创伤的尝试——反映在敏化和脱敏的情感系统中——正在寻求从大脑的记忆库中消除强烈体验情感的内隐残留。许多患者在被鼓励重新体验这些早期创伤时报告了显著的益处。我们需要考虑如何将这些治疗模型与我们对哺乳动物大脑古老、普遍的情感原则的新兴理解相结合。这里没有明确的答案,但在治疗性关爱(CARE),甚至是玩耍(PLAY)的背景下,过去创伤性经历的”再巩固”这一想法再次浮现在脑海中。
因为我们现在可以掌握初级过程情感的神经动力学,我们可以设想目前处于地平线上的新型情感基础疗法(ABT)变体。我们或许甚至应该考虑长期被忽视的想法,比如简单地尝试用积极情感的疗愈力量来对抗消极情感。可能的例子包括不仅旨在直接快速触及个体情感生活的情感导向疗法,还利用各种身体疗法,使用身体-大脑-身体连续体的特性来快速将情绪转向积极情感。每种情感都有这样的联系。当患者的情感行动装置已经僵硬地”冻结”在消极情感状态时,治疗师最初鼓励那些允许心智、大脑和身体转换到不同情感状态的动作和身体重新定位类型,难道不明智吗?希望转向更灵活的积极感受,在那里可以考虑不同的情感视角。例如,玩耍式互动,以及直接的身体工作,有时是否会比仅停留在认知层面的互动产生更快的进展(Ogden等,2006)?我们是否需要考虑脑心(BrainMind)组织的所有层面,从初级到三级,以实现最佳治疗进展?如果进化上更古老的情感过程指导着高级心智装置如何运作([图2.3]),怎么可能不是这样呢?我们需要同时思考身体和大脑、身体和心智,以最有效地处理情感问题。
如今,研究人员和心理健康专业人员对理解支撑精神病理学的神经情感失衡和障碍,以及心理治疗带来的心智大脑(MindBrain)变化越来越感兴趣;这些过程可以通过当代脑成像和其他神经科学技术来呈现。由于这些问题在其他地方已有充分讨论(例如,Cozolino, 2002, 2010; Doidge, 2007),我在本章的主要目标是关注我们对情感感受如何在大脑中产生以及记忆如何巩固以促进临床干预进一步发展的新兴理解。
尽管我们终于进入了一个时代,大多数深思熟虑的研究者都认同弗洛伊德关于心理的生物学和情感基础的信念(Freud, 1937/1968, p. 357),但本书中设想的那种相当直接的情感性脑心(BrainMind)动力学仍有待广泛整合到治疗思维中。这部分是由于历史障碍,其中一些已经讨论过。但还有许多其他障碍。事实上,弗洛伊德自己的精神分析元心理学,也许被过多的概念包袱”丰富”了(例如,俄狄浦斯情结、阴茎嫉妒),可以作为一个例子。它是从有限的、受文化束缚的临床观察中创造性地构建出来的,导致了不太有利的历史轨迹。多年来,许多问题源于理论创造力的过度,伴随着对大脑和心智的进化层次缺乏扎实的理解。
由此产生的戈尔迪之结(Gordian knot)无法完全解开,但我们可以思考情感神经科学知识如何作为未来临床思维的新的、有望坚实的基础。主要教训是,不同个体之间的情绪在初级过程层面最为相似;它们通过学习和记忆在个体中变得多样化。而在三级过程层面,它们的差异将最大。精准的药物治疗可能在初级层面最有用,特别是因为这也可以对所有更高层面产生强大影响。行为治疗方法将在二级过程层面发挥最佳效果,而认知方法可能在思想和沉思层面非常有效,有效的程序具有一直向下的有益调节作用。新的动态情感平衡方法可以有效地处理整个包裹。
尽管哺乳动物大脑的初级过程情绪网络的唤醒被人类和其他动物强烈体验,但特别重要的是要认识到,脑心(BrainMind)的二级过程,即学习、记忆和习惯形成的基本形式,是最无意识的”心理”过程。一旦我们理解了这一点,那么心理学过去的许多怪异和错误观点可能会得到纠正。例如,“自由意志”并不是我们想象的虚构,尽管如今太多科学家准备这样宣称。自由意志是一种更高的三级神经认知功能,我们经常使用它(当我们情绪不太激动时非常有效)来规划未来的行动。这在Ryan和Deci(2006)发展的”自主性”和”自我决定”概念中得到了很好的体现。然而,我们无法轻易地将自己从通过在脑心(MindBrain)组织的初级和二级层面巩固不适应性情感模式而产生的潜在情绪混乱中解脱出来。在情绪处理的初级过程层面,没有自由意志,没有”受控认知”。由我们野性动物激情在发展过程中塑造的自动二级过程学习和记忆功能也不表现出自由意志。这只能从精心雕琢的、深刻的自我反思性的认知态度中出现。
我们原始的情感需求和身体动机在我们知道之前——在我们”意识到”之前——塑造了我们成为什么样的人,往往在认知上产生没有我们”个人”同意的最终结果。因此,重要的是要认识到,我们对情绪的原始的、情感性的现象体验和我们对情绪的认知反思性意识是非常不同类型的心理过程。对于快速的治疗改变,也许通常情感体验本身需要成为起点。但情感体验一直是神经科学的最大问题之一,很少被谈论,因此抵制实证理解……事实上,甚至抵制进行有力的科学分析的尝试。本书的主要目标之一是介绍如何改变这种状况。
为什么对情感产生机制的详细神经科学理解出现得如此缓慢?部分原因是它只能从那种在人类中完全不可能进行但在动物模型中越来越可行的详细大脑研究中产生。由于传统上反对在分析动物和人类行为的神经控制的科学分析中使用初级过程心理构念的保守偏见,进展进一步延迟。因此,基本情绪网络及其产生的情感性感受根本没有得到应有的关注。而且这些感受通常继续被那些最适合揭示其神经基础设施的人(即行为神经科学家)所忽视,因此在现代生物精神病学讨论中不像它们需要的那样明显。
从某种意义上说,大多数对学习和记忆感兴趣的研究者如此有效地使用经典恐惧条件反射作为主要方法(见[第5章]),却尚未明确承认他们研究的动物大脑中存在无条件恐惧系统,这是令人遗憾的(Panksepp et al., 2011)。杏仁核产生情绪行为和相关的自主反应,但它们通常被描述为动物的无意识运动”输出”(Davis et al., 1995, 2010; LeDoux, 1996),而不是产生情感的情绪系统。这种短视阻止了那些对恐惧学习感兴趣的人设想”无条件恐惧反应”——即恐惧系统的唤起——可能对恐惧学习的产生至关重要(见[第6章])。一旦我们意识到这个回路也是控制焦虑感受的位置,我们就能设想记忆如何变得令人恐惧。更重要的是,我们可能开始积极考虑各种积极情感回路如何抵消这种心理消极性,有望产生让麻烦记忆在更可接受的情感框架中重新巩固的方法。
为了实现新的治疗进展,我们需要理解无条件恐惧系统的持续唤起如何对慢性焦虑障碍的产生起到关键作用(Panksepp, 1990b; Panksepp et al., 2011)。新的焦虑治疗应该旨在抑制这个系统的心理影响,无论是通过药物脱敏恐惧系统,还是通过心理治疗来化解可怕的记忆。目前可以通过直接药物降低恐惧系统的可唤起性来实现这一点,例如使用苯二氮卓类药物,以及在较小程度上使用唤醒(如大脑去甲肾上腺素)抑制剂——“β受体阻滞剂”如普萘洛尔。这些药物可以减少通常唤起这个系统的麻烦记忆的影响,并可能让心理治疗增强对抗性积极情感系统的”力量”。在考虑这些选择时,始终重要的是牢记脑心智进化中的控制层次:恐惧系统促进充满焦虑的记忆,导致令人不安的想法,这些想法因人而异。然而,源于原始恐惧系统的习得性焦虑在人类和其他哺乳动物中必然非常相似。
由于遗传学和神经科学的进步,现在很清楚动物模型可以促进对许多仍然控制人类生活的古老情感原则的准确考古。因此,跨物种情感神经科学帮助阐明了许多主观体验的原始情绪感受,这些感受是进化提供的鞭子和胡萝卜——我们无穷复杂的认知能力的情感纽带和指引。最终,许多动物和人类的学习都与某些行动方式如何让有机体感受密切相关。因此,损害心理健康的消极情感纽带需要用能促进幸福的积极情感指引来对抗:这些可以包括极富想象力和创造性的思维(由我们的寻求系统促进)到伦理和道德决策(由我们所有的亲社会情绪——关爱、悲伤和玩耍促进)。如果我们理解这些感受的神经生物学本质,以及它们如何控制学习,我们可能就有了关于人类心智体验积极情绪意味着什么的坚实神经科学基础的开端,从而通过明确的、情感上有益的临床干预来更好地对抗情绪障碍。现代脑成像将有助于这些努力(例如,概述见Cozolino, 2010; Northoff, 2011)。但同时,我们应该对这些技术的解剖学和功能限制持现实态度。
在我们当前的脑成像时代,情绪脑的古老区域受到的关注少于新皮质功能,部分原因是这些技术对大型、高放电神经系统更敏感。结果是专注于情绪过程的认知调节。相对较小、缓慢放电的神经元脑区(其中释放的化学物质比动作电位的频率更重要)不容易被这些技术可视化。然而,这些神经生理学上”迟缓”的低级脑区对我们的情绪生活具有决定性重要性。
正如我之前指出的,基本情绪系统的皮质下定位已经被以下事实戏剧性地证实:研究者可以在各种”简单”实验动物出生时手术切除所有新皮质,就基本情绪能量而言,这些受试者成长为看似正常的生物。它们表现出探索冲动和寻求行为、恐惧、愤怒、性欲、母性关爱和玩耍。最后一点尤其令人惊讶,因为身体玩耍是一种动态灵活的行为。在没有新皮质出生的人类儿童中也观察到了类似的模式(Shewmon et al., 1999; 见[图13.2])。
当成年人遭受类似的脑损伤时,功能障碍会严重得多,这可能主要是因为一旦原始冲动在成熟的新皮质区域内被认知性地重新表征(cognitively rerepresented),人类和其他动物就会越来越依赖那些更高级的、发育程序化的”软件”功能。一旦开始依赖这些精良的新皮质认知工具来实现更高形式的意识,就无法有效地回到更简单的存在方式。新皮质是否具有任何基于进化的情感功能,而非依赖学习的发育,目前尚未解决。它确实可以产生大量的情绪性思维和行为。然而,情绪性情感(emotional affects)的中心似乎仍然位于皮质下,尽管古老的皮质区域如岛叶(insula)可以产生各种特定的感觉性情感感受,如厌恶和疼痛(Craig, 2002, 2009),但肯定离不开皮质下回路的参与。眶额叶区域(Orbitofrontal areas)参与许多与味觉、温度和其他感觉奖赏和惩罚相关的负性和正性感受。
当人脑的皮质下情绪力量变得狂暴(或失调到难以理解的程度)时,可能会出现压倒性的、往往持久的情绪问题。在人类中,这些问题总是伴随着认知变化,如情绪纠缠的归因(attributions)、反刍(ruminations)、各种计划和担忧,以及关于世界如何组织的认知”命题态度(propositional attitudes)“。这一事实有助于解释为什么每次在完整的心脑(MindBrain)中发生情绪时,环境中似乎总是存在引发性的认知原因,以及对我们思考和感知世界方式的认知后果。但情感变化是大多数精神疾病的基础。
这种观点表明,心理治疗不仅需要处理情绪动荡的认知诱因,而且需要更直接地处理并发的情感问题。情感神经科学表明,有些人会在没有诱发事件的情况下变得过度情绪化,这是因为内部脑刺激。一个例子是当个体因敏感化的情绪网络而出现”边缘性癫痫发作(limbic seizures)“时(Lewis & Pincus, 1989)。严重的情感失衡可能纯粹出于神经生物学原因而发生。这类问题可以通过直接操纵大脑来缓解。认知干预是不必要的,尽管明智的咨询总是有用的,特别是关于弗洛伊德称为”修通(working through)“的重新调整过程。
还有一些童年创伤主要在情绪系统的反应性上留下印记,而没有明确的认知残留(Janov, 2007)。皮质下回路可以通过经验而敏感化和脱敏化。这些问题也可能通过有针对性的药物干预得到帮助,也许不需要花费大量时间谈论自己的生活,尽管关注由此产生的人格特质如何影响一个人的生活应该能提供有用的见解。重要的是要考虑到,已经建立了强大治疗联盟(therapeutic alliance)的治疗师有能力通过在更原始的层面与患者互动,引导患者进入不同的、更积极的情绪状态。例如,当允许表达负面情绪,但熟练的治疗师也能唤起积极情绪时,可能可以明确地逐步引导患者走向更积极的情绪状态,使持久的治疗性变化得以扎根。
显然,慢性情绪压力会改变个体的反应方式和适应世界的方式。随着针对各种疾病的众多新药的出现,精神药理学革命为那些处于相对轻度但具有精神病学意义的痛苦中的人提供了许多例子,说明简单地操纵脑化学物质可以产生巨大的治疗效果(Kramer, 2005)。事实上,大多数精神药物都是ABTs(情感基础疗法),因为当它们效果良好时,它们严格在非认知神经化学水平上以理想的方向改变初级过程的情绪反应性和情绪。但这种不良感受的减少,往往使认知的观点采择(perspective-taking)变得更加有效。
显然,大多数人类情绪问题是由生活事件引起的。有一个关心的人简单地倾听情绪事件的全部影响,可以说是直接从战场上听取(Belenky, et al., 1996),这本身就是治疗性的。众所周知,当人们将感受用言语表达出来时,情绪的主观强度会降低(Lieberman, et al., 2011)。即使是由明确的非认知创伤引起的慢性情感问题,也会迅速嵌入复杂的认知叙事中,需要在治疗性对话中充分交流和探索。正如我们所看到的,基本情感和认知总是形成一条双向街道:情绪唤起改变我们的思考方式,而我们的思考方式可以改变我们的感受。许多表征人际关系和悲剧的”日常疯狂”需要在情绪和认知两个层面上处理。然而,认知促进的情绪困境的尖锐边缘可以通过在社会支持背景下进行”存在性见证(existential testimony)“的机会来软化,这可以促进正念(mindfulness),即以超越烦恼的平静专注于日常生活的能力(Siegel, 2007, 2010)。而在恰当的时刻明智地运用游戏性(playfulness),也应该有所帮助。
实际上,情绪和认知在完整的人类心智大脑中如此紧密地协同工作,以至于大多数心理学家不愿意区分两者,这种观点在自下而上的神经科学严格审查下站不住脚。初级过程情绪通过学习变得认知化——与内部和外部事件的特定意识表征交织在一起。因此,在大多数人类心理问题中,认知与初级情绪纠缠在一起,以至于它们无法轻易区分。尽管如此,情感神经科学强调了初级过程、前命题情绪能量状态如何拥有自己的心智,作为古老的情感心理活动形式,在漫长的进化时间中先于语言和思维出现。清楚地认识到这些情绪能量及其在精神生活中的作用,使人能够同时在更充分整合的情感和认知层面上进行治疗工作。
理解初级过程情绪如何使人类生活脱轨,对于科学理解所有类型的心理治疗以及建立新的基础神经科学基础设施来服务于生物精神病学的未来发展具有价值(Panksepp, 2006a, 2006b; Panksepp & Harro, 2004)。因此,情感神经科学提出了新的心理治疗视角,可以很好地补充已建立的行为、认知、人本主义、人际关系和正念治疗传统。
心理治疗可以从我们对大脑的情绪-本能行动动力学日益增长的理解中受益。简单地让人们理解他们大脑中这些普遍的、共享的系统本身可能就具有治疗作用。通过这种方式,情绪困扰的个体可能会以更多的信心和对情绪状态产生和调节背后的普遍原则的更好理解来面对世界。跨物种情感神经科学不仅为思考基本人类问题提供了连贯的结构,而且还提供了关于情感如何从大脑中产生的具体视野。无条件情绪动力学提供了一种科学方法来理解初级过程情绪感受实际上是如何在大脑中产生的。
情感平衡疗法可能提供基于证据的新方法来直接修改情绪感受,使临床医生能够使用新的心理治疗促进的情感态度作为重构认知扭曲和改善由此产生的内心压力的基础。实际上,这种基于情感的互动可能会增强经典药物疗法的有效性,这些疗法在上世纪中叶彻底改变了精神病学,最近又出现了各种直接脑刺激程序,从电休克和其他形式的脑刺激如经颅磁刺激和深部脑刺激(Panksepp, 2004)。其中一些方法能够直接修改神经系统的情感基调,因为它们作用于大脑皮层下区域的初级过程情绪网络(Coenen, et al., 2011)。
虽然初级过程情绪动力学来自我们与其他动物共享的皮层下大脑网络,但它们可能在心理治疗环境中比目前的典型做法更有效地被调用。例如,心理科学中早已知道,人们可以通过模拟情绪行为来诱导情绪感受(Stepper & Strack, 1993)。实际上,人们可以仅通过模拟笑和哭的行动动力学来快速获得情绪典型的情感变化,如喜悦和悲伤;这甚至可以通过心理行动意象来实现(Panksepp & Gordon, 2003)。同样,音乐是一种强大的诱导情绪的方式,可以用于治疗目的(Bernatzky, et al., 2011)。
我们如何对情绪表达以及情感状态进行自愿控制,以及如何在心理治疗情境中加以利用,仍有待系统研究。将这些特定情感的能量应用于各种体验式心理治疗似乎相当直接。高度集中的情绪练习的整合可以极大地促进针对人类问题的心理治疗方法(Ogden et al., 2006),同时也为教育人们了解他们情绪生活的初级过程方面提供机会。
以这种方式逐渐掌握自己的情绪动力学可能有助于为各种情况下的情绪智力(emotional intelligence)和稳态铺平道路。每天进行积极的情绪练习,可能会以能够抵消过去创伤影响的方式增强一个人的”情绪肌肉”,并使情绪回路免受未来逆境的影响。例如,当在治疗环境中唤起负面情绪时,可以定期跟随各种积极情感——从情绪强烈的音乐片段到富含积极情感的身体表达性动作。需要进行大量的基础科学研究来评估这些新技术的有效性,无论是在现有身体疗法的背景下(Ogden et al., 2006),还是作为情绪教育计划的一部分,该计划可能有助于为可能升级为重大心理问题的极端情绪提供预防。理解自己的情绪动力学,作为综合治疗计划的一部分,可能有助于减少压力引起的心理问题的发生率。
将情感原则的完整光谱融入心理治疗实践可能有助于重新定位行为主义在心理治疗中的遗产。行为主义为心理治疗提供了一颗珍贵的宝石:基于外部强化contingencies(contingencies)重新安排的行为矫正。例如,可以通过付钱让人们避免坏习惯来轻易减少不良行为,这一程序目前常用于治疗成瘾冲动。然而,这种外在主义观点继续扭曲心理学领域的认知思维,导致持续将生物体误解为被动的信息处理机器,而非情感上主动的生物。相比之下,对哺乳动物心智的情感机制的连贯理解提供了脑心(BrainMind)基础设施的清晰图景——即主动的、情感调谐的人际心理装置——这是治疗思维所需的指南。
然而,目前在临床思维中,精神病理学的认知概念仍然比明确的情感概念更突出。尽管情绪调节问题无疑与心智的认知和情感方面的失调都有关联,但也许我们当前的时代精神鼓励心理治疗师寻求对我们认知本质而非情感本质更全面的科学理解。这可能是因为在认知神经科学而非情感神经科学方面投入了更大规模的机构资源。无论原因是什么,我们对高级脑功能如何编织我们的认知的理解,并不比我们对低级脑如何产生情绪的理解多多少。我们确实知道,治疗师的人格特征——无疑尤其是他们情感调谐的能力——通常比他们使用的具体程序更重要。众所周知,当一个人感觉不好时,他人的关怀可以迅速减少负面情绪。十二步计划之所以如此显著有效,可能是因为它们提供了重新连接到积极感受潜能所需的社会关怀和肯定。他人心智的社会-情感力量可以帮助人们有效处理负面情绪,因此大脑的情感地形可能为大多数人类心理问题核心的心理力量,以及需要调动以获得最佳治疗效果的内在和人际心理动力学提供更清晰的描述。
确实,与心理治疗相关的认知问题可能比潜在的情感问题在科学上更难以把握、更难理解。当然,纯认知收益往往比情感收益更容易消失。人们在心理治疗过程中很容易达到表面清晰的时刻,但随着他们在预约之间回归旧的情感习惯,所有进展都会消散。这可能是因为每个初级过程情绪都为自私的目的”奴役”了大量认知领域。如果是这样,那么更直接地尝试实现情绪稳态可能比严格在认知层面工作提供更简单、更有效的途径来促进期望的认知重新定向。
自我中心情绪系统(关于这一概念的扩展,见Northoff & Panksepp, 2008; Panksepp, 1998a; Panksepp & Northoff, 2009)对认知过程施加的控制可能极其强大。例如,PTSD可以反映出由简单的次级过程记忆(例如,[第6章]中描述的经典条件反射)引发的高度厌恶情绪,这些记忆通常是无意识触发的(通过未注意的刺激)。令许多治疗师惊讶的是,最近发现相当简单的认知型干预——例如本章末尾将描述的眼动疗法——不追求任何深层认知”洞察”,却可以像其他治疗方式一样有效地抑制创伤记忆的力量。此外,也许PTSD的情感风暴可以通过知道如何帮助患者在情感积极体验中重新框定其创伤感受的治疗师而大幅减弱,这些体验可以通过非认知方法轻易触发。重申一下,现在人们普遍认识到记忆并不像大多数人过去认为的那样稳定。如[第6章]所讨论的,每次检索记忆时,都有机会帮助它们以不那么麻烦的形式”重新巩固”。目前,这一现象使人看到希望,即情感上令人困扰的记忆可以在积极的情感框架中重新配置,借助相当简单的身体操作,甚至可能系统地呈现积极的情感体验,如聆听舒缓、安慰、快乐的音乐。
由于所有心理治疗都必须从认知层面开始,而且大多数心理治疗旨在重构人们思考问题的方式,因此更直接的情感方法可能没有得到应有的广泛考虑。可以理解的是,在像人类这样语言几乎调节每一次互动的物种中,认知方法将在心理治疗事业中保持主导地位。但我们是否知道,即使有来自脑成像的充分证据显示高级脑区发生变化,主要的治疗效果实际上是在脑-心智的高级认知动力学中产生的?没有人真正知道,但也许许多有益的转变实际上是在大脑中所有嵌套的、分层的情感-认知交互层面上隐式发生的([图2.3])。事实上,如果治疗路径是通过改变初级加工情感基调铺平的,也许最持久的效果就会出现。如果是这样,临床从业者的工作可能会通过更充分地吸收和利用来自情感神经科学的关于大脑情绪系统的现有证据,并致力于更充分地利用最直接的情感手段而得到促进。虽然人们喜欢谈论构成他们外显心智的无尽情节记忆,而精神分析也以这种方式很好地服务于个人成长,但这种人际体验的认知方面还是情感方面在提供长期心理缓解方面更重要,这绝不清楚。毫无疑问,两者都很重要,但我预计,如果没有持续的情感变化,认知重构可能不会那么有效。
无论如何,很明显心理治疗正处于情绪革命之中。心智的原始情感方面不再被边缘化,而是被认为是心理的真正引擎(Fosha等,2009a,2009b)。例如,Greenberg及其同事在过程-体验疗法中的工作强调了在治疗情境中体验和表达清晰分化的初级情绪感受(如愤怒和恐惧)的必要性(Elliot等,2004;Greenberg,2002)。Foa及其同事(1998)的研究表明,在治疗过程中实际体验恐惧对于焦虑障碍的暴露治疗成功至关重要。相比之下,在处理战争创伤时,该领域的从业者发现,立即沟通和讨论所发生的事情,会为立即和明显的益处打开大门(Belenky等,1996)。但关键是,使用这些新方法取得进一步进展必须建立在理解潜在致病因素本质的基础上。这里没有足够的空间详细讨论所有主要精神障碍,所以我选择了抑郁症作为情感神经科学思维如何有助于进一步进展的范例。
那么大脑中抑郁症的来源是什么?这是目前神经科学探究的热门话题,而且抑郁症的持久答案必须部分来自对人们在经历生活起伏时所体验到的情感风暴的更好理解。当某些初级加工情绪系统变得长期失衡时,可能会出现抑郁症。如果是这样,我们需要从神经科学角度更好地理解初级加工情绪系统——存在于所有哺乳动物大脑中的原始情感内表型(endophenotype)(Panksepp,2006a)。每个基本情绪系统都可以通过重复的情感体验而变得敏感化或脱敏。每次情感体验在与我们对内部和外部现实的表征交互时,都可以促进各种形式的内隐和外显学习。通过这种对慢性感受-思维模式的塑造,人们的态度会变得僵化和消极,削弱更流畅的积极推理。
因此,感受可以以各种持续的方式变得极端,原因也有很多。毫无疑问,关于这些过程,我们需要知道的大部分内容仍有待发现。但抑郁症作为精神障碍的”常见病”,是需要关注的一个关键问题,特别是因为它在现代社会中的患病率很高且似乎不断增加。当然,这种增加可能只是表面现象,部分是由制药公司推动的,它们试图通过复杂的营销策略为其高利润、往往效果有限的抗抑郁药物创造需求。当然,历史上许多人在生活中都经历过短暂的抑郁发作,但新情况是,目前多达20%的人口寻求这些症状的医疗援助。而且,药物通常不是按需提供,而是长期提供。现在看来,持续使用抗抑郁药物很可能会在大脑神经化学模式中产生持续性转变(例如,通过受体超敏反应的发展),当停止用药时,这些转变会导致更强烈的负面情绪(关于精彩概述,请参见Marcia Angell在《纽约书评》中的分析:http://www.nybooks.com/articles/archives/2011/jun/23/epidemic-mental-illness-why/)。
但我们对抑郁的脑心(BrainMind)了解还不够深入,无法得出任何确定性结论。正如患者安德鲁·所罗门(Andrew Solomon)(2001, p. 29)所说:“我们必须坦白承认:我们并不真正知道是什么导致了抑郁。我们并不真正知道什么构成了抑郁。我们并不真正知道为什么某些治疗方法可能对抑郁有效。我们不知道抑郁是如何在进化过程中保留下来的。我们不知道为什么同样的情况会让一个人抑郁,却不会困扰另一个人。”我们无法理解抑郁,部分原因可能是神经科学尚未对哺乳动物脑心(MindBrain)中最相关的古老情感回路进行足够深入的研究。
事实上,到目前为止的讨论还没有概念化哪些类型的大脑情绪系统受抑郁影响最大。为了帮助解决这个问题,瓦特(Watt)和我(2009)最近提供了一个基于情感-社会神经科学观点的综合性论述,包括同行评论和扩展回应,这可能会促进这一顽固问题的进展。我们的提议是,要理解抑郁症,需要理解由持续分离痛苦引起的精神痛苦——悲伤系统(GRIEF system)的过度和持续激活——这是大脑中最重要的社会情绪系统之一。此外,可能由于悲伤系统的持续激活,寻求系统(SEEKING system)的唤醒能力在抑郁中会减弱(如前所述,另见Coenen等人,2011;以及Panksepp & Watt, 2011;和Zellner等人,2011)。
我们已经看到,悲伤系统——集中在前扣带回(anterior cingulate gyrus)、腹侧隔核(ventral septal nuclei)、背侧视前区(dorsal preoptic area)以及终纹床核(bed nucleus of the stria terminalis, BNST)、背内侧丘脑(dorsomedial thalamus)和导水管周围灰质(periaqueductal gray, PAG)——在产生悲伤和哀痛情绪以及哭泣冲动方面起着重要作用(Herman & Panksepp, 1981; Panksepp等人,1988)。加剧痛苦感受的脑化学物质(例如促肾上腺皮质激素释放因子的释放)和那些强有力缓解痛苦的化学物质(例如脑阿片类物质、催产素和催乳素),是社会依恋形成的重要因素,并且都可能在社会联结的调节(Nelson and Panksepp, 1998)以及抑郁情感中发挥作用。海伦·梅伯格(Helen Mayberg)及其同事(2005)通过对膝下前扣带(subgenual anterior cingulate)进行深部脑刺激,为难治性抑郁症提供了显著缓解,在那里可以抑制介导悲伤型沉思的高级脑区域。这种脑刺激很可能正在扰乱由悲伤系统产生的精神痛苦的神经来源。事实上,这个系统的慢性过度激活可能是抑郁性烦躁的主要来源之一。
寻求系统的全局性关闭,这是从”抗议”到”绝望”转变的特征,可能是持续抑郁中的一个关键脑心变化。已经充分证实,早期的分离和失落经历会使人容易患抑郁症,或者可以触发第一次抑郁发作(Heim & Nemeroff, 1999)。此外,介导依恋和分离的机制在女性中更为敏感,女性患抑郁症的可能性是男性的两倍。我们也早就知道,调节大脑分离/依恋机制的情感性阳性阿片类物质具有强大的抗抑郁特性。如果不是因为阿片类药物的成瘾风险,它们可能仍会像20世纪50年代现代精神药理学出现之前那样被用作抗抑郁药。因此,抑郁可能部分反映了那些让我们在安全可靠地依恋他人时感到良好的天然大脑化学物质活性的降低(见第9章)。部分而言,抑郁可能反映了我们天然的内源性阿片类物质未能提供足够的安全感——简而言之,充满爱的社会依恋纽带是一种原始的成瘾形式(Panksepp, 1981a)。
如前所述,一种安全、非成瘾性的抗抑郁药物(混合阿片受体激动剂/拮抗剂——丁丙诺啡(buprenorphine))目前已经可用(Bodkin等人,1995),尽管还需要进行双盲、安慰剂对照试验来测试其疗效。它仍然是一种”安全的阿片类物质”,在低剂量下,丁丙诺啡可以直接对抗精神痛苦。它不会像大多数阿片类物质那样严重成瘾,因为随着剂量增加,该药物会发挥阿片类拮抗作用。可以预期,这类药物在减少自杀意念方面也可能相当有效。对悲伤系统的进一步研究应该会产生各种新药物,用于从抑郁症到社交恐惧症等各种疾病。然而,只有当我们现实地考虑潜在情感过程的本质时,这种转化研究才能有效地应用于临床问题。
尽管这些阿片驱动的依恋系统可能在抑郁中起关键作用,但可能还有许多相关机制介导各种抑郁亚型。例如,强啡肽促进的多巴胺驱动的欲求系统的关闭(当个体在绝望中精神上”放弃”时)可能在某些病例中形成独立的病因机制(Knoll & Carlezon, 2010)。另一种类型的抑郁可能源于与物质损失相关的情绪——尤其是在生物体竞争资源时,在支配性遭遇中产生的失败感(Panksepp, Moskal等人,2002)。
顺便说一句,在最极端的情绪环境下,分离痛苦系统的急剧唤起可能是恐慌发作的潜在原因之一(Panksepp, 2006a; Preter & Klein, 2008)。我们对社会依恋的心理生物学的理解,主要源于对这些神经化学物质的研究,也与对自闭症等儿童障碍的初步理解联系起来。有可能某些患有这种疾病的儿童在社交上冷漠,是因为他们对自己释放的阿片类物质上瘾,而不是对重要他人激活的阿片类物质上瘾(Panksepp, Lensing et al., 1991)。最近,关于自闭症的催产素成分的想法得到了关注(Panksepp, 1992b to Green & Hollander, 2010)。
如果这一分析是正确的,那么我们需要对依恋和分离痛苦(GRIEF系统)的大脑机制,以及它们在抑郁症的病因学、机制、治疗和预防中的作用进行有力的新研究。就像传统的行为神经科学研究一样,此类研究将涵盖从遗传和分子水平,通过解剖和化学,到功能成像、脑刺激和药物试验的全部方法(Watt & Panksepp, 2009)。然而,根据这一研究模型,将特别强调心理学和神经学方法的整合(Panksepp & Watt, 2011)。
如前所述,现代临床前研究除了少数例外,继续关注与精神病学相关的外部症状,而忽视情感的重要性(以及产生情绪的大脑系统)。例如,针对恐惧的研究者很少讨论FEAR回路的性质,而是专注于焦虑样行为的条件反射,以及最近关注他们的工作如何与解决PTSD症状相关(Davis et al., 2010)。[2] 研究人员在讨论焦虑的来源时甚至可能不考虑GRIEF系统,但他们在大鼠中发现了一种新形式的”焦虑”,由称为BNST的”扩展杏仁核”的一部分整合。然而,长期以来人们就知道这是大脑中非常容易引起动物分离叫声的区域;因此他们的发现可能是这种社会情绪反应,而不仅仅是传统恐惧反应的变体。大脑中有两个不同的焦虑型系统,FEAR和GRIEF,它们都促进负面感受。如果我们不区分这两个促进”焦虑”的系统,我们可能会在思维和治疗中犯许多错误。目前,对哺乳动物大脑情绪系统的分析,试图根据情感回路失衡来建模精神疾病,仍然很少见(Panksepp, 2010b)。
更好地理解情绪大脑系统也为促进更好的育儿实践和运作良好的社会指明了道路。最近广泛讨论了可能源于长期母乳喂养、母婴同睡以及充足的儿童早期身体游戏的重要性对儿童茁壮成长的影响(例如,Narvaez et al., 2012),以及婚姻关系的质量对儿童心理健康的影响(Code, 2009)。
这些问题贯穿整个生命周期,其复杂性在此无法涵盖。然而,让我考虑一个每个人都面临的最常见问题——亲人的去世。最有效的悲伤方式是什么?随着现代文化的快速变迁,如何应对这种人生转折的优秀范例越来越少。传统上,家庭成员在社区支持的背景下应对。虽然这仍然是现代社会悲伤的核心特征,但这种做法在许多方面已经改变和削弱。在传统社会中,感知到的社区支持的安全网通常比今天强大得多。尽管许多哀悼的外在形式保持不变,但在现代文化中,随着”社区”的逐渐缩小,人们获得的情感支持往往变得更加脆弱。许多个人主义现代文化所特有的社会流动性削弱了许多人在哀悼中感受到的整体社会支持水平。以下是Ellen Dissanayake对生命悲剧转折如何被构建以维持社会联系并从扩展的”家庭”中获得所需情感支持的生动描述:
传统仪式和习俗在斯里兰卡人的生活中所占的比重远大于我们。斯里兰卡人去世后,哀悼者在一天中陆续来到逝者家中,逝者躺在客厅桌上的敞开式棺材里,周围摆满鲜花。丧亲的家庭成员在门口迎接每一位访客,每当新访客到来时,他们都会哭泣,谈论死亡的经过和逝者的优点。客人进入房子,加入其他客人的行列;他们安静地相互交谈各种话题(我们听到过关于电影、生意和政治事务的讨论);在适当的间隔时间后,他们离开。最终,家人和亲密朋友前往火化或安葬地点,佛教僧侣加入他们,诵读相应的巴利文经文——关于生、死、衰败和轮回的思考。遗体处理三天后,家人和僧侣举行布施仪式;之后在三个月、一年以及此后每年一次举行追思布施。我们意识到,这种正式化的悲伤处理方式,通过定期的、受社区认可的机会,让人们在逐渐拉长的时间间隔内哭泣并表达失去亲人的痛苦,给予丧亲者一种有规律的程序可以遵循,一种可以塑造和包容他们情感的形式。丧亲者不必为了表现勇敢或”现实”而压抑悲伤和失落感,也不必随意或独自释放这些情感,而是通过哀悼仪式得以——被迫——在预定的结构中一次又一次地公开承认和表达这些情感。哀悼仪式的时间结构虽然简单,但确保了关于失去亲人的思绪和感受会在规定的时间得到重申。即使一个人可能没有自觉地产生适当的哀悼情感,连续布施的习俗也能确保这些情感被唤起。规定的正式仪式成为扩展的社会网络公开表达悲痛的场合。(2003年,第19-20页)
这是一种社会形式的情感”疗法”。这些进程凸显了传统文化如何学会以优雅、同情和团结的方式处理我们永恒的悲伤。传统社会中的人们通常深切关心彼此的生活,这使得经历失去的个体能够经历悲伤的各个阶段,从而更不容易陷入抑郁。同样很难想象,如果临床医生和患者之间缺乏基本的依恋感,心理治疗如何能取得成功。因此,治疗关系的质量长期以来一直被认为是有效治疗的关键。
这一观点由人本主义治疗师卡尔·罗杰斯(Carl Rogers,1902-1987)通过其无条件积极关注(unconditional positive regard)的概念得以推进(Rogers,1961,1980)。如果治疗师无法采取一种能够共情他人心理痛苦的立场,就永远不会有那种对治愈之触至关重要的信任感。没有这种信任,无处不在且美妙的”安慰剂效应”(placebo effect)的基础——即内源性阿片类物质介导的社会支持感——就无法在患者心中扎根。没有真诚的共情(empathy),而共情应该是每一次治疗互动的核心,总会有残留的怀疑,一种被操纵的感觉,而不是打开积极改变之门的深度接纳——救赎和拯救的感觉。同情(compassion)(如精神在场的咨询:Brammer,2011)对于有效治疗所必需的关怀立场可能至关重要。
上述观点目前可能是生物精神病学中的少数派观点,但我相信它们反映了一种自然且相当结构化的方式,将神经生物学原因与精神病学的首要情感关注以及体验性、情感导向的治疗联系起来。事实上,敏锐的临床医生开始意识到这些概念图谱、实验探究和神经科学发现如何能够为他们的努力提供信息(Valliant,2008)。尽管在我之前章节对许多临床前事实的叙述中这一点可能并不总是清晰,但我的总体希望是,基于证据的理解——即原始情感(primal affects)如何在心脑(MindBrain)中产生——将促进临床思维。这些方法最终也可能使我们更好地设想未来精神病学诊断方案中情感问题的本质,在那里思维需要沿着情感内表型(emotional endophenotypes)的路线重组,而不是人为的综合征思维(syndromal thinking)(Panksepp,2006a)。同时,显然生物精神病学中关于初级过程情感(primary-process emotions)的研究仍处于早期阶段。
显然,心理治疗师无需被告知情绪失调(emotional dysregulation)是困扰患者的关键问题。这是不言而喻的。许多治疗师目前已经认识到,只有真诚地处理患者的情绪动态,并创造性地、敏感地促进患者情感生活的重构,同时不忽视人类也是认知性存在这一根本事实,才能取得最佳进展。到目前为止,这种多维度的治疗工作更像是一门艺术而非科学。就像熟练演奏乐器一样,临床医生需要扎实、严谨和娴熟的技术,以及基于相关实证理论的广泛知识,来支持那些标志着真正临床艺术性的灵感迸发和突破性互动。心理治疗实践越是建立在情感上合理的思维和技术基础上,就应该变得越持续有效。情感神经科学(affective neuroscience)的结构可以帮助临床医生在方法上变得更加系统化,无论是当下还是日常实践中,从而减少对偶然性和临床直觉的依赖。
儿童尤其如此,如果在临床互动中始终关注他们真实的情感关切,他们会对治疗帮助更有回应。为了更好地治疗他们的社会情感障碍,我们可能应该持续调动他们的玩耍(PLAY)能量。我们每个人心中都有一个小丑。(谢天谢地,因为它可以把工作变成游戏——有时也包括心理治疗工作。)像所有原始情绪冲动一样,玩耍的冲动源自新皮层之下的神经网络。然而,越来越清楚的是,玩耍行为对皮层有着显著的影响,将其编程为完全社会化的状态,基因表达的许多变化正让我们看到治疗抑郁症的新方法(Burgdorf et al., 2010, 2011)。只要游戏能量在临床实践中得到充分利用,临床干预必将尽可能快速地取得积极进展。
例如,共同的笑声可能标志着极具价值的治疗时刻。如果治疗师在处理非常困难的生活境况时,能够促进积极情感,甚至与来访者一起欢笑,那么记忆研究的新发现,如再巩固(reconsolidation),是否可以被用于更持久地软化生活痛苦的边缘?所有记忆在被提取时都是不稳定的。它们倾向于返回到半永久性的”储存库”中,并携带着最近的情感体验结构。根据这一观点,治疗师在转向其他议题之前,将来访者从负面情绪和绝望转变为积极情感的能力,应该成为确保那些负面记忆对来访者自我感受和生活境况的影响力有所减弱的途径。
因此,也许玩耍的积极情感对成年人和儿童一样重要。情绪韧性(emotional resilience)的能力会通过直接的身体性游戏互动而增强。这些真实生活中的人际愉悦在传统心理治疗中很少被使用,即使是在儿童治疗中。儿童不可能在不移动身体的情况下玩耍。当然,成年人可以仅通过言语互动获得乐趣,但人们必须思考,治疗师关注身体并鼓励非常紧张的来访者采取不同的身体姿势是否有用——从坐在椅子上,到坐在地板上,再到站立,也许还用各种非攻击性的情绪性姿态与治疗师互动(我见过Pat Ogden及其同事(2006)演示这种方法,他们将感觉运动(sensorimotor)方面纳入心理治疗方法)。因为初级过程情绪(primary-process emotions)都关乎动态运动,也许这种治疗灵活性可能为真正游戏性的社会互动打开情感表达的”门”,产生丰富的长期治疗影响。
简而言之,我们需要学习如何将痛苦的负面记忆,甚至创伤性记忆,包裹在新的积极情感”包装”中——如前所述,记忆研究的新进展,特别是关于再巩固的研究,促使我们考虑这种可能性。也许没有比在重新体验强烈负面情绪记忆后不久唤起积极情绪唤醒更好的方法来软化困扰性记忆了。如果治疗师能够优雅地将与患者的互动转移到积极情感或游戏性的空间,记忆会以较少痛苦的方式再巩固吗?如果我们对学习的看法是正确的(见第6章),即新的、非条件性的初级过程情绪状态调节学习过程(通过自下而上的信息巩固控制),我们应该能够为最初在负面情感状态下形成的旧记忆提供新的情感-情境变量。我预测,过去记忆的痛苦、破碎边缘可以被”打磨平滑”,以允许积极情感重新语境化(recontextualize)令人困扰的记忆。我和同事已经在临床前研究中观察到这样的效果:压力后的玩耍可以减少抑郁反应;实际上,如果在大鼠暴露于恐惧情境后逗它玩,负面情感的力量就会减弱。
当然,在使用这些策略时,我们必须记住,真实的游戏体验也可能引发负面情绪;这在儿童游戏中尤为常见。在我们尝试评估幼儿游戏干预的效用时(Scott, 2001),我们发现如果立即、温和地处理问题行为,并将回归游戏作为奖励,就可以将问题行为最小化。此外,在与曾经遭受创伤的儿童工作时,必然会面临许多特殊问题,但我们预期,如果能够在信任的氛围中促进这些儿童的社交愉悦,其益处将远远超过更认知化治疗的疗效(Panksepp & Scott, 2012)。为了实现这些目标,仍然需要大量的研究,以及培训治疗师在社交安全的环境中促进自然身体游戏的技巧。
任何能够在相互分享的愉悦时刻把握治疗契机的治疗师,都会将来访者带到快乐生活的门槛和源泉。如果患者能够在身心两方面都停留在那里,治疗师可能提供了心理治疗所能给予的最伟大的情感礼物之一。游戏(PLAY)在从童年到老年的身心治疗中应该占据非常特殊的地位。当然,由于幽默也有其阴暗面——有人最终会成为笑话的靶子,幽默作为游戏的一种形式可能是一把双刃剑。因此,治疗师必须准备好识别新出现的危机,并在当下处理它们。
在操控关怀(CARE)和悲伤(GRIEF)系统方面也存在治疗可能性。这是社交依恋的两面神双胞胎。婴儿的痛苦表达唤起母亲的养育冲动。在更广泛的情境中,我们对他人的痛苦感到自然的同情。深层皮层下情绪共振,包括他人处于困境时的深度共情,似乎是哺乳动物大脑的自然属性。实际上,治疗师或许需要特别擅长使用他们皮层的”镜像神经元”系统来促进情感上有意义的接触和解读。换句话说,他们的身体需要与患者的情绪状态产生共鸣和协调,而不是仅仅成为一个毫无表情的”说话的头”。
观察源自关怀(CARE)和悲伤(GRIEF)系统共同化学机制的化学制剂,包括安全的阿片类药物(例如丁丙诺啡)和催产素,最终如何被治疗性地使用,将会很有意思。这些很可能是促进我们与他人创造积极主体间空间能力的大脑化学物质。这些社交化学物质的药用可能有一天会允许临床医生选择性地增强促进治疗进展的亲社会情感。用养育活动补充治疗情境可能会增加内源性阿片、催产素和催乳素的释放。然而,补充此类激素——例如,在夫妻治疗开始前鼻内给予催产素——也可能通过允许夫妻双方更有效地在当下工作来增强治疗流畅性。
Macdonald和Macdonald (2010)对大量的社交依恋研究和社交疼痛的大脑机制进行了很好的总结。除了大脑阿片类物质的社交诱导安抚效应外,现在已经证明大脑中的催产素在经济决策中介导信任行为,或许还能更敏感地解读他人思想的能力(Pincus et al., 2010)。思维的这些三级加工方面的变化可能主要源于催产素减少分离焦虑和孤独感这一事实;换句话说,它促进自信(Panksepp, 2009c)。在临床前模型中,它可以逆转社交隔离的一些有害和抑郁效应(Grippo et al., 2009)。可以预期的是,在这种天然化学物质的帮助下,过度胆怯、患有”社交恐惧症”的人可能会在与他人互动时感到更舒适。
情感神经科学致力于阐明存在于哺乳动物心脑(MindBrain)中的实际一级加工情感。三角验证方法——整合来自行为、神经和心理分析的证据——是直接的,不依赖于推测。我希望它为进一步研究需要指导临床思维、精神病学研究和发展新的以情感为中心的治疗实践的基础性问题提供了更稳定的平台。使用这种方法,临床前研究人员可以将精力集中在特定的、希望是相关的情绪大脑网络上,而不是精神障碍的模糊行为指标上。由于情绪系统的组织和功能可以在多种物种和良好控制的实验情境中进行研究和评估,情感神经科学渴望为思考一级加工情绪提供比迄今为止更连贯的实证基础。它还让我们看到为什么我们的大多数想法、我们的认知如此显著地锚定于我们的情感状态。在心脑(MindBrain)进化中,感受是第一位的。
这并不是否认大多数心理治疗关系也必须在认知层面进行协商。因此,初级过程情感神经科学尚未为如何在治疗环境中处理心智的情感力量提供确凿证据。然而,它提供了一个替代性视角,希望是一个清晰的视角,说明各种负面情感如何可能导致痛苦和苦难,以及如何更好地利用积极情感来抵消负面情感。与早期情感生活的观点相比,这些原则可能更好地为最佳治疗实践和理论提供背景,因为早期观点缺乏坚实的神经科学基础。
至少,为了理解情感体验的核心本质,我们必须认识到在脑心(BrainMind)发展早期出现的前命题性情感过程,以及它们如何能够独立于人类高级心理过程的巨大复杂性而存在。在婴儿早期,初级过程情感状态并未与认知和语言过程纠缠在一起,而这些过程在生命后期总是与情感相互作用。通过理解我们认知装置极具可塑性的神经生物学本质(Doidge, 2007),它主要是从充满情感的生活经验中动态地、发展性地构建起来的,我们能够更好地理解如何消除麻烦的高级情感程序设计(programming),其中一些已经在表观遗传学上成为大脑适应不良硬件的一部分。
我们应该注意到,本书中对认知和情感过程的区分在当代认知和神经科学的某些领域仍然相当新颖,甚至不受欢迎。大多数对情感感兴趣的研究者,其中许多人来自心理学中基于语言的建构主义传统,声称我们无法在认知和情感过程之间做出有意义和有用的区分。然而,在原始神经层面这是可能的,并且是未来神经科学进步的重要基石。
尽管如此,情感和认知的互动性如此之强,以至于在大脑顶部——在新皮层网络的习得性三级过程功能中——区分变得困难。但它们可以在皮层下初级过程脑心层面轻易地区分开来。这决不是要否认理解认知和情感领域如何互动的重要性,特别是对于心理治疗和其他心理品质的动态系统理论(例如,参见Lewis, 2005)。
概括地说,认知是那些与心智的感觉-知觉门户紧密相连的大脑信息处理功能,而原始情感和情绪反映了一些最重要的脑内组织原则。认知方面与每个个体高级大脑发展的程序设计联系更紧密,而原始情感和情绪代表了祖先遗传的生活工具。尽管情感和认知的互动在每个个体高级心智的独特拼图中不可分割地交织在一起,但我们必须能够将认知和情感设想为不同但相互关联的心理处理类型。从解剖学角度来说,它们就像我们的心脏和骨骼肌一样既独特又相互作用。为了清晰地思考基础问题,我们必须考虑大脑内这些不同控制层次的独特贡献性方面(Cromwell & Panksepp, 2011)。
这倒数第二章面向所有助人专业人士,他们致力于理解人类心智的基础——我们基本的哺乳动物情感本质——以及这些知识如何有助于增进我们对各种人类精神问题的理解。我们可以预期,对情感-情绪网络的深入研究最终将产生有益的知识,服务于那些生活被情感困扰所困的人。在最后一章中,我将详细阐述这些知识对于理解一些长期存在的科学困境的哲学意义,以及关于人类意识和动物心智本质的一些结论性思考。
对于未来的研究者来说,阐明某些新兴心理治疗干预措施如何改变大脑的情感基调尤为重要。我最近亲身经历了一种新型心理治疗——称为眼动脱敏和再处理(Eye Movement Desensitization and Reprocessing, EMDR)的程序——在过去几十年中,它已成为治疗PTSD和其他分离状态的流行方法,但并未被普遍接受。在这些状态中,强烈的情感记忆被区隔化,几乎与心智的其他部分分离,而不是整合在一个统一的、情感功能良好的人格中。在这里,我将叙述我在这种治疗中的个人经历,以及关于这种创伤治疗为何如此有效的一些假设。

作为引言,我想先说明我一生中经历过不少创伤。第一件”大事”发生在1944年,当时我才1岁,在我家人逃离爱沙尼亚躲避红军时,我的下半身被烫伤,几乎丧命。这些婴儿期的创伤记忆可能仍然部分存活在我大脑深处的某个角落,尽管已经减弱,从我的外显认知记忆(explicit cognitive memories)角度来看,已经完全无意识了。不过,这样的经历必然留下了某种情感残留(affective residue),也许反映在一种天生的焦虑和过度担忧倾向上,甚至可能是抑郁。事实上,大量证据表明,早期创伤会增加成年后创伤后应激障碍(PTSD)和未来抑郁症的严重程度(有关总结,见Watt & Panksepp, 2009)。然而,幸运的是,我拥有那种能够克服困难的气质,可以称之为勇气。
我最近的创伤与我在写这本书的整整一年里接受的一系列日益严苛的癌症治疗有关。在西雅图一家世界级医疗机构——亲切地被称为”Fred Hutch”——接受治疗期间,Sandra Paulsen博士(《透过创伤和解离的眼睛看》[2009]一书的作者)好意地为我提供了几次EMDR(眼动脱敏与再加工)治疗(Paulsen博士还为本书提供了插图:[图1.7]和[5.1])。她很想看看我是否能以我的情感神经科学(affective neuroscience)视角,对EMDR多年来在众多患者身上产生如此显著治疗效果的原因提供一些见解。她引导我体验了针对婴儿期创伤以及癌症创伤的EMDR治疗样本。
让我为这一切建立一个医学背景。在近10年的缓解期后,我再次接受淋巴系统恶性肿瘤——非霍奇金淋巴瘤(non-Hodgkin’s lymphoma)的治疗。早在1998年,在双侧疝气修补术前的常规X光检查中,在我的肺部发现了一个拳头大小的肿瘤。这有助于解释我多年来胸部感到的沉重感。但因为没有心脏病专家发现任何问题,我将其合理化为我女儿Tiina去世(1991年耶稣受难日)后深刻慢性悲伤(GRIEF)的残留。也许我的核心自我(core SELF)——嵌入在心血管控制系统中——正在经历持续的悲伤之痛。错了。这些胸部症状在癌症治疗后消失了,这表明主要是巨大的肿瘤压迫我的心包膜。
但在我走上正确的治疗道路之前,发生了一次深刻的打击!当我肿瘤活检的初步病理报告到达时,发现肿瘤的年轻外科医生让我和我妻子坐下,告诉我们,我最多还有一年可活。确实,他对这种大小的小细胞癌的诊断,从统计学上来说意味着我的日子屈指可数了。幸运的是,在一个月的深深担忧之后(在此期间我在安排后事),梅奥诊所的病理学家正确地将肿瘤重新诊断为小细胞淋巴瘤,这让我如释重负,从此我将其戏称为”弱瘤”(wimpoma),因为我了解到它是可以治疗的。尽管我的第一位肿瘤科医生已经在为我提供各种严苛的化疗方案,但在对治疗选择进行了富有信息量的讨论后,我问了关键问题:“医生,您以前治疗过这种淋巴瘤吗?”他害羞地耸了耸肩,回答说:“没有。”我友好地推了一下,立即分享了我的真诚愿望:“那么,我真的很想找一个治疗过的医生。”他笑着回应,并亲切地拍了拍我的肩膀,说”好选择!“在密歇根大学医院肿瘤科连续6周的每日放疗中,肿瘤被成功治疗,我是Allen Lichter的最后一位患者,他后来从医学院院长职位退休,在华盛顿特区的一家医学基金会担任领导职务。
我的淋巴瘤在2007年复发——同样类型的肿瘤,但集中在胃部——这表明是同一疾病的全新发作(事实上,我的联系医生Oliver Press很感兴趣,进行了分子生物学比较这次和之前的恶性肿瘤,但结果证明这不是之前肿瘤的残留)。然而这一次,麻烦的组织——新的克隆——已经扩散,并侵入了我的骨髓。命运使然,我的妻子在同一时间被诊断出患有不同的非霍奇金淋巴瘤。我们最近刚搬到华盛顿州的普尔曼,这样我就可以加入华盛顿州立大学的兽医学院。我们俩很快就被一系列医疗问题困扰,首先是一场几乎致命的肺炎合并败血症,让我在重症监护室”濒临死亡”地待了5天。然后我的第一次化疗周期惨败。接着我经历了几个疗程更加激进的”联合”化疗,不祥地被称为R-CHOP,产生了部分缓解。不幸的是,在6个月内,我的疾病进展远远超过了诊断时发现的阶段,表明它已经迅速对常规化疗产生了耐药性。如果不尽快用更强的疗法治疗——即干细胞移植——两位医生独立给出了我不超过一年半的悲观预后。命运使然,我的妻子Anesa通过R-CHOP实现了完全缓解,但她也在6个月内复发了。
我们决定最明智的做法是前往西雅图癌症护理联盟(Seattle Cancer Care Alliance)接受治疗,那里已经完善了干细胞移植技术。我们对整个医疗团队的专业知识和经验充满信心,同时也面临着因长期医疗休假而搬迁的巨大压力。2009年春天,当我完成本书初稿时,我也刚刚完成了4个月的治疗,期间出现了几次轻微副作用和一次危及生命的重大副作用(一种抗生素耐药的”超级细菌”)。我当时处于部分缓解状态,但残留病灶通过局部胃部放疗得到了清除。我的妻子也实现了完全缓解。在整个治疗过程中,她担任我的全职护理人员,因为没有人可以在没有全天候协助者的情况下进入移植诊所。我们一起经历了多次急诊室之行——我去了很多次,她也去过一次。我感谢她在整个过程中表现出的奉献精神和勇气。对我来说,经历了六种连续的治疗方案,逐渐升级,伴随着越来越多的疲劳和各种形式的身体不适。总的来说,这段经历至少可以说是轻度创伤性的,我很想看看心理治疗,特别是EMDR(眼动脱敏与再加工疗法),是否能带来一些益处。
在EMDR治疗中,保尔森博士首先使用了一种”早期创伤”方案,旨在处理内隐的婴儿期记忆。然而,我无法提取任何婴儿期创伤的外显记忆(也许我可以使用阿特·贾诺夫的革命性方法——参见他2007年关于《原始疗愈》的著作),所以我们继续采用她专门为处理癌症创伤问题而设计的标准EMDR方案。大多数经历过艰苦医疗程序的人,都会遭遇许多医源性副作用——也就是治疗本身引发的额外医疗问题——有很多可抱怨的。我当然也不例外。
我的脑海中充满了自体干细胞移植后的新鲜而令人不安的记忆,期间我经历了超级细菌引起的血液和胃肠道感染。这导致频繁的医院就诊和多次抗生素治疗失败(我不愿回忆),需要进行的诊断测试多得我永远不想重复。这导致我手术植入的静脉希克曼导管(Hickman Line)(一种放置在心脏附近的导管,如果运作良好,可以避免无休止的静脉穿刺)被反复取出和重新植入,因为担心它可能是感染源,随后又换了两根PICC导管(外周静脉慢性导管),但由于渗漏和其他问题,它们经常不得不被拔出。最终,我的血液感染在整整一个月的时间里通过每天三次自我注射美罗培南静脉输液得到了控制。我还厌倦了医源性情绪副作用——从每天的深度疲劳和冷漠,到许多次凌晨4点在恐惧中醒来(为什么在这样的凌晨时分,负面情绪通常会占上风?),经常担心我可能会失去我的妻子、我的生命,以及如果最后一种可用的抗生素在杀死革兰氏阴性超级细菌之前无法奏效,我就无法完成这本书。确实,我准备好尝试一些EMDR了。
EMDR治疗创伤的疗效由弗朗辛·夏皮罗(Francine Shapiro)发现(2001, 2002)。治疗包括系统地提取创伤记忆,然后在治疗师指导下进行横向眼球运动,以减轻这些记忆的情感强度。据报道,通过各种简单注意力活动的双侧重复,创伤记忆的力量会减弱。实际上,除了横向眼球运动,还可以专注于交替拍打膝盖,或者先听一只耳朵的音调,然后听另一只耳朵的音调(换句话说,不同类型的双侧刺激)。保尔森博士和我决定使用直接的”标准EMDR”方法。正如她所说,我们将尝试”清除任何关于癌症诊断本身、治疗(包括医源性影响)以及诸如对医疗行业、保险行业的愤怒、对亲人的担忧、对死亡的恐惧等干扰,或任何其他在顺利航行道路上设置情感障碍的事情”。
我与桑德拉共度的几个小时是令人大开眼界的(更不用说”眼球运动”的)经历:她让我系统地提取与癌症治疗相关的情感感受(这一点也不难做到),然后她立即让我跟随一排在大约一码尺长的横杆上来回闪烁的灯光。虽然我自我诱导的情感感受清晰明确,通过大量的自传体记忆储备不难提取,但一旦我开始移动眼睛,这些感受就会迅速消退。这一现象在不同的感受中反复出现:愤怒、焦虑、悲伤等等。它总是非常迅速地起作用。换句话说,一旦我通过双侧眼部刺激转换到外部感官注意力框架,情感强度就像热煎锅上的黄油一样融化(但没有滋滋声)。如果反复进行,这种治疗据称可以像其他任何心理治疗一样快速有效地化解创伤记忆。虽然这需要更多的实证评估,但EMDR对我来说是一种相当直接的ABT(情感-行为疗法)。目前,还没有人完全了解大脑中发生了什么(van der Kolk, 2006),但无论它如何起作用,EMDR都可以带来深远的帮助。
那么在EMDR治疗期间,大脑中发生了什么?为什么如此简单的程序会产生如此显著的情绪益处?让我分享一些关于它如何起作用的理论推测。目前有相当多未经测试的理论流传,我经常被问到我最喜欢的观点是什么。我通常提出的观点是,探索性眼球运动代表了一种基本的灵长类SEEKING反应。这种扫描运动在上丘深层组织,就在PAG上方,而PAG是阐述所有初级过程情绪的最重要的大脑区域(更多的负面情绪集中在背侧PAG,更接近眼球运动回路,而更多的正面网络位于腹侧)。从眼球运动区域向下有神经连接,特别是连接到PAG的背侧负面情感区域。如果它们被证明主要是抑制性的(例如,可能充满GABA),我们可能会对为什么负面情绪会随着探索性眼球运动的开始而迅速消散有一个现成的解释。它们可能主动抑制我们大脑中一些最令人痛苦的神经回路。当然,还有许多其他可能性——从认知重新聚焦、有限的注意力资源、自上而下的情绪调节等等(这是每个观察的科学可能性的本质)——但这些都还没有用关键证据来证实。
为什么情感益处会持久,创伤性记忆在情感上被”软化”,尽管没有被遗忘?有相当多未经测试的理论流传,但这里”记忆再巩固”的概念可能再次发挥作用(见第6章),就像本章主要部分讨论的许多可能性一样。首先,EMDR治疗师通常会建立一个”安全岛”,在其中可以系统地重新处理创伤性记忆。例如,在心理治疗师Katie O’Shea的EMDR协议中,来访者(i)首先被指导如何将未解决的情绪材料放在一个”想象的容器”中,然后(ii)训练”在安全情境中获得感到安全的能力”,例如治疗联盟,最后(iii)使用EMDR”将自动情绪回路重置到健康的反应水平”(2009年,第290页)。换句话说,EMDR情境可能允许人们访问与创伤性记忆相关的情感,然后在安全领域内相当快速地重新语境化它们。
最后这种可能性与最近基础学习和记忆研究中”再整合”或”再巩固”的有趣发现一致。使用动物模型的大量临床前工作现在已经表明,被提取的记忆往往会带着修改返回到它们的记忆库(Nader & Einarsson, 2010)。如果这种记忆修改可以在初级过程情感水平上进行,人们就可以很容易地理解为什么关于过去创伤事件的认知信息不再充满负面情感。换句话说,情绪刺痛已被提取,情感”炎症”已通过在安全感中逐渐重新语境化情感负面记忆而得到抚慰。
当然,这目前只是一个理论,而不是已证明的神经科学事实。事实上,我们目前对心理治疗益处的大部分了解都属于这一类别,尽管越来越多的大脑成像研究已经表明各种疗法——认知行为疗法、精神分析和人际关系疗法——如何改变已知控制情绪性的大脑区域内情感唤醒的平衡(Cozolino, 2002, 2010; Doidge, 2007)。
相比之下,本书中提出的关于初级过程情绪的”控制位点”的观点,以及其他哺乳动物经历类似情绪的事实,都是基于大量事实。任何不相信这些结论的人都有责任用其他方式解释所有这些数据。我强烈敦促许多仍然不相信动物有情感感受的行为神经科学家,不要只是反对这个结论,因为大多数人倾向于这样做,而是要通过实验否定结论所基于的数据。这就是科学的运作方式,但出于某种原因,在这个话题上,观点长期以来一直凌驾于事实之上。因此,我最近选择在2011年写了一系列坦率的论文,讨论行为神经科学需要认真对待其他动物的情感感受(例如,Panksepp, 2011b, 2011c)。

当他们(我的长辈)命名某个物体时,并相应地向某物移动,我看到了这一点,我明白了这个东西被他们在想要指出它时发出的声音所称呼。他们的意图通过身体动作表现出来,就像所有民族的自然语言一样:面部表情、眼神的变化、身体其他部位的运动,以及表达我们在寻求、拥有、拒绝或避免某物时心理状态的声调。因此,当我听到单词在各种句子中反复在适当的位置使用时,我逐渐学会理解它们所指的物体;在我训练我的嘴形成这些符号之后,我用它们来表达我自己的愿望。
——圣奥古斯丁(343-430),维特根斯坦《逻辑哲学论》题词
我们知道所有人类语言都是习得的。我们的大脑回路中是否有内在的”语言本能”,还是仅仅是一种进化而来的交流欲望,这仍然是未知的。但我们确实知道,我们的交流欲望与古老的皮层下过程密切相关(Lieberman, 2001)。我们所有的认知能力也是如此(Koziol & Budding, 2009)。与所有其他哺乳动物一样,人类的高级心智植根于我们祖先的情感,而对于语言习得来说,或许植根于我们的社会情感:事实上,小婴儿在开始吸收命题内容之前,首先会被语言的韵律语调和旋律所吸引。我们”音乐性”的情绪语调可能是语言习得的门户(Panksepp, 2008b; Panksepp, 2009/2010)。
在哲学家路德维希·维特根斯坦用作其《逻辑哲学论》题词的圣奥古斯丁的上述反思中,我们发现了对学习反映世界的大脑过程的直观把握——镜像神经元在精炼语言和社会理解中的作用(Iacoboni, 2009b; Rizzolatti & Sinigaglia, 2008)。圣奥古斯丁反思了我们先天倾向背后那些神秘的情感心理状态——拥有、拒绝和回避世界的各个方面——我们原始的情绪”行动中的意图”(见图1.4)。很少有现代心智学者设想过我们先天的情感感受对我们成为何种认知生物的重要性(但参见Davies, 2011),因为我们成长为具有物种典型倾向去习得语言的生物(综述见Panksepp, 2008b, 2009/2010)。
大卫·休谟(1711-1776),这位撰写了著名的《人类理解研究》(1748/1910)紧随其《人性论》(1739)之后的自然主义哲学家,是意向性情感观点的重要支持者。他提出的观点是,人类行为受到情绪感受的强烈影响,这一观点在他那个理性主义时代以及许多世纪以来基本上处于休眠状态(见McGilchrist, 2009),直到达马西奥在其《笛卡尔的错误》(1994)中有力地复兴了它。但达马西奥当时提出了他自己可能的错误:即人类感受主要源于高级大脑功能。我们已经注意到,当我们完成这本书时,达马西奥在他的第四本书《自我来到心智》(Damasio, 2010)中已经完全重新考虑了那一观点,在那里他支持关于心智生活起源的更深层的皮层下进化观点,与这里和其他地方发展的观点没有太大不同(Panksepp, 1982, 1998b),但直到最近他还没有完全接受原始情感已经在皮层下大脑区域中完全发展的观点。在他的新观点中,他接受了。
然而,大多数神经科学家和心理学家对心智的皮层下来源保持沉默、不可知论或否认态度。他们在大量的学习研究中使用奖励和惩罚来训练——强化——他们的动物。但许多人似乎仍然相信,就像我们的行为主义前辈一样,动物什么也感觉不到——情感感受的大脑机制并不对学习和记忆过程做出贡献。人类研究长期以来表明并非如此。动物研究的证据也长期以来支持相反的结论。但目前,跨物种大脑科学中关于情感体验在控制动物行为中的作用的沉默是震耳欲聋的。
为什么大多数神经科学家选择对他们研究的动物的情感感受保持沉默,这对”局外人”来说是一个谜……但答案很简单:研究在无情的还原主义原则下成功发展,该原则认为大脑机制和行为很重要,但动物大脑中的心理活动并不重要。这就是资金所在。这种偏见长期以来一直不利于我们对人类情绪的科学理解,以及知识界对心智生活根源的科学认识。它也使大多数公民——他们本可以从更好地理解各种情感感受以及祖先情绪如何控制他们的高级意图中受益——在这个后现代时代对他们的心智比他们应有的更加困惑。
显然,情绪感受已经进化。正如达尔文在《人类的由来》(第127页)中推测的那样,哺乳动物心智生活的差异是”程度而非种类”的差异。我们的情绪感受有着悠久的进化历史,这些感受的祖先根源仍然被许多现存物种所共享。这对科学来说是好消息,对人类来说也应该是好消息。这种知识可以将我们的物种重新定位在先于我们的心智生活框架中,同时尊重我们在认知深度方面巨大且肯定独特的能力。这最后一章不仅旨在提供本书的概念概要,而且旨在坦率地处理大脑科学内外的文化抗拒,即承认理解其他动物的原始情绪体验对于理解我们自己感受来源的重要性。
为了将本书总结的诸多证据线索放入背景中,让我们回顾一下杰里米·边沁(Jeremy Bentham,1748-1832),这位功利主义之父,是如何建议我们设想人类情感的。他指出”效用”反映了”任何物体中的那种属性,使其倾向于产生利益、优势、快乐、善或幸福……或……防止伤害、痛苦、恶或不幸的发生”,并强调”自然将人类置于两位至高无上的主宰——痛苦和快乐——的统治之下。只有它们才能指出我们应该做什么,以及决定我们将要做什么”(《道德与立法原理导论》,1779/1879,第1页)。显然,正如所有行为主义者本应清楚的那样,他们用来奖励动物的”物体中的属性”之所以起作用,是因为它能够唤起积极情感。对于”惩罚”来说难道不是一样的吗?虽然正如许多介导奖励的情感一样,存在着大量多样的负面情感。如果宇宙中不存在所谓的心灵微尘(mind-dust,威廉·詹姆斯创造的一个短语,暗示无生命物质可能包含某种原始意识),那么无生命的物质世界,当它在大约十亿年前凝聚成复杂生命时,找到了传递支持生命的内在价值的解决方案。这种属性就是情感体验(affective experience),完全由神经活动构建而成。心智伴随着某些类型的神经回路活动而产生,可能至少可以追溯到在大脑深处创造有机体情绪连贯性的古老网络。
目前,我们可以确信哺乳动物大脑具有许多内在的情感价值,这些价值在种类上(即使不是精确形式上)仍然为人类和所有其他哺乳动物以及鸟类,可能还有许多其他类型的生物所共享。然而,很少有神经科学家或心理学家知道(或研究)情感是如何在哺乳动物大脑中构建的,这主要是因为行为主义的极化效应,以及最近心理学导向的研究界未能承认,甚至可能未能欣赏动物模型对于探讨意识本质的重要性。这必然是由于动物大脑研究者在这个话题上仍然保持着震耳欲聋的沉默。然而,我们在精神病学中一些最紧迫问题的答案必须来自对情感状态的”临床前”模型的使用,这些模型源于情绪系统的秩序与失调。
脑心智(BrainMind)显然是一个进化分层的器官,建立在情感基础之上,主要通道在大脑组织中仍然明显可见——更古老的功能集中在更低和更内侧的脑区,而更新近的功能则位于更高和更外侧的区域。在进化框架内,动物大脑研究可以为理解人类情感的基础提供最深刻的指导。实际上,通过这样的工作,我们最终可能会理解人类的情感体验是如何从哺乳动物大脑动力学中产生的。这并不是说动物会发展出人类那种复杂的认知-情感情操(cognitive-affective sentiments),它们也不会像我们那样反复思考自己的不幸,但我们应该认识到,以原始形式遗传内置于动物脑心智中的初级过程情感(primary-process affects),与指导人类大脑情感倾向的情感并没有太大不同。令人遗憾的是,人类心理学和哲学中看似无休止的概念辩论常常淹没了神经进化敏感的动物研究长期以来提供的经验信号:所有哺乳动物都是高度情感化的生物。
为了从实践和临床角度考虑上述哲学问题,我想再次向所有对心智感兴趣的神经科学家和生物精神病学家提出以下问题:神经科学中最重要的问题是什么?当然会有各种各样的答案,从记忆的分子本质到介导认知的神经”计算”,偶尔有人投票给自由意志的本质。也许许多生物精神病学家目前会把希望寄托在精神疾病的特定大脑和遗传基质上,少数人投票给意识体验的本质。我会投票给:“原始情感体验是如何在大脑中创造的?”为什么这如此重要?答案可以帮助阐明一般体验的基础性质(即初级过程意识),以及人类灵魂可能遭受的各种情感障碍(Solms & Panksepp, 2012)。
因此,对于抑郁症,我会特别问:“为什么抑郁症感觉如此糟糕?”为什么抑郁症会带来痛苦?为什么它在心理上如此痛苦?体验社会性痛苦意味着什么(MacDonald & Jensen-Campbell, 2011)?很少有神经科学家愿意提出这样的问题,但一些工作假设已经从情感神经科学视角对初级过程情绪性的研究中获得,这基于约翰·鲍尔比(John Bowlby)的开创性观点:悲伤(GRIEF)的唤起——即与母性关怀(CARE)分离所产生的急性心理痛苦——如果持续存在,会导致持续的绝望,而这正是通向抑郁症的门户(Panksepp & Watt, 2011; Watt & Panksepp, 2009)。同样,成瘾不仅由积极情感维持,还由强烈的消极情感的潜在可能性维持,这些消极情感在内部积累,因为人们通过改变心智的药物寻求快乐(Kassel, 2010)。通过阐明实验室大鼠具有一组独特的情感性积极发声,我们现在可以使用这些测量作为动物心智在”情感空间”中位置的直接指标,这可以为成瘾、抑郁症和总体幸福感提供新的理解(Brudzynski, 2010; Burgdorf et al., 2007; Knutson et al., 2002; Panksepp, Knutson et al., 2002; Zellner et al., 2011)。
很少从深层神经进化视角看待心智本质的心理学家,开始接受积极和消极情感的某些方面是人类心智进化物理景观的一部分(Lambie & Marcel, 2002)。即使是一些顽固的社会建构主义者和那些赞同情绪维度观点的人(这是当前学术心理学中的一股强大力量),也准备接受人类情感的生物学基础(Barrett, 2006; Russell, 2003, 2009)。然而,这些人类情绪的研究者并不轻易接受在初级过程层面存在比感觉良好或糟糕更高度解析的情感生活的证据。
心理科学界普遍未能认识到大脑组织的初级过程情绪方面(也有例外:如 Buck, 1999; Izard, 2007),这使得许多类似的辩论缺乏神经考量的支撑,因此主要局限于非常困难且本质上令人困惑的三级过程考量:心智的这些更高层次肯定在很大程度上是社会建构的,导致了巨大的个体差异性。但动物大脑研究表明,人类大脑中也必然存在许多天生的情感,不仅因为本书中讨论的经验证据,还因为这将是进化构建大脑的明智方式。
如果初级过程情感具有任何进化功能,除了简单地指导学习之外,那就是内在地预测未来的生存需求。例如,如果情感为行为提供即时的无条件”评价性”指导,那么对威胁生存的各种内部状态和外部刺激,以及那些促进令人满意甚至快乐生活的刺激,拥有准确的情感信号将最为有用。当然,这并不否认某些初级过程情绪可能跨越各种情感(例如,特别是寻求系统(SEEKING system)产生的欲望和兴趣的冲动),我们也不建议高级大脑功能不能以独特的人类方式进一步解析情感感受和意义。一种复杂的人类反思性-情感性意识随着学习和思考而出现。鉴于在脑心智(BrainMind)进化的许多层次上存在的层级系统,许多复杂性在脑心智功能的嵌套层级中实例化,其中较低的情感性大脑功能在较高功能中被重新表征。随着时间和教育,较高功能发展出对情绪表达的递归监督(执行)控制(见第2章,图2.3)。
但在其核心,原始情感是促进生存的内部评价过程。从存在主义角度看,它们是使我们的体验对我们重要的大脑过程,不仅在生存方面,而且在日常价值观方面。它们是行为主义者用来将动物学习塑造成几乎任何他们希望的形式的奖励和惩罚——无条件刺激和反应——除非他们试图违背动物最强的本能。例如,几乎不可能训练大鼠为了食物而在迷宫中倒退跑。多年来,许多带着行为主义偏见来到我这里的学生都尝试过但失败了。这是因为大鼠是通过进化选择设计的,用它们的鼻子而不是屁股来追求世界的果实。
如果没有从爱到恨的各种情感感受,它们用意义为我们日常生活的结构着色——从日常的欢乐和折磨到伟大音乐、舞蹈、戏剧和其他艺术的微妙且有时崇高的情感丰富性——我们的生活会是什么样子?我们的情感生活诱使我们珍惜和厌恶世界上的各种事件和物体,如果没有我们的情感能力,其中许多将没有心理深度,没有深刻性。因此,无论在我们人类心理世界中存在什么基本价值,它们都存在于人类大脑内部,并在很大程度上存在于我们从早期动物那里继承的祖先心智中。
我们的核心价值观源自于进化而来的情绪——以及我们大脑中许多古老网络的激励响应特性——尤其是那些集中在所有哺乳动物共享的内侧皮层下脑区的同源复杂网络,这是因为它们有共同的祖先。这些心智的原始力量通过学习与次级生活经验相连接。当然,如果我们人类没有情感体验(emotional feelings),我们就不会费心在其他动物的大脑中寻找它们。但在跨物种科学地探究这些问题时,现代情感神经科学(affective neuroscience)最终向我们保证,我们不仅仅是在沉溺于无意义的拟人化(anthropomorphism)(就好像现实位于我们真理图的右下象限,图1.5)。这使得比较神经现象学(comparative neurophenomenology)的研究——即对心智脑(MindBrains)内部心理内容的研究——成为跨物种的一项极其重要的科学事业(Panksepp, 1999)。在其他动物中,情感状态是它们心智内容中最容易研究的,因为产生情绪性行动(actions)(而不仅仅是”反应”)的神经回路易于观察,并且与动物的情感体验紧密交织。这允许一种双重视角认识论(dual-aspect epistemology),即可观察的行为可以用作隐藏感受的代理(proxies)(Panksepp, 2005b)。
然而,情感体验是彻底主观的,迄今为止没有任何物理科学接受物质世界隐秘角落中存在任何主观性。尽管达尔文(Darwin, 1872)在《人类和动物的情绪表达》(The Expression of Emotions in Humans and Animals)中开创性地接受了动物的感受,但二十世纪初行为主义(behaviorism)和逻辑实证主义(logical positivism)的统治对其他动物是否具有指导其行为的情感体验的科学讨论施加了严厉的、长达一个世纪的限制。在实验层面,特别是在脑研究中,这种对话几乎没有重新开启(例如,Mendl et al., 2010; Panksepp, 2010a)。
由于动物感受的边缘化,基本人类价值观(即情感状态)的神经科学与能够凭经验解决此类问题并能够阐明我们自身情感体验本质的相关动物模型脱节了。尽管有现代脑成像技术,但这些感受的基础神经机制尚不能在人类身上进行任何因果细节研究,尽管相关性分析强烈表明我们可以确信主要的控制来源是皮层下的(subcortical)(例如,Damasio et al., 2000; Northoff et al., 2009),位于我们与其他动物共享的古老脑区(Panksepp, 1982, 1998a)。现在,随着心智科学中进化观点的日益被接受,动物模型可以开始填补我们对人类心智的初级过程情感基础理解中的许多空白。
目前,人类”心智肉体”(mind flesh)的功能细节及其如何产生内部主观体验的感受必须从不精确的测量中推断——即主观自我报告结合现代功能性脑成像,这最终支持了基本情绪观点(Vytal & Hamann, 2010)。然而,最近有人指出,使用这些技术时大脑和心理变化之间的相关性相当一致且令人怀疑地高(见Vul et al., 2009,附六篇评论)。由于需要大量平均才能使数据具有功能意义,许多观察到的脑-心理关系可能在很大程度上是由于相关性分析中出现的统计伪影(statistical artifacts),即在计算相关系数之前汇总数据时产生的偏差,这一偏差曾一度阻碍了我自己在能量平衡调节方面的工作(Panksepp, 1973)。
遗憾的是,尽管我们人类有科学上的傲慢,我们在这一领域令人印象深刻的人类脑成像工具仍然更像伽利略的望远镜而不是哈勃太空望远镜。现代功能性脑成像主要提供关于大脑中值得详细实验审查的感兴趣区域的证据。这种精细审查在人类研究中几乎不可能实现。但这些技术可以为我们提供各个脑区如何协同工作的统计估计(区域血流变化的相关性,产生连接图谱(connectivity maps)的统计估计),这可以与实际连接性相关联。最近的改进允许通过使用弥散张量成像(diffusion tensor imaging, DTI)来可视化大脑中的主要束(tracts)(连接脑区的通路),甚至可以突出显示外科医生在精细心理外科手术(psychosurgery)时代可能对情绪网络做了什么——例如,汇聚的证据表明抗抑郁神经外科手术可能一直在放大寻求系统(SEEKING system)的积极感受(Schoene-Bake et al., 2010)。这表明直接刺激寻求系统应该会产生抗抑郁效果(见下文Schlaepfer和Coenen的工作)。
人类现有的一些因果性工具(例如精神药理学干预和脑深部刺激[DBS])可以通过与主观状态变化的相关性联系到心理过程。最近,在许多其他治疗都无效的患者身上观察到了强烈的抗抑郁效果,这是通过在前扣带回区域,特别是Brodman Area 25区域进行局部脑深部刺激(DBS)实现的(Mayberg et al., 2005),该区域是GRIEF系统的情感源头(见[图9.1])。据推测,Area 25区域的DBS破坏了GRIEF系统的功能,迅速缓解了抑郁性心理痛苦。在SEEKING系统的前端区域(伏隔核)使用DBS也产生了类似的效果。例如,Thomas Schlaepfer及其同事(2009)报告如下:
开启刺激后,一名患者……自发地表示他意识到自己在科隆,他从未参观过著名的科隆大教堂,并计划在近期去参观,他确实在手术后的第二天去了。当被问及抑郁症状时,他没有报告任何急性主观变化。第二名患者对刺激的即时(60秒)反应非常相似;她没有报告抑郁症状的任何急性变化,但自发地提到她希望重新开始打保龄球(这是她12年前抑郁症发作之前最喜欢的消遣)。她说:“这会很愉快。”这些即时且未经提示的行为反应表明探索动机急剧增加,这与伏隔核在奖励寻求行为中的作用一致。考虑到这些患者在长期抑郁发作期间严重缺乏动机,这一点尤其值得注意。
如前所述,我们现在可以估计过去用于治疗人类精神障碍(例如难治性抑郁症)的各种不同精神外科技术如何在未从其他治疗中获得缓解的个体中产生益处。这些效果似乎是由于对享乐通路(如SEEKING系统的内侧前脑束)的汇聚影响,为难治性抑郁症提供了另一个合理的治疗靶点(Coenen et al., 2011; Schoene-Bake et al., 2010)。当然,这类实验研究问题只能在严格知情同意的情况下,作为先前规定的医学治疗策略的次要附加研究。尽管如此,为了开发这类新型精神病学工具,仍需要进行可行性研究。
可用于动物大脑研究的技术,包括对特定神经系统的电刺激和化学刺激,以及对区域大脑化学物质(包括基因表达谱)的非常详细的测量,对于指导新型因果性研究来说要精确得多(Burgdorf et al., 2010)。然而,由于动物无法谈论它们的体验,太多研究者认为我们永远无法了解它们的主观思维。正如本书所论证的,只要我们认识到通过人工刺激特定大脑区域获得的”奖励”和”惩罚”证明了某些大脑变化对动物很重要,这种长期(几乎一个世纪)的偏见就明显偏离了正轨。
从我们对人类心智从遥远的动物祖先中涌现的认识,加上对进化微妙方式的理解,强烈表明许多其他动物具有与我们自己非常相似的情感生存价值。因此,我们原始情感感受的来源最容易通过跨物种情感神经科学来阐明。我们必须记住,对我们的动物同胞的大量实验工作如何促进了对改善人类生活具有重大意义的医学进步。如果没有对动物的胰岛素研究,过去一个世纪将有数千万儿童过早死亡。动物研究可以阐明支配我们主要情感和相关次级过程学习机制的神经机制的基本原理。当更多学者开始研究这些与意识和人类及动物福祉问题直接相关的议题时,心理学、精神病学和心理治疗将会改变并得到丰富。有理由相信,我们甚至可以重新构想我们文化机构的基础,从哲学到宗教视角(Davies, 2011; Thandeka, 2005; 另见Journal of Consciousness Studies中关于情感神经科学哲学的研讨会[Panksepp, Asma, et al., 2012])。
本书的目标是概述我们对人类精神的这些原始动物基础的知识,以及对帮助人类的意义,省略了许多可以进一步补充总体论证的细节。我们没有充分讨论古老的情感系统如何在认识到进化产生了一丛有时日益复杂的生物分支而非上升阶梯的背景下,与人类更高的认知能力相互作用。目前,我们对这些重要相互作用的精确了解不如对原始情感过程和简单学习形式(如经典条件反射和操作性条件反射)的了解。然而,可以肯定的是,我们巨大的大脑”思考帽”——我们广泛的皮层-认知装置——在心智上将我们与动物祖先区分开来。这增加了无法通过动物模型轻易解决的复杂性层次(例如,Harmon-Jones & Winkielman, 2007; Northoff et al., 2011)。
随着哺乳动物大脑皮层的扩大和复杂化,我们的认知意识也相应扩展,产生了更高级的(例如,三级过程)意识形式(Damasio, 1999),以及一种以自我为中心的主张,即基于语言的理性是人类意识的基础(Fogelin, 2003; McGilchrist, 2009)。正如[第11章]所讨论的,这些”扩展的”认知意识形式仍然与更古老的情感存在形式密不可分。另一种观点,尚未得到现有神经科学的支持,认为认知意识从根本上源于感知外部世界的一级能力,与情感感受没有关键联系。这种观点似乎明显是错误的,因为当皮层下情感基础被大量破坏时,所有形式的意识也会消失(Panksepp, 2005b, 2007a)。语言是我们最独特的大脑技能,但即使语言也是通过情感引导而产生的。然而,通过语言,我们可以独特地研究人类的”扩展的”三级过程认知-情感意识。这就是为什么心理学中描述性(即非神经科学)情绪研究持续大量增长的原因(Davidson et al., 2003; Lewis et al., 2008)。
Damasio指出”扩展意识是一个比核心意识更大的主题,但在科学上更容易研究”(1999,第201页)。我们同意这一点,尽管这种科学必然缺乏机制性,因此在信息量上会较少,至少在因果问题方面是这样。然而,由于研究的便利性,基于人类语言各种用途的情绪研究方法往往会隐含地阻碍对动物一级过程核心情感意识的研究,就像严格的行为主义观点所倾向的那样。研究资金,以及由此带来的快速进展,需要科学界的共识。但在我们原始的动物情绪方面,并不存在这种共识。因此,关于动物情感感受的明确神经科学工作很少(这对理解人类感受具有潜在的深远意义),特别是与人类情绪研究的复兴相比。在西方知识传统中,这些问题中更古老和基础的层面被严重忽视,尽管这些方面对于揭示人类情感感受的本质以及困扰许多人生活的相关精神疾病都极为重要。
我想按照Damasio的观点建议,在当前的心脑科学研究中,核心情感意识的主题在人类身上很难进行实证研究。但是,为了在心理学家关注的许多核心问题上取得实质性进展,现在可能更为关键的是理解人类和动物感受的”一级过程”进化来源,而不是那些备受关注的扩展文化意识。该领域的许多领导者可能不这样看,但我认为他们没有从所有相关的神经进化视角,如一级到三级分析层次,来全面思考这些问题。确实,如果一级过程情感天赋是我们复杂人类心理装置仍然依赖的大脑功能,那么忽视理解我们更深层本质的机会将是悲剧性的,而这些本质无法通过人类研究轻易阐明。
尽管人类可能有理由为我们扩展认知意识的特殊品质感到自豪——比如我们象征性说话的能力,它创造了文化、文明和我们丰富而详细的精神生活——但我们没有可靠的方法来理解我们自己心智的情感基础。这只能通过研究其他动物的类似过程来揭示。我们的许多高级心理功能更像是”意识工具”——完全建立在本书中描述的皮层下过程的基础上,并完全依赖于它们的完整性。因此,我们应该珍惜我们的感知装置——特别是我们敏锐的听觉和视觉——但如果失去它们,我们”仅仅”失去了许多珍贵的意识”内容”,同时仍然是完全有意识的存在。更古老感知装置的某些组成部分,即我们的前庭感觉(vestibular senses),很少被明确体验到,直到它们受损。我们应该认识到,对动物核心情感意识的科学研究提供了途径,也许是唯一的途径,来理解支撑我们高级心理装置的祖先心智。
因此,本书始终关注动物一级过程情感意识形式的本质,而没有试图讨论它们的三级过程心智中”可能”也存在的充满情感的思想——这些过程在动物身上更难研究(Mendl et al., 2010; Panksepp评论, 2010a)。我们需要开启并激发关于动物心智本质的持续讨论,这个讨论在近一个世纪前被行为主义(behaviorist)主宰力量封闭——据称是永久关闭。行为主义者的观点,也许在他们的时代是适当的,但在神经科学时代已被证明是短视的。它导致过早地抛弃了一级过程情感心智,仅仅因为三级过程认知心智在动物身上无法得到很好的研究。而且他们的选择变得过于容易,因为达尔文的思想后继者们过早地使用心理主义(mentalistic)概念来解释动物行为(Romanes, 1882)。
这些决策的残留至今仍以强大但无法估量的负面方式持续存在于我们的大学中。由于行为神经科学家选择受到那些剥离了情感的旧行为主义概念的约束(例如”奖励系统”和”奖励预测误差”——见[第3章]),神经科学家几乎没有关于动物情感过程的直接讨论这一传统仍在继续。情感感受甚至被那些本应更了解的人所轻视。严谨的科学家不希望看到他们的工作被当作拟人化(anthropomorphism)而被忽视([图1.3])。但神经科学证据的主要权重表明,世界上那些被称为”奖励”和”惩罚”的实体实际上是由大脑内的情感变化构成的。事实上,控制一系列情感行为协调的大脑非条件性情感反应系统并非无意识的。这些大脑网络正是情感感受的源头。这种视角上的微小转变可以大大激发我们如何使用精神疾病的临床前模型。我们生活在一个时代,广泛使用的科学概念”强化”应该被视为可能只是对核心情感——动物的非条件刺激和非条件反应——在世界波动事件中运作的复杂方式的简短总结术语。强化的普遍程序非常有效,但强化的过程仍然是一个假定的,而不是被证明的大脑功能。现在,“强化”被广泛认为是一个真实的大脑过程,但它可能只不过是行为分析的燃素理论(phlogiston)(见[第3章]脚注2),逐渐被接受为对现实的描述。
由于动物心智的潘多拉魔盒在几代人之前就被封存了,对动物情感功能的充分讨论仍有待全面展开,特别是对于那些最适合进行必要研究的学术界人士。如果神经科学家之间及时地(至少在20世纪70年代)展开辩论,正如一些人在认知动物行为学(cognitive ethology)中认真尝试的那样(例如,Donald Griffin,1915-2003),我们现在可能会处于更好的位置来研究人类和其他动物祖先情感心智的主观方面。
无论如何,意识的认知形式——即关于我们在世界中特定环境的思考,整合了陈述性记忆(declarative memories)和自传体记忆(autobiographical memories)——在机制上比核心情感更难研究,特别是在动物中。一个简单的原因是,据我们所知,特定的认知不像原始情感那样具有明确的神经通路。此外,核心情感的神经化学编码包括许多调节特定情感的神经肽([图13.1])。这一事实对于跨物种预测来说是一个福音。未来的神经肽研究应该能够测试这些神经控制是否能在动物和人类中产生可比的情感变化;这些类型的效应已经通过催产素(oxytocin)和社会情感得到了很好的验证(见Panksepp, 1992, 2009c; Pincus等人, 2010)。
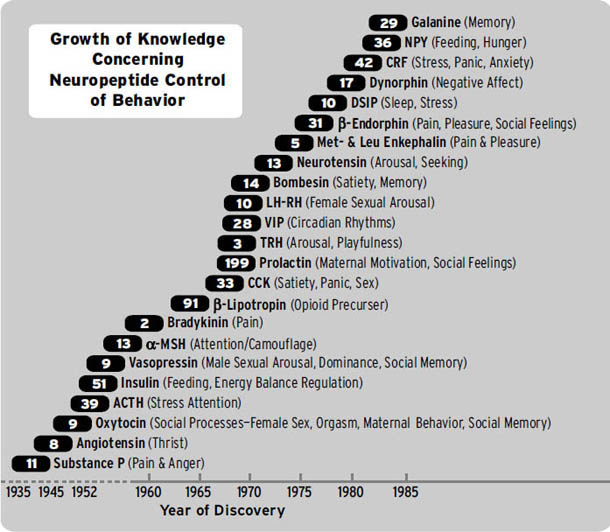
图13.1. 参与控制行为以及各种情感和动机过程的相关大脑功能的主要神经肽的发现时间线。开始时进展缓慢(见虚线),但在1970年左右大幅加速。方框内的数字表示这些神经肽中每一种的氨基酸数量(来自Panksepp, 1998a;经牛津大学出版社许可改编)。
换句话说,与认知相比,情感的神经化学编码的丰富性,允许预测的丰富交流通过从动物大脑到人类心智研究的坚实神经科学桥梁流动,并再次返回。相比之下,我们所有的认知更严格地依赖于动态变化的谷氨酸能(glutamatergic)兴奋性传递,由GABA能(GABAergic)抑制性引导机制塑造。此外,皮层认知和感知也肯定需要比皮层下核心情感感受更复杂和快速的神经元放电模式。控制认知的大规模复杂神经动力学必然会在不同物种之间变化更大。这将使对高级心理过程的神经科学理解成为一个比进化上保守的初级过程情感的跨物种研究困难得多的问题。
尽管关于思想的思想,由命题语言(propositional language)介导——在成熟的人类中发展得如此完善——在大多数其他动物中肯定不存在,但我们不知道有什么方法可以排除这种可能性。其他哺乳动物可能更多地用内部感知图像来思考,这可能更好地反映了动物记忆和思想的祖先形式(Grandin, 2005)。但这很难评估,除非通过全局脑成像的相关性(例如PET扫描)。目前几乎不可能进行良好的因果研究。因此,我们必须对人类和动物认知的三级过程层面的同源性推迟任何确定的结论。另一方面,作为条件行为基础的基本二级过程学习机制,主要是无意识的神经机制,允许非常有效的跨物种转化(LeDoux, 1996, 2007)。
然而,没有证据表明这种条件控制为动物体验的情感感受类型增加了多少多样性,因为学习只是修改了情绪的强度以及时间和空间表达。例如,当条件反应逐渐取代非条件反应(UCRs)时,可以预期情感会大幅减弱。大脑中至关重要的UCR机制不仅对产生情感至关重要,而且可能为条件刺激唤起条件反应提供了”粘合剂”——即”强化”(参见第6章)。
同样,理解初级过程情感和相关的条件反射过程,对于我们自己的三级过程、自我反思倾向可能至关重要,尽管我们目前还没有关于这如何发生的明确科学数据。显然,我们所有更高层次的心智复杂性——从科学研究到哲学、精神分析和艺术——都需要我们记住许多基于符号、语言的记忆,以及过去和未来可能性的庞大模式。我们不知道有任何动物拥有类似的大脑技能,但我们确实知道许多其他动物以自己的方式相当聪明(Romanes, 1882, to Griffin, 2001),并且它们确实是充满活力的情感生物。这给了我们特殊的责任,要求我们以特定方式进行研究,并照顾所有动物生命。这一知识对动物福利问题的影响是巨大的(Bekoff, 2000; Grandin, 2005; McMillan, 2005)。
我们有意没有详细讨论高度分辨的感知-认知心智装置的本质。这样做并不意味着我们否认独特的扩展意识在使我们完全且深思熟虑地成为人类方面的重要性。我们的目标是开辟智识空间,通过培养一种充分尊重其他动物情感生活的观点,对人类激情的祖先来源进行现实而有效的科学探讨。我无意贬低每个动物物种、每个独特的人类个体和人类文化为我们这个世界丰富的心智存在盛宴所带来的许多重要差异。我主要寻求的是那些仍将我们彼此联系在一起的心智一般原则……就像一个需要回顾其共享祖先财富的大家庭。现在有许多实证发现支持我所提出的观点,还有更多有待发掘。
对我来说,情感只是认知的一种变体这种广泛流传的说法不过是文字游戏,尽管我确实接受神经系统的许多赋值(好与坏)感受总是在大多数人类和动物心智的全部复杂性中与认知(想象、学习、记忆、思想)相互作用。注意力和原始动机(例如,口渴和饥饿)以及所有心智的祖先功能也是如此。就像身体器官一样,我们内部的一切都在相互作用。我在神经科学中学到的第一课是,进入眼睛的频闪视觉信号的特定频率波纹可以在大脑的几乎每个角落测量到。这并不意味着在追求理解视觉的真正进展时,我们不应该将视觉系统与听觉系统或其他大脑感觉系统区分开来。
对于那些坚持认为情感和认知在人类心脑中完全混为一谈的人(这可能包括当今大多数心理学家和哲学家),我建议他们不仅从自己的角度考虑他们的论点,还要考虑这里提出的自下而上的观点。当我们从皮层认知的高台俯视心智生活时,是的,一切都在相互作用。然而,如果我们理解认知通常是情感的”侍女”(或使者),这种混淆就不再成立了。积极情绪——迄今为止无法以任何有意义的方式”计算”——可以直接促进积极情感稳态的自下而上促进,或者当人类受到消极感受困扰时恢复良好调节的心智平衡。自上而下的认知技能也有效地服务于寻找大量可以抵消消极感受的积极情感。
因此,这里有一个关键问题:认知和情感是否通过非常相似的神经元原理运作,并且在大脑的相同区域?这里首选的进化自下而上观点给了我们三个以上的充分理由来坚持初级过程情感具有独立存在,其在心脑进化中的历史远比通常被认知概念所涵盖的大脑过程要久远得多:
当动物和人类在生命早期失去其主要认知领域——新皮层——后,其情感-情绪存在仍然保持显著完整。
对于这个问题,“认知处理的主要领域(即丘脑-新皮层轴)与富含情感-情绪处理的领域(即皮层下和皮层中线系统,或SCMS [参见Northoff & Panksepp, 2008],传统上称为扩展边缘系统)之间是否存在重大神经生理学差异?”答案是是的。仅就神经元的放电频率而言,认知-躯体领域富含放电频率非常高的神经元(例如,每秒数百次动作电位),而情感-内脏领域则充满放电非常缓慢的神经元(例如,很难找到许多每秒放电超过十次的神经元)。
在正常的新皮层或丘脑中,没有任何地方可以通过刺激大脑的局部区域而持续获得相同的认知或思维(尽管Penfield证明,通过刺激某些与边缘系统接壤的颞叶区域,可以获得刻板的知觉现象)。相比之下,在SCMS内很容易找到可以重复刺激相同大脑位置并获得相同情感状态的地方。
情感性情绪的古老MindBrain基质不仅是我们行为的调节器,而且还促使我们在社会世界中航行时思考生活的复杂性。最重要的是,情感与认知之间的区别——尽管它们在完整的大脑中相互交织——使我们能够承认其他动物也拥有内在生存价值全光谱的体验。它们具有情感体验的神经系统状态,与我们自己的状态并没有太大不同。
认识到这一事实使我们负有特殊责任,要以持久的敏感性、对我们为获取这些知识而牺牲的动物怀有深深的尊重和关切来进行研究(McMillan, 2005)。遗憾的是,如果不对动物的相关大脑系统进行科学研究,就无法对人类状况及其情感困境有更全面的理解。这个结论与它们是有感知能力的生物这一事实密不可分,它们的情感能力源于与我们相同类型的神经基础。人类可能更加”理性”,对自己的心理状态更具”反思性”,但哺乳动物都以情感方式体验情绪。正如Merker (2007)和Shewmon等人(1999)的临床研究所揭示的那样,这些感受源于人类和动物大脑的非常深层区域([图13.2])。显然,我们人类可以比任何其他物种更深入地思考生活的存在意义。毕竟,我们可以说话和进行符号思维。但这并不赋予我们获取原始情感体验的特权。如果我们人类是生命这块密不可分的交织织物中唯一有意识的生物,那将是一个多么空虚和孤独的世界。当我们意识到无论我们的动物同伴在地球上的何处漫游都存在意识的泡泡时,这是多么美妙的慰藉。
图13.2. 一名无脑畸形儿童对被放在她腿上的婴儿的情绪反应(上图)。此类儿童的脑发育不全类型(下图)(数据来自Merker, 2007;感谢Bjorn Merker提供照片使用权。X光片经美国放射学会许可转载(ACR Learning File, Neuroradiology, Edition 2, 2004)。未经美国放射学会明确书面许可,不得对本材料进行其他形式的呈现)。
二十世纪初,行为科学失去了与我们心理装置内在价值的联系。随着二十世纪物理学、天文学、化学和脑科学的显著进步,出现了一种主流科学观点,认为人类存在就像物理科学对物质世界所确认的那样,是无价值的或本质上无意义的。许多心理学家,尤其是行为主义者,认为这是研究小鼠和人类的唯一正确方法。主观体验的神经系统状态被从科学讨论中抛弃,尽管没有从智识辩论中消失。因此,学术心理学与情感性感受疏远了。从动物身体及其行为中获取的冷酷证据才是唯一重要的。
这在很大程度上反映在整个智识生活中。经典的”存在主义”立场认为,很少有内在价值是每个个体的与生俱来的权利——即”本质”,人们必须通过每天的生活重新创造意义。生活本质上是荒谬的,除了人们在自己的存在中构建的意义外,没有更多意义。一方面,这导致了虚无主义;另一方面,它鼓励了所有”意义”都是由每个人创造的观点。对于心智的三级方面来说,这可能是真实的,在那里镜像神经元(mirror neurons)、共情(empathy)和同情感必须通过文化、教育、学习和个体发展注入新皮层矩阵。但它并不能很好地描述进化在我们心理装置的低层次中构建的情感工具(Panksepp & Northoff, 2009)。
随着对行为和心智的科学分析的出现,人们开发出了研究学习和记忆的简单而有效的方法。可以使用任何中性信号——音调、闪光、拍肩膀等——作为CS(条件刺激),然后伴随电击或其他同样强效的UCS(非条件刺激),仅需几次试验,动物就会开始逃跑、僵住和排便,血压升高、心跳加速,而这一切仅仅是因为CS的出现。所有这些之所以有效,是因为许多UCS能够产生众多UCR(非条件反应),这些反应是神经系统极其重要的本能功能。这些相同的UCS可以用于训练动物在”工具性条件反射”(如走迷宫)或操作性条件反射(如按压杠杆)程序中做各种事情。这一切就像钟表机械一样精确运作,但这是因为动物”只是”机器吗?[1] 还是因为进化已将原始情感体验构建到大脑许多UCS和UCR网络的神经矩阵中?科学家们仅仅将动物视为无感觉的机器。很大程度上未被研究的是本能性UCR,至少在情绪领域(如恐惧系统),这些反应对于恐惧条件反射能够如此有效地进行是必不可少的。
一个常被遗忘的历史注释是,很少有研究条件反射的所有环境参数,以及最终研究情绪条件反射的大脑机制的科学家,会停下来深入研究他们实验对象UCR的本质。当然,在行为主义的鼎盛时期这是困难的,因为这需要对神经系统有相当深入的了解和密集的大脑研究。这也可能要求科学家们认真考虑一个现实可能性:UCR在很大程度上是由大脑心智内的情感性初级过程变化构成的。因此,这些科学家用来训练动物的UCS在大脑心智上的各种后果,在”奖励和惩罚”的名目下,从未被概念化为激发进化嵌入神经系统内的原始情感状态。在桑代克的情感律被转化为效果律之后(即世界的”满足”和”不适”被转化为”奖励”和”惩罚”;见第2章),关于动物情感感受的科学讨论几乎停止了。
现在我们知道,实验动物心理学中使用的大多数非条件刺激之所以具有效力,是因为它们不仅引发客观的行为变化,还引发大脑内主观体验的情感变化(即各种不同类型的情绪性UCR),因此开发更清晰的愿景和更聚焦于情感的研究计划,以揭示大脑众多情感”本能”的本质,是至关重要的。但现在这比应该的更困难,因为即使”本能”这个词在那个极端实证主义时代也声名狼藉。此外,要让当代科学家转变研究重点,需要社会在”强化偶联性”上发生转变,具体而言,慷慨的研究资金来源(如美国国立卫生研究院和国家科学基金会)需要鼓励采用更灵活的神经心理学方法来研究动物情绪,特别是动物大脑的众多情感性UCR。如果我们不这样做,我们将永远无法了解人类情绪是如何进化的。这尚未发生,因此很少有科学家坦率地(并以基于证据的方式)谈论其他生物所体验的情感状态。如果我们理解这些原始情绪,我们也将有更好的方式来概念化有机体行为的心理和运动一致性,从而更好地理解核心自我和整个大脑如何作为一个整体运作。
尽管我们人类意识的广度和深度因我们扩大的大脑和文化的智力潜能而得到极大拓展,但实际上,我们继承了构成我们心智内意义基础的古老生物价值。遗憾的是,这种情感性的意义基础可能难以言说。因此,我们最近最伟大的礼物——语言的发现——既是祝福也是诅咒。除了给我们带来优美的歌曲、诗歌和其他文学作品外,它也非常适合播种分歧、异议以及边缘化其他人类和我们共有的动物性本质。心智的初级方面无法仅通过使用文字来理解。这种理解需要神经科学探究,不能拒绝承认古老大脑功能的心理属性。20世纪哲学在打破文化阻力方面往往帮助不大,而这些阻力最终正引领神经科学在理解心理动力学方面取得进展。路德维希·维特根斯坦的有影响力的哲学为我们提供了一个窗口,让我们了解阻碍在理解人类尤其是其他动物心理生活方面取得进展的困境。
在过去的一个世纪中,许多人在我们使用词语的方式中寻求人类意义。当诗人和作曲家展示我们如何能够艺术性地象征我们最深切的渴望、喜悦和绝望时,其他人则在语言逻辑中寻求我们存在和意义的基础。路德维希·维特根斯坦(Ludwig Wittgenstein)(1889-1951)沿着这条道路,寻求语言的终极逻辑。在他的《逻辑哲学论》(Tractatus Logico-Philosophicus)中,他试图提供一个关于语言结构如何与世界结构相关联的确定性陈述。这份宣言由七个命题和大量子命题组成。与我们讨论情绪最相关的是命题6.5:
当答案无法用语言表达时,
问题也无法用语言表达。
谜题并不存在。
如果一个问题能够被提出,
那么它也就可能被回答。
如果将这一规则应用于本书涵盖的情感主题,问题就是:关于情绪感受的本质,是否可能有一个可信的科学答案?我相信这最终是可以实现的,但仅仅是因为神经科学的最新进展。我们现在已经对非凡大脑的复杂性有了足够的了解,可以设想心智如何从神经动力学中涌现,并与身体状态和环境条件密切相关地构建起来。在一个子命题(6.52)中,维特根斯坦继续断言:
我们感到,即使所有可能的科学问题都已得到回答,生活的问题仍然完全未被触及。当然,那时就不再有问题了,而这本身就是答案。
在情绪的认识论中,人们可以认为这是对能否研究自然最深层面(如情感意识)的可能性表达深刻怀疑。确实,有学者认为科学研究我们基本价值观的来源是不可能的。但达尔文时代的脑科学,更不用说维特根斯坦时代的,与我们现在所拥有的相比都很原始。维特根斯坦最接近承认感受的是他在6.522中对心理命题的隐晦概述:
确实有些东西无法用语言表达。它们自行显现。它们就是神秘的东西。
简而言之,他的语言逻辑无法处理情感之谜。维特根斯坦在《逻辑哲学论》的引言和最后独立的第七命题中总结了他的观点:“凡是可以说的都可以说清楚,而我们不能谈论的就必须保持沉默。”在他那个时代——心理学行为主义(behaviorism)时代——情绪感受属于世界上那些”神秘”的方面,超出了命题逻辑的范围,超越了语言,科学无法穿透。心智的这些神秘方面似乎永远超出了连贯的科学分析之外。直到今天,人们仍未广泛认识到这种科学理解现在已经成为可能,或者它必须与识别创造我们情绪感受和价值观的大脑过程紧密相关(Panksepp, 1998b; Russell, 2003)。
在完成他关于知识语言基础的”确定性”陈述后不久,维特根斯坦认识到他的世界体系存在严重缺陷。他在余下饱受情感折磨的一生中,探索意义如何从我们使用词语的极其灵活的方式中涌现。在他的第二本著名著作《哲学研究》(Philosophic Investigations)(1953/1967)中,他耗费了20年心血,在他去世2年后出版,维特根斯坦几乎完全背离了《逻辑哲学论》的严苛立场,开始探索我们如何从无限多样的”语言游戏”中创造意义。
他的思想遗产给我们留下了一种心智生活的观点,其中意义是我们如何玩弄词语的灵活表现。这一直是后现代文化的标志之一,也是社会科学中情绪研究的轨迹——研究我们如何使用词语以及如何在语义上构建情绪。直到最近,在这一传统中工作的学者才开始考虑情绪感受存在更深层的神经生物学现实这一想法——至少,大脑中存在积极和消极情感维度的自然机制(Russell, 2003)。这是进步,但在涉及初级加工(primary-process)情绪系统时还不够。也许情绪的维度理论在三级加工(tertiary-process)层面最有效,在那里我们情感生活的多样性被转化为简化的情绪概念,可以促进研究(Zachar & Ellis, 2012; 另见Jim Russell及其同事编辑的《情绪评论》(Emotion Review)[2011]特刊,以更好地捕捉”基本”情绪理论家所讨论的内容)。无论如何,如果基本情绪和情绪的维度观点反映了心智-大脑组织的不同层次,它们可以很好地协同工作(Panksepp, 2007d)。如前所述,基本情绪的有力证据确实来自现代人脑成像(Vytal & Hamann, 2010)。
当人们思考人性时,似乎最终在心智科学中很少有人能抵抗自然主义的诱惑。即使是维特根斯坦,在上述第二本书中也提到”人类的身体是人类灵魂的最佳图景”(第二部分,第178页)。正是这样一种愿景——聚焦于大脑中所表征的”身体”——我们独立地寻求理解初级加工情绪系统如何在哺乳动物大脑中实际创造感受(Panksepp, 1998b)。
Damasio的理论正沿着这条道路发展,逐渐从我们感受的皮层来源转向认识到情绪-情感过程源自大脑深层皮层下区域。当我完成本书时,我很高兴Damasio(2010)的观点如前所述,正在向我的观点靠拢。我相信许多人会追随这位明智的学者。在《意识之我》(Self Comes to Mind)中,他认识到大脑多样化的皮层下情感网络,在进化过程中,原始心智从原始自我(primal SELF)中产生(或与之共同产生)。各种古老的情绪性、稳态性和感觉性情感是大脑的内在功能,由各种身体输入触发和调节(Denton, 2006)。然而,只有情绪性情感对科学精神病学具有深远意义,我希望这能为临床医生通过心理治疗需要达成的目标提供更好的概念化。
具有讽刺意味的是,维特根斯坦在个人层面上如此接近,但在智识层面上却如此远离对与古老脑心(BrainMind)实在相联系的前语言心理的理解。正如他为第二本书选择的题词所动人描绘的那样——来自圣奥古斯丁的《忏悔录》(Confessions)(第I.8节;见本章开头)——语言部分源于儿童与社会世界的情感互动。从维特根斯坦对心智根本本质的困惑,以及我们通过神经科学能够和不能理解什么的困惑中,他构建了一种关于人性和人际互动的过度相对主义观点,这种观点非常适合人类心脑(MindBrain)的三级过程区域。但这种不完整的视野目前阻碍了心理学成为一门完整的科学,无法充分寻求理解心理生活真正的进化基础。
也许这要追溯到我们学习说话的艰难而漫长的过程。协调的人类主体间性(intersubjectivity)是语言习得事业的核心,包括有节奏的非语言社会信号——一种自然的身体语言——这与我们对世界对象的”寻求、拥有、拒绝或回避”有关,这些对象可以成为我们”自己欲望”的目标。人类的存在不仅仅是感觉联结的问题,尽管这些联结最终填满了我们的心理景观,以至于有时我们几乎认识不到其他东西。应该记住的是,尽管广阔的人类新皮层是维特根斯坦所设想的那种相对主义器官,但它不是我们与所有其他哺乳动物共享的皮层下区域。我们新皮层的巨大计算空间在出生时几乎是空的,它最终知道的几乎一切——用Endel Tulving的术语来说,知性意识(noetic consciousness)和自我知性意识(autonoetic consciousness)(参见Vandekerckhove & Panksepp, 2009)——都是习得的。在新皮层以下的情况并非如此,那里驻留着我们无知性意识(anoetic consciousness),即无理解的意识。
我们的祖先大脑包含基于遗传的特殊类型的意义;原始感受的潜能被建构在身体的本能(即遗传的)神经行动机制中。这些感受有许多种类。有些与来自外部的感觉输入密切相关(感觉的愉悦和不愉悦),有些与进入大脑的内部身体输入相关(例如饥饿、口渴及其满足),还有一些反映行动动力学,至少在原始形式上是进化性地建构在大脑中的。所有这些生存工具都在一定程度上具有可塑性;它们可以通过经验得到强化和削弱。这些原始感受与我们以某些方式向世界伸展的内在冲动以及对我们遇到的原型挑战的反应密切相关。它们不具有高阶意向性(intentionality)(即”行动的意图”),但它们确实具有内在意向性(“行动中的意图”;参见Panksepp, 2003a, 图1.4和1.8)。感受使我们成为主动的有机体,而不是简单的被动信息处理机器。
许多人可能同意情绪感受是我们最早人类交流的根源。但许多人仍然将它们视为感觉过程的一种变体,而不是运动过程,也就是说,更多是被动的(即发生了什么的感受——可能是三级过程的观点)而非主动的(即天哪,我要让这件事发生!——一级过程的视角)。事实上,情绪感受和意识本身可能与运动-行动过程的关系不亚于与感知-知觉过程的关系(参见[第12章])。我们的知觉心智以及我们祖先的情感心智似乎都锚定在行动坐标中,这些坐标是我们可以轻易在哺乳动物物种中识别的各种本能情绪行动。在情绪感受和意识的分析中,本能行动机制的核心作用传统上被边缘化了。它常被简单地视为神经系统的”输出”,而不是一个复杂的整合过程。正如达尔文所暗示的,情绪表达性行动提供了我们基本情绪本质的连贯图像。我们与世界的最早互动是自发主动的。只需看看任何婴儿、任何儿童、任何年幼的脊椎动物:寻求(SEEKING)位于他们所有抱负的基础。
我们面临一个严峻的选择。我们和其他动物要么是各种内在价值的继承者,代表着我们大脑的情感潜能,要么是可以作为机械部件来研究的无感受僵尸。我们选择哪个选项?我们在神经科学中如何回答这个问题,将决定我们将创造什么类型的知识,也许还有什么样的文化。
让我们不要低估摆在我们面前的科学问题的重要性。意识的所有方面都是作为广泛神经元网络相互作用的结果在动物和人类大脑中出现的。意识没有单一的回路或”中心”,尽管存在关键的汇聚点(Sukhotinsky, et al., 2007)。正如我长期以来所主张的,导水管周围灰质(PAG)可能是大脑中最重要的位置,因为它与高级和低级大脑功能都有丰富的连接。它是我们情感生活的中央车站,对于不同情绪体验的原始整合至关重要。它将触角深入到大脑的低级和高级区域。大脑中的大部分这种”暗能量”不容易用现代脑成像技术可视化(Zhang & Raichle, 2010),但通过正确的任务,可以生成非凡的图像(Mobbs et al., 2009)。
导水管周围灰质及其相关的脑干网络对于构建高级心智至关重要,其中大脑的分布式但专业化的网络模型比高度预定的模块化专业化更现实,正如许多进化心理学家倾向于假设的那样。当我们开始设想无数的神经元和神经网络,以及它们看似无穷无尽的神经化学物质,在多个再入活动循环中相互影响——相互作用并作用于自身——并产生目前几乎无法测量的多样化全局场动态时,我们从一开始就因破译大脑心智(BrainMind)实际如何详细工作这一任务的复杂性而感到谦卑。但如果我们想了解自己,就必须一步一步沿着这条道路前进。理解大脑如何产生原始情绪感受可能是意识研究中最容易解决的——“最简单的”——问题。
意识肯定不是运行中的大脑的单一全局属性。它有着悠久的进化历史,可以追溯到编码对生存至关重要的大脑和身体状态的古老系统。从心理学上讲,那些源自各种内在大脑系统神经动力学的”基因的祖先之声”被体验为原始感受或原始情感状态。我们主要关注其他哺乳动物(以及一些鸟类;见Bernroider and Panksepp, 2011),主要是因为神经解剖学和神经化学的同源性非常显著,允许可信的跨物种概括。
无脊椎动物物种的意识问题是一个更困难的问题,因为神经相似性在减少。但正如我们之前提到的,即使是小龙虾(基本上是大型昆虫)也对人类滥用的药物和其他哺乳动物物种认为有奖励性的药物表现出条件性场所偏好(Huber et al., 2011; Nathaniel et al., 2009, 2010; Panksepp & Huber, 2004)。因此,对于”低等”物种中的这些问题保持开放的心态,并看看预测将我们引向何方是最明智的。但神经科学存在一个核心困境。在心智科学中,我们想要理解大规模过程——“整体”——但神经科学最擅长研究小的离散现象,或”整体”的”部分”。由于这种倾向,我们很容易将两者混淆,产生本体论谬误(mereological fallacies),即部分-整体混淆(Bennett & Hacker, 2003)。目前神经科学正在提供如此多的部分——如此多的大脑机制——但它们在心智中执行什么功能,即”整体”,更难以破译。
科学家想要理解世界,但他们知道他们的技术更擅长研究自然的部分而不是其复合整体。不同的人对这个困境有不同的解决方案。一个常见的方法是专注于相当狭窄的问题(出于纯粹的必要性,这是科学家所青睐的),在这里人们开始越来越清楚地看到树上的每一片叶子,但他们往往会忽视树木和森林。大多数人必然会关注勒内·笛卡尔(1596-1650)在其《方法论》中的第三条科学规则:“在寻求真理时以有序的方式思考,从最简单和最容易理解的事物开始,逐渐地、一步一步地达到更复杂的知识,甚至将那些不一定如此的材料当作有序的来处理。”或者,正如爱因斯坦据说所说的,“简化,但不要超过必要的程度”(强调为新增)。这就是我们在试图理解人类情感意识时所采取的道路。
活着的大脑,连同它们的心智——它们网络级神经生物学功能的无形表现——反映了一种微妙的平衡,目前还知之甚少,存在于大量相互作用的神经回路之间,这些回路在活着的身体中并为其工作,通过创造期望的环境和避免有害的环境来应对世界的挑战。情绪感受是这种相互作用的体验性情感表现;它们是心智的主观品质,其某些方面最终可以在其他生物中系统地、详细地研究。因此,我们可以开始从神经科学角度理解我们自己的心智。如果不研究其他动物的相关过程,就无法实现这种理解。正如生物科学中预示医学进步的其他成功案例一样,理解一直由动物研究的发现引导,每一步都是如此。正如查尔斯·达尔文所认识到的,我们获得的知识将对理解人类状况产生深远的影响。作为一个物种,我们对自己还有很多需要了解的地方。我们还在等什么?
Abbass, A. A., Hancock, J. T., Henderson, J., & Kisely, S. (2006). 短程精神动力学心理治疗用于常见精神障碍。系统综述Cochrane数据库, 4, CD004687。
Adamec, R, (2001). 导水管周围灰质(PAG)的长时程增强是否介导捕食者应激在啮齿动物焦虑样行为(ALB)中产生的持久变化?—PAG的低频刺激(LFS)对位置偏好和捕食者应激产生的ALB变化的影响。行为脑研究, 120(2), 111–135。
Adamec, R., Blundell, J., & Burton, P. (2005). 介导对捕食者应激产生持久脑和行为反应的神经回路变化。神经科学与生物行为评论, 29, 1225–1241。
Adamec, R. E., & Young, B. (2000). 特定边缘系统回路的神经可塑性可能介导特定点燃诱导的动物情感变化——对理解与癫痫相关焦虑的启示。神经科学与生物行为评论, 24, 705–723。
Adkins-Regan, E. (2009). 社会行为的神经内分泌学。实验动物研究所杂志, 50, 5–14。
Adolphs, R., Tranel, D., and Damasio, A. R. (2003). 识别情绪的可分离神经系统。大脑与认知, 52, 61–69。
Agrawal, A., Timothy, J., Pandit, L., & Manju, M. (2006). 创伤后癫痫:概述。临床神经学与神经外科学, 108, 433–439。
Ahern, T. H., & Young, L. J. (2009). 早期生活家庭结构对单配偶草原田鼠(Microtus ochrogaster)成年社会依恋、异亲照料行为以及调节亲和行为的神经肽系统的影响。行为神经科学前沿, 3, 17。
Aichhorn, A. (1925). 误入歧途的青少年。伦敦:Imago出版社。
Ainsworth, M. D. (1982). 依恋:回顾与展望。见C. M. Parkes和J. Stevenson-Hinde主编,依恋在人类行为中的地位。伦敦:Tavistock。
Ainsworth, M. D., & Boston, M. (1952). 幼儿期长期分离后儿童的心理诊断评估。英国医学心理学杂志, 25, 169–201。
Airan, R. D., Thompson, K. R., Fenno, L. E., Bernstein, H., & Deisseroth, K. (2009). 体内时间精确控制细胞内信号传导。自然, 458, 1025–1029。
Al-Azzawi, F., Bitzer, J., Brandenburg, U., Castelo-Branco, C., Graziottin, A., Kenemans, P. & Zahradnik, H. P. (2010). 绝经后女性性功能障碍的治疗选择。更年期, 13, 103–120。
Alcaro, A., Huber, R., & Panksepp, J. (2007). 中脑边缘多巴胺系统的行为功能:情感神经行为学视角。脑研究评论, 56, 283–321。
Alcaro, A., Panksepp, J., Witczak, J., Hayes, D. J., & Northoff, G. (2010). 抑郁症中的皮层下-皮层中线活动是否由谷氨酸和GABA介导?跨物种转化方法。神经科学与生物行为评论, 34(4), 592–605。
Aldis, O. (1975). 玩耍性打斗。纽约,纽约州:学术出版社。
Alexander, M., & Perachio, A.A. (1973). 目标性别和支配地位对恒河猴诱发攻击的影响。美国体质人类学杂志, 38, 543–547。
Amano, K., Notani, M., Iseki, H., Kawabatake, H., Tanikawa, T., Kawamura, H., & Kitamura, K. (1979). 人类脑室周围灰质电刺激前后第三脑室脑脊液的高香草酸浓度。见E. R. Hitchcock、H. T. Ballantine, Jr.和B. A. Meyerson主编,精神外科的现代概念(第65–76页)。阿姆斯特丹:Elsevier。
Amaral, D. G., Price, J. L., Pitkanen, A., & Carmichael, T. (1992). 灵长类杏仁核复合体的解剖组织。见J. Aggleton主编,杏仁核:情绪、记忆和精神功能障碍的神经生物学方面(第1–66页)。纽约,纽约州:Wiley-Liss。
Amaral, O. B., & Roesler, R. (2008). 针对恐惧相关障碍的NMDA受体靶向。中枢神经系统药物发现最新专利, 3, 166–178。
American College of Radiology. (2004). ACR学习档案:神经放射学,第2版;ACR:弗吉尼亚州雷斯顿,美国。
Antelman, S. M., Szechtman, H., Chin, P., & Fisher, A. E. (1975). 捏尾诱导的大鼠进食、啃咬和舔舐行为:对黑质纹状体多巴胺系统的依赖。脑研究, 99(975), 319–337。
Austin, J. H. (1998). 禅与脑。马萨诸塞州剑桥:MIT出版社。
Austin, J. (2006). 禅脑反思。马萨诸塞州剑桥:MIT出版社。
Avena, N. M., Rada, P., & Hoebel, B. G. (2008). 糖成瘾的证据:间歇性过量摄入糖的行为和神经化学效应。神经科学与生物行为评论, 32, 20–39。
Averill, J. R. (2010). 关于愤怒的十个你可能从未想过要问的问题。见F. Pahlavan主编,愤怒的多重面向:发火还是恢复正义?(第1–25页)。纽约,纽约州:Nova科学出版社。
Baddeley, A. D., & Hitch, G. J. (1974). 工作记忆(Working memory)。见G. A. Bower主编,学习与动机心理学:研究与理论进展(第8卷,第47–89页)。纽约,纽约州:学术出版社。
Bagemihl, B. (1999). 生物的丰盛:动物同性恋与自然多样性。纽约,纽约州:圣马丁出版社。
Bailey, P., & Davis, E. W. (1942). 猫导水管周围灰质损伤的影响。实验生物学与医学学会会议录, 351, 305–306。
Bailey, P., and Davis, E. W. (1943) 导水管周围灰质损伤对猕猴的影响。神经病理学与实验神经学杂志, 3, 69–72。
Balthazart, J., Tlemçani O., & Ball, G. F. (1996). 大脑的性别差异能否解释激素诱导生殖行为的性别差异?对日本鹌鹑25年的研究告诉我们什么,激素与行为, 30, 627–661。
Bandler, R. (1988). 电刺激和化学刺激揭示的攻击性脑机制:提示中脑导水管周围灰质区域的中心作用。见 A. N. Epstein & A. R. Morrison (主编),《心理生物学和生理心理学进展》(第13卷,第67-154页)。圣地亚哥:学术出版社。
Barbour, J. (2000). 《时间的终结》。纽约:牛津大学出版社。
Barrett, L. F. (2006). 情绪是自然类别吗?《心理科学观点》,1,28-58。
Barrett, P., & Bateson, P. (1978). 猫游戏行为的发展。《行为学》,66,105-120。
Bartz, J. A., & Hollander, E. (2008). 催产素(Oxytocin)与自闭症谱系障碍的实验性治疗。《脑研究进展》,170,451-462。
Bartz, J., Simeon, D., Hamilton, H., Kim, S., Crystal, S., Braun, A., Vicens, V., & Hollander, E. (2010, 11月29日). 催产素(Oxytocin)可能阻碍边缘型人格障碍患者的信任与合作。《社会认知与情感神经科学》。[网络预先发表]。
Bauby, J-D. (1997). 《潜水钟与蝴蝶》。纽约:AA Knopf出版社。
Beatty, W. W., Costello, K. B., & Berry, S. L. (1984). 苯丙胺对游戏性打斗的抑制:儿茶酚胺拮抗剂、激动剂和合成抑制剂的效果。《药理学、生物化学与行为》,20,747-755。
Beatty, W. W., Dodge, A. M., Dodge, L. J., Whike, K., & Panksepp, J. (1982). 精神运动兴奋剂、社会剥夺与幼年大鼠的游戏。《药理学、生物化学与行为》,16,417-422。
Beebe, B., & Lachmann, E. M. (1988). 母婴相互影响对自我和客体关系起源的贡献。《精神分析心理学》,5,305-337。
Bejjani, B. P., Damier, P., Arnulf, I., Thivard, L., Bonnet, A. M., Dormont, D., Cornu, P., Pidoux, B., Samson, Y., & Agid, Y. (1999). 高频深部脑刺激诱发的短暂性急性抑郁。《新英格兰医学杂志》,340,1476-1480。
Bekkedal, M. Y., Rossi, J., 3rd, & Panksepp, J. (2011). 情绪的人脑脑电图指标:通过测量额叶θ事件相关同步化来描绘对情感发声的反应。《神经科学与生物行为评论》,35(9),1959-1970。
Bekoff, M. (2000). 《海豚的微笑:动物情感的非凡记录》。纽约:兰登书屋/探索图书。
Bekoff, M. (2007). 《动物的情感生活》。诺瓦托,加州:新世界图书馆。
Belenky, G., Martin, J. A., & Marcy, S. C. (1996). “战斗后的行动后关键事件压力汇报和战斗重建。”见 J. Martin, L. Sparacino, & G. Belenky (主编),《海湾战争与心理健康:综合指南》(第105-114页)。韦斯特波特,康涅狄格州:Praeger出版社。
Benedek, T., & Rubenstein, B. B. (1942). 《女性的性周期》。华盛顿特区:国家研究委员会。
Bennett, M. R., & Hacker, P. M. S. (2003). 《神经科学的哲学基础》。马尔登,马萨诸塞州:Blackwell出版社。
Bentham, J. (1779/1879). 《道德与立法原理导论》。牛津:克拉伦登出版社。
Berlin, H. A. (2007). 抗癫痫药物治疗创伤后应激障碍。《当代精神病学报告》,9,291-300。
Berlin, H. A., Rolls, E. T., & Kischka, U. (2004). 眶额皮质损伤患者的冲动性、时间知觉、情绪和强化敏感性。《脑》,127,1108-1126。
Bernatzky, G., Presch, M., Anderson, M., & Panksepp, J. (2011). 音乐作为现代医学非药物疼痛管理工具的情感基础。《神经科学与生物行为评论》,35,1989-1999。
Bernroider, G., & Panksepp, J. (2011, 3月3日). 镜子与感受:你看到外面的演员了吗?《神经科学与生物行为评论》,35,2009-2016。
Berridge, K. C. (1996). 食物奖励:想要和喜欢的脑基质。《神经科学与生物行为评论》,20,1-26。
Berridge, K. C. (2000). 测量动物和婴儿的享乐冲击:情感味觉反应模式的微观结构。《神经科学与生物行为评论》,24,173-198。
Berridge, K. C. (2004). 快乐、无感情感和非理性欲望。见 A. S. R. Manstead, N. Frijda, & A. Fischer (主编),《感受与情绪:阿姆斯特丹研讨会》(第243-262页)。剑桥,英国:剑桥大学出版社。
Berridge, K. C., & Robinson, T. E. (1998). 多巴胺(Dopamine)在奖励中的作用是什么:享乐冲击、奖励学习还是激励显著性?《脑研究评论》,28,309-369。
Berridge, K. C., Robinson, T. E., & Aldridge, J. W. (2009). 解剖奖励的组成部分:“喜欢”、“想要”和学习。《药理学当前观点》,9,65-73。
Berridge, K. C., & Valenstein, E. S. (1991). 什么心理过程介导外侧下丘脑电刺激诱发的进食?《行为神经科学》,105,2-14。
Berta, P, Hawkins, J. R., Sinclair, A. H., Taylor, A., Griffiths, B. L. & Goodfellow, P. N. (1990). 等同SRY和睾丸决定因子的遗传证据。《自然》,348,488-450。
Birbaumer, N. (2006). 打破沉默:用于沟通和运动控制的脑机接口(BCI)。《心理生理学》,43,517-532。
Black, D. (1982). 病理性笑:文献综述。《神经与精神疾病杂志》,170,67-71。
Blackburn, J. R., Pfaus, J. G., & Phillips, A. G. (1992). 多巴胺(Dopamine)在食欲和防御行为中的功能。《神经生物学进展》,39,247-279。
Blair, R. J., Morris, J. S., Frith, C. D., Perrett, D. I., & Dolan, R. J. (1999) 对悲伤和愤怒面部表情的可分离神经反应。《脑》,122,883-893。
Blakemore, S. J. (2003). 欺骗运动系统。《意识与认知》,12,647-655。
Blakemore, S.-J., Wolpert, D., & Frith, C.D. (2000). 为什么你不能给自己挠痒痒?《神经报告》,11,R11-R16。
Bliss, T., & Lomo, T. (1973). 在未麻醉兔子穿通路刺激后海马齿状区突触传递的长时程增强。生理学杂志, 232, 357–454。
Blood, A. J., & Zatorre, R. J. (2001). 对音乐的强烈愉悦反应与大脑奖赏和情绪相关区域的活动相关。美国国家科学院院刊, 98, 11818–11823。
Blumer, D. (2000). 情绪障碍和阵发性情感:癫痫相关精神障碍的识别和治疗。哈佛精神病学评论, 8, 8–17。
Bodkin, J. A., Zornberg, G. L., Lukas, S. E, & Cole, J. O. (1995). 丁丙诺啡治疗难治性抑郁症。临床精神药理学杂志, 1, 49–57。
Bohus, M. J., Lanwhremeyer, G. B., Stiglmayr, C. E., Limberger, M. F., Bohme, R., & Schmahl, C. G. (1999). 纳曲酮治疗边缘型人格障碍患者的分离症状:一项开放标签试验。临床精神病学杂志, 60, 598–603。
Boldrini, M., Underwood, M. D., Hen, R., Rosoklija, G. B., Dwork, A. J., John Mann, J., & Arango, V. (2009). 抗抑郁药增加人类海马的神经祖细胞。神经精神药理学, 34, 2376–2389。
Bolwerk, E. L. M., & Swanson, H. H. (1984). 催产素在大鼠母性行为开始中起作用吗?内分泌学杂志, 101, 353–357。
Borod, J. C. (主编) (2000). 情绪的神经心理学。纽约,NY:牛津大学出版社。
Bos, P. A., Hermans, E. J. & van Honk, J. (2010). 睾酮降低社交天真人类的信任。美国国家科学院院刊, 107, 9991–9995。
Bos, P. A., Panksepp, J., Bluthé, R. M, & Honk, J. V. (2011). 类固醇激素和神经肽对人类社会情绪行为的急性影响:单次给药研究综述。神经内分泌学前沿。出版中。
Bouvard, M. P., Leboyer, M., Launay, J.-M., Racasens, C., Plumet, M.-H., Waller-Perotte, D., Tabuteau, F., Bondoux, D., Dugas, M., Lensing, P., & Panksepp, J. (1995). 低剂量纳曲酮对自闭症患者血浆化学和临床症状的影响:一项双盲安慰剂对照研究。精神病学研究, 58, 191–201。
Bowlby, J. (1953). 儿童照护与爱的成长。伦敦:企鹅图书。
Bowlby, J. (1960). 分离焦虑。国际精神分析杂志, 41, 89–113。
Bowlby, J. (1980). 依恋与丧失,第3卷。丧失:悲伤与抑郁。纽约,NY:基础图书。
Brammer, R. (2011). 咨询中的多样性:探索种族和性别问题(第2版)。太平洋丛林,CA:Brooks/Cole出版社。
Brandão, M. L., Borelli, K. G., Nobre, M. J., Santos, J. M., Albrechet-Souza, L., Oliveira, A. R., & Martinez, R. C. (2005). 中脑顶盖恐惧神经组织的GABA能调节。神经科学与生物行为评论, 29, 1299–1311。
Brandão, M. L., Tronscoso, A. C., Silva, M. A. S., & Huston, J. P. (2003). 中脑顶盖防御神经基质与焦虑和压力的相关性:实证和概念性考虑。欧洲药理学杂志, 463, 225–233。
Brandão, M. L., Zanoveli, J. M., Ruiz-Martinez, R. C., Oliveira, L. C., & Landeira-Fernandez, J. (2008). 大鼠导水管周围灰质组织的不同僵直行为模式:与不同类型焦虑的关联。行为脑研究, 188, 1–13。
Brayley, K. N., & Albert, D. J. (1977). 通过刺激外侧隔核抑制VMH损伤诱导的大鼠反应性和攻击性,而非内侧隔核或扣带皮层。比较与生理心理学杂志, 91, 290–299。
Breedlove, S. M. (1992). 大脑和行为的性分化。见 J. B. Becker, S. M. Breedlove, & D. Crews (主编),行为内分泌学。剑桥,MA:麻省理工学院出版社。
Brown, P. L., & Jenkins, H. M. (1968). 鸽子啄键的自动塑造。实验行为分析杂志, 11, 1–8。
Browning, J. R., Browning, D. A., Maxwell, A. O., Dong, Y., Jansen, H. T., Panksepp, J., & Sorg, B. A. (2011). 可卡因和蔗糖自我给药期间的积极情感发声:大鼠自发药物渴望模型,神经药理学, 61, 268-275。
Bruchas, M. R., Land. B. B., & Chavkin, C. (2010). 强啡肽/κ阿片受体系统作为压力诱导和促成瘾行为的调节器。脑研究, 1314, 44–55。
Brudzynski, S. M. (2007). 大鼠超声波叫声作为负性或正性状态的指示变量:乙酰胆碱-多巴胺相互作用和声学编码。行为脑研究, 182(2), 261–273。
Brudzynski, S. M. (主编). (2010). 哺乳动物发声手册。牛津,英国:学术出版社。
Brudzynski, S. M, & Holland, G. (2005). 直接记录中气流诱导的22-kHz警报叫声的声学特征。神经科学与生物行为评论, 29, 1169–1180。
Brudzynski, S. M., Silkstone, M., Komadoski, M., Skullion, K., Duffus, S., Burgdorf, J., Kroes, R. A., Moskal, J. R., & Panksepp, J. (2010). 伏隔核内苯丙胺对三个选择性繁育Long-Evans大鼠品系50-kHz发声产生的影响。行为脑研究, 217(1), 32–40。
Buck, R. (1999). 生物性情感:一个类型学。心理学评论, 106(2), 301–336。
Burgdorf, J., Knutson, B., & Panksepp, J. (2000). 对奖赏性电脑刺激的期待引发大鼠超声波发声。行为神经科学, 114, 320–327。
Burgdorf, J., Knutson, B., Panksepp, J., & Ikemoto, S. (2001). 伏隔核苯丙胺微注射无条件地引发大鼠50-kHz超声波发声。精神药理学, 155, 35–42。
Burgdorf, J., Kroes, R. A., Beinfeld, M. C., Panksepp, J., & Moskal, J. R. (2010). 揭示积极情绪的分子基础:大鼠粗暴游戏中胰岛素样生长因子I的作用。Neuroscience, 163, 769–777.
Burgdorf, J., Kroes, R. A., Moksal, J. R., Pfaus, J. G., Brudzynski, S. M., & Panksepp, J. (2008). 大鼠(rattus norvegicus)在交配、游戏和攻击过程中的超声波发声:行为伴随、与奖励的关系以及回放的自我给予。Journal of Comparative Psychology, 122, 357–367.
Burgdorf, J., & Panksepp, J. (2006). 积极情绪的神经生物学。Neuroscience and Biobehavioral Reviews, 30, 173–187.
Burgdorf, J., Panksepp, J., Beinfeld, M. C., Kroes, R. A., & Moskal, J. R. (2006). 青春期大鼠粗暴游戏行为中区域脑胆囊收缩素的变化。Peptides, 27, 172–177.
Burgdorf, J., Panksepp, J., Brudzynski, S. M., Beinfeld, M. C., Cromwell, H. C., Kroes, R. A., & Moskal, J. R. (2009). 选择性繁育不同50-kHz超声波发声率对大鼠情绪行为的影响。Developmental Psychobiology, 51, 34–46.
Burgdorf, J., Panksepp, J., Brudzynski, S. M., & Moskal, J. R. (2005). 繁育具有50-kHz积极情感发声的大鼠。Behavior Genetics, 35, 67–72.
Burgdorf, J., Panksepp, J., & Moskal, J. R. (2011). 频率调制的50kHz超声波发声:揭示积极情感分子基础的工具。Neuroscience & Biobehavioral Reviews, 35, 1831–1836.
Burgdorf, J., Wood, P. L., Kroes, R. A., Moskal, J. R., & Panksepp, J. (2007). 大鼠50-kHz超声波发声的神经生物学:电极定位、损伤和药理学研究。Behavioral Brain Research, 182(2), 274–283.
Burghardt, G. M. (2005). 动物游戏的起源:测试极限。剑桥,马萨诸塞州:麻省理工学院出版社。
Busnel, M. C., Granier-Deferre, C., & Lecanuet, J. P. (1992). 胎儿听觉。Annals of the New York Academy of Sciences, 662, 118–134.
Byers, J. A. (1997). 美洲叉角羚:社会适应与过去捕食者的幽灵。芝加哥:芝加哥大学出版社。
Byers, J. A., and Walker, C. B. (1995). 完善游戏进化的运动训练假说。American Naturalist, 146, 25–40.
Cabanac, M. (1992). 愉悦:共同的货币。Journal of Theoretical Biology, 155, 173–200.
Cacioppo, J., & Patrick, W. (2008). 孤独:人性与社交联系的需求。纽约:W. W. Norton出版社。
Caggiula, A. R. (1970). 雄性大鼠后下丘脑刺激的交配奖励特性分析。Journal of Comparative and Physiological Psychology, 70, 399–412.
Caggiula, A. R., & Eibergen, R. (1969). 疼痛性外周刺激引发的处女雄性大鼠交配行为。Journal of Comparative and Physiological Psychology, 69, 414–419.
Calder, A. J., Keane, J., Lawrence, A. D., & Manes, F. (2004). 腹侧纹状体损伤后愤怒识别受损。Brain, 127, 1958–1969.
Caldwell, J. D. (2002). 涉及质膜类固醇效应的性唤起模型。Neuroscience and Biobehavioral Reviews, 26, 13–30.
Campeau, S., & Davis, M. (1995). 皮层下和皮层传入杏仁核外侧核在恐惧条件反射中的参与:通过恐惧增强惊吓反应测量同时接受听觉和视觉条件刺激训练的大鼠。Journal of Neuroscience, 15, 2312–2327.
Cannon, W. B. (1927). 詹姆斯-兰格情绪理论:批判性检验与替代理论。American Journal of Psychology, 39, 106–124.
Carter, C. S. (1998). 社会依恋与爱的神经内分泌视角。Psychoneuroendocrinology, 23, 779–818.
Carter, C. S. (2007). 催产素和加压素的性别差异:对自闭症谱系障碍的影响?Behavioural Brain Research, 176, 170–186.
Carter, S. C., DeVries, A. C., & Gets, L. L. (1995). 哺乳动物一夫一妻制的生理基础:草原田鼠模型。Neuroscience and Biobehavioral Reviews, 19, 303–314.
Castellanos, F. X., & Tannock, R. (2002). 注意力缺陷/多动障碍的神经科学:寻找内表型。Nature Reviews Neuroscience, 3, 617–628.
Chalmove, A. S. (1978). 孤立恒河猴的治疗:不同伙伴与社会行为。Child Development, 49, 43–50.
Champagne, F., Francis, D. D., Mar, A., & Meaney, M. J. (2003). 大鼠母性照料的变异作为环境对发育影响的中介因素。Physiology & Behavior, 79, 359–371.
Chase, T. N., & Friedhoff, A. J. (主编). (1982). 抽动秽语综合征。纽约,纽约州:Raven出版社。
Chen, Q., Panksepp, J. B., & Lahvis, G. P. (2009). 共情受小鼠遗传背景调节。PlosOne, 4, e4387.
Chen, R. C., & Forster, F. M. (1973). 跑动性癫痫与发笑性癫痫。Neurology, 23, 1019–1029.
Chiron, C., Jambaque, I., Nabbout, R., Lounes, R., Syrota, A., & Dulac, O. (1997). 右脑半球在人类婴儿中占主导地位。Brain, 120, 1057–1065.
Chomsky, N. (1968). 语言与心智。纽约,纽约州:Harcourt, Brace & Jovanovich出版社。
Chugani, H. T. (1996). 发育非线性和发育病理学的神经影像学。见 R. W. Thatcher, G. Reid Lyon, J. Rumsey, & N. Krasnegor (主编), 发育神经影像学:绘制大脑和行为的发育图谱 (第187–198页)。圣地亚哥:学术出版社。
Chugani, H. T. (1998). 大脑发育的关键期:用PET研究脑葡萄糖利用。Preventive Medicine, 27, 184–188.
Cibrian-Llanderal T., Tecamachaltzi-Silvaran, M., Triana-Del Rio, R., Pfaus, J. G., Manzo, J., & Coria-Avila, G. A. (2010). 阴蒂刺激调节食欲性行为并促进大鼠繁殖。生理学与行为(Physiology & Behavior), 100, 148–153.
Ciocchi, S., Herry, C., Grenier, F., Wolff, S. B., Letzkus, J. J., Vlachos, I. Ehrlich, I., Sprengel, R., Deisseroth, K., Stadler, M. B., Müller, C., & Lüthi. A. (2010). 中央杏仁核抑制回路中条件性恐惧的编码。自然(Nature), 468, 277–282.
Cirulli, F., Francia, N., Berry, A., Aloe, L., Alleva, E., & Suomi, S. J. (2009). 早期生活压力作为心理健康的风险因素:从啮齿类动物到非人灵长类动物的神经营养因子作用。神经科学与生物行为评论(Neuroscience and Biobehavioral Reviews), 33, 573–585.
Clancy, S. (2009). 创伤神话:儿童性虐待的真相及其后果(The trauma myth: The truth about the sexual abuse of children—and its aftermath). 纽约:基础图书出版社。
Clarke, S., & Trowill, J. A. (1971). 大鼠的嗅探与动机行为。生理学与行为(Physiology & Behavior), 6, 49–52.
Clayton, N. S., Bussey, J. T., & Dickenson, A. (2003). 动物能回忆过去并计划未来吗?自然神经科学评论(Nature Reviews Neuroscience), 4, 685–691.
Clayton, N.S., & Russell, J. (2009). 在动物和幼儿中寻找情景记忆:新极简主义的前景。神经心理学(Neuropsychologia), 47, 2330–2340.
Clynes, M. (1977). 感知学:情感的触碰(Sentics: The touch of emotions). 纽约:双日出版社。
Code, D. (2009). 要培养快乐的孩子,先把婚姻放在第一位(To raise happy kids, put your marriage first). 纽约:十字路出版社。
Coenen, V. A., Schlaepfer, T. E., Maedler, B., & Panksepp, J. (2011). 内侧前脑束的跨物种情感功能——对人类情感性疼痛和抑郁治疗的启示。神经科学与生物行为评论(Neuroscience & Biobehavioral Reviews), 35, 1971–1981.
Colonnello, V., Iacobucci, P., Fuchs, T., Newberry, R. C., & Panksepp, J. (2011). 八齿鼠(Octodon degus):社会情感神经科学研究的有用动物模型:分离痛苦、社会依恋和玩耍的基本描述。神经科学与生物行为评论(Neuroscience & Biobehavioral Reviews), 35, 1854–1863.
Comings, D. E., Comings, B. G., Muhleman, D., Dietz, G., Shahbahrami, B., Tast, D. & Flanagan, S. D. (1991). 多巴胺D2受体基因座作为神经精神疾病的修饰基因。美国医学会杂志(Journal of the American Medical Association), 266, 1793–1800.
Conrad, C. D. (2008). 慢性压力诱导的海马脆弱性:糖皮质激素脆弱性假说。神经科学评论(Reviews of Neuroscience), 19, 395–411.
Conway, A. R. A., Kane, M. J., & Engle, R. W. (2003). 工作记忆容量及其与一般智力的关系。认知科学趋势(Trends in Cognitive Sciences), 7, 547–552.
Copleston, F. (1962a). 哲学史(A History of Philosophy). 第1卷:希腊与罗马(Greece and Rome), 第一部分. 花园城:形象图书出版社。
Copleston, F. (1962b). 哲学史(A History of Philosophy). 第4卷:现代哲学:从笛卡尔到莱布尼茨(Modern Philosophy: Descartes to Leibniz). 花园城:形象图书出版社。
Cousins, N. (1983). 治愈之心:恐慌和无助的解药(The healing heart: Antidotes to panic and helplessness). 纽约:W. W. 诺顿出版社。
Cozolino, L. (2002). 心理治疗的神经科学:构建和重建人类大脑(The neuroscience of psychotherapy: Building and rebuilding the human brain). 纽约:W. W. 诺顿出版社。
Cozolino, L. (2010). 心理治疗的神经科学:治愈社会脑(The neuroscience of psychotherapy: Healing the Social Brain). 第2版. 纽约:W. W. 诺顿出版社。
Craig, A. D. (2002). 你感觉如何?内感受:身体生理状态的感知。自然神经科学评论(Nature Reviews Neuroscience), 3, 655–666.
Craig, A. D. (2003a). 内感受:身体生理状态的感知。神经生物学当前观点(Current Opinion in Neurobiology), 13, 500–505.
Craig, A. D. (2003b). 疼痛机制:标记线与中枢处理中的汇聚。神经科学年度评论(Annual Review of Neuroscience), 26, 1–30.
Craig AD (2009). 你现在感觉如何?前岛叶与人类意识。自然神经科学评论(Nature Reviews Neuroscience), 10, 59–70.
Crawley, J. N. (2007). 我的小鼠怎么了?转基因和基因敲除小鼠的行为表型分析(What’s Wrong With My Mouse? Behavioral Phenotyping of Transgenic and Knockout Mice) (第2版). 纽约:威利父子出版社。
Cromwell, H.C. & Panksepp, J. (2011). 从神经视角重新思考认知革命:行为神经科学中”认知”一词的过度使用/误用以及对情感控制的忽视如何可能延缓对脑心(BrainMind)理解的进展,神经科学与生物行为评论(Neuroscience and Biobehavioral Reviews), 35, 2026–2035.
Dahan, L., Astier, B., Vautrelle, N., Urbain, N., Kocsis, B., & Chouvet, G. (2007). 矛盾睡眠期间腹侧被盖区多巴胺能神经元的显著爆发式放电。神经精神药理学(Neuropsychopharmacology), 32, 1232–1241.
Daitzman, R., & Zuckerman, M. (1980). 去抑制感觉寻求、人格与性腺激素。人格与个体差异(Personality and Individual Differences), 1, 103–110.
Daly, M., & Wilson, M. (2001). 冒险、同性竞争与杀人。内布拉斯加动机研讨会(Nebraska Symposium on Motivation), 47, 1–36.
Damasio, A. R. (1994). 笛卡尔的错误:情感、理性与人类大脑(Descartes’ error: Emotion, reason, and the human brain). 纽约:雅芳图书出版社。
Damasio, A. R. (1999). 发生的感觉:身体与情感在意识构建中的作用(The feeling of what happens: Body and emotion in the making of consciousness). 纽约:哈考特·布雷斯出版社。
Damasio, A. R. (2003). 寻找斯宾诺莎:快乐、悲伤与感知大脑(Looking for Spinoza: Joy, sorrow and the feeling brain). 奥兰多:哈考特出版社。
Damasio, A. R. (2010). 自我进入心智:构建意识大脑(Self comes to mind: Constructing the conscious brain). 纽约:万神殿图书出版社。
Damasio, A. R., Grabowski, T. J., Bechara, A., Damasio, H., Ponto, L. L. B., Parvizi, J., & Hichwa, R. D. (2000). 自我产生情感的感受期间的皮层下和皮层大脑活动。自然神经科学(Nature Neuroscience), 3, 1049–1056.
Damoiseaux, J. S., Rombouts, S. A., Barkhof, F., Scheltens, P., Stam, C. J., Smith, S. M., & Beckmann, C. F. (2006). 健康受试者的一致静息状态网络。美国国家科学院院刊(Proceedings of the National Academy of Sciences U.S.A.), 103, 13848–13853.
Dantzer, R., Bluthe, R. M., Koob, G. F., &Le Moal, M. (1987). 神经垂体肽对雄性大鼠社会记忆的调节。精神药理学(Psychopharmacology), 91, 363–368.
Dantzer, R., Koob, G. F., Bluthe, R. M., & Le Moal, M. (1988). 隔膜加压素调节雄性大鼠的社交记忆。Brain Research, 457, 143–147.
Dantzer, R., O’Connor, J. C., Freund, G. G., Johnson, R. W., & Kelley, K. W. (2008). 从炎症到疾病和抑郁:当免疫系统征服大脑时。Nature Reviews Neuroscience, 6, 46–57.
Darwin, C. (1871). 人类的由来及性选择。Princeton, NJ: Princeton University Press.
Darwin, C. (1872). 人类和动物的情绪表达。London, UK: John Murray.
Darwin, C. (1872/1998). 人类和动物的情绪表达(第3版)。New York, NY: Oxford University Press.
Daston, L., & Mitman, G. (2005). 与动物一起思考:拟人论的新视角。New York, NY: Columbia University Press.
Davanloo, H. (2005). 强化短程动力心理治疗。见 H. Kaplan 和 B. Sadock(编),精神病学综合教科书(第8版,第2卷,第2628–2652页)。Philadelphia: Lippincott Williams & Wilkins.
Davidson, R. J. (2004). 幸福感和情感风格:神经基础和生物行为相关性。Philosophical Transactions of the Royal Society of London, B, 359, 1395–1411.
Davidson, R. J., Scherer, K. R., & Goldsmith, H. H. (2003). 情感科学手册。New York, NY: Oxford University Press.
Davies, P. S. (2011). 哺乳动物思维中的祖先之声:Jaak Panksepp情感神经科学的哲学意义。Neuroscience and Biobehavioral Reviews, 35, 2036–2044.
Davis, K. L., Panksepp, J., & Normansell, L. (2003). 情感神经科学人格量表:常模数据和意义。NeuroPsychoanalysis, 5, 21–29.
Davis, K. L., & Panksepp, J. (2011). 人类人格的大脑情感基础和情感神经科学人格量表。Neuroscience and Biobehavioral Reviews, 35, 1946–1958.
Davis, M. (1992). 杏仁核在恐惧和焦虑中的作用。Annual Reviews Neuroscience, 15, 353–375.
Davis, M., Campeau, S., Kim, M., & Falls, W. A. (1995). 情绪的神经系统:杏仁核在恐惧和焦虑中的作用。见 J. L. McGaugh, N. M. Weinberger, & G. Lynch(编),大脑与记忆:神经可塑性的调节和中介(第3–40页)。New York: Oxford University Press.
Davis, M., & Lang, P. J. (2003). 情绪。见 M. Gallagher, R. J. Nelson, & I. B. Weiner(编),心理学手册:第3卷 生物心理学(第405–440页)。Hoboken, NJ: John Wiley & Sons.
Davis, M., Ressler, K., Rothbaum, B. O., & Richardson R. (2006). D-环丝氨酸对消退的影响:从临床前到临床工作的转化。Biological Psychiatry, 60, 369–375.
Davis, M., Walker, D. L., Miles, L., & Grillon, C. (2010). 大鼠和人类的阶段性恐惧与持续性恐惧:扩展杏仁核在恐惧与焦虑中的作用。Neuropsychopharmacology Reviews, 35, 105–135.
Decety, J., & Ickes, W.(编)(2009). 共情的社会神经科学。Cambridge, MA: MIT Press.
De Dreu, C. K., Greer, L. L., Handgraaf, M. J., Shalvi, S., Van Kleef, G. A., Baas, M., Ten Velden, F. S., Van Dijk, E., & Feith, S. W. (2010). 神经肽催产素调节人类群体间冲突中的狭隘利他主义。Science, 328, 1408–1411.
de Jong, T. R., Chauke, M., Harris, B. N., & Saltzman, W. (2009). 从这里到父亲身份:加州小鼠(Peromyscus californicus)父性行为开始的神经相关性。Hormones & Behavior, 56, 220–231.
Del-Ben, C. M., & Graeff, F. G. (2009). 恐慌障碍:PAG参与其中吗?Neural Plasticity, 1–9. doi:10.1155/2009/108135.
Delgado, J. M. R. (1969). 心智的物理控制:迈向心理文明社会。New York, NY: Harper and Row.
Delgado, J. M. R., Roberts, W. W., & Miller, N. E. (1954). 通过大脑电刺激激发的学习。American Journal of Physiology, 179, 587–593.
DeMolina A. F., & Hunsperger, R. W. (1962). 猫的皮层下系统中支配防御和逃跑反应的组织结构。Journal of Physiology (London), 160, 200–213.
Denton, D. (2006). 原始情绪:意识的黎明。New York, NY: Oxford University Press.
Descartes, R. (1960). 方法论(L. Lafleur 译)。New York; Macmillan.
De Vries, G. J., & Panzica, G. C. (2006). 脊椎动物中枢加压素和加压素系统的性别分化:不同机制,相似结果。Neuroscience, 138, 947–55.
de Waal, F. (2009). 灵长类动物与哲学家:道德如何进化。Princeton, NJ: Princeton University Press.
Diamond, M. (2004). 多年来的性、性别和身份:不断变化的视角。Child and Adolescent Psychiatric Clinics of North America, 13, 591–607.
Dissanayake, E. (2003). 艺术的核心:制造特殊。Journal of the Canadian Association for Curriculum Studies, 1, 13–38.
Ditzen, B., Schaer, M., Gabriel, B., Bodenmann, G., Ehlert, U., & Heinrichs, M. (2009). 鼻内催产素增加积极沟通并降低夫妻冲突期间的皮质醇水平。Biological Psychiatry, 65, 728–731.
Doidge, N. (2007). 重塑大脑的自己。New York: Penguin.
Donaldson, Z. R., & Young, L. J. (2008). 催产素、加压素和社交性的神经遗传学。Science, 322, 900–904.
Doron, N. N., & LeDoux, J. E. (1999). 大鼠丘脑听觉和视觉区域到外侧杏仁核投射的组织结构。Journal of Comparative Neurology, 412, 383–409.
Droit-Volet, S., & Meck, W. H. (2007). 情绪如何给我们的时间感知着色。Trends in Cognitive Sciences, 11, 504–513.
Ehrhardt, A. A., Meyer-Bahlburg, H. F. L., Rosen, L. R., Feldman, J. F., Veridiano, N. P., Zimmerman, I., & McEwen, B. S. (1985). 产前暴露于外源性雌激素后的性取向。性行为档案, 14, 57–78.
Ehrlich, I., Humeau, Y., Grenier, F., Ciocchi, S., Herry, C., & Lüthi, A. (2009). 杏仁核抑制回路与恐惧记忆的控制。神经元, 62, 757–771.
Ehrsson, H. H., Spence, C., & Passingham, R. E. (2004). 那是我的手!前运动皮层的活动反映了对肢体的所有权感。科学, 305, 875–877.
Eibl-Eibesfeldt, I. (1989). 人类行为学. 纽约,纽约州:Aldine de Gruyter出版社.
Eickhoff, F. W. (2006). 关于延后性(Nachträglichkeit):一个古老概念的现代性。国际精神分析杂志, 87(第6部分), 1453–1469.
Einon, D., & Potegal, M. (1991). 幼年时期被剥夺玩耍打闹经验的成年大鼠表现出增强的防御行为。攻击性行为, 17, 27–40.
Eisenberger, N. (2010). 社交疼痛的神经基础:发现与启示。见 G. MacDonald & L.A. Jensen-Campbell (编), 社交疼痛:失去和排斥的神经心理学与健康影响 (第53–78页). 华盛顿特区:美国心理学会.
Ekman, P. (1994). 面部表情普遍性的有力证据:对Russell错误批评的回应。心理学通报, 115, 263–287.
Ekman, P., & Davidson, R. J. (编). (1994). 情绪的本质:基本问题. 纽约,纽约州:牛津大学出版社.
Elbert, T., & Rockstroh, B. (2004). 人类大脑皮层的重组:使用和损伤后变化的范围。神经科学家, 10, 129–141.
Eleftheriou, B. E. (编). (1972). 杏仁核的神经生物学. 纽约,纽约州:Plenum出版社.
Elliot, A. (编). (2008). 趋近与回避动机手册. 纽约,纽约州:Taylor & Francis出版社.
Elliott, R., Watson, J. C., Goldman, R. N., & Greenberg, L. S. (2004). 学习情绪聚焦疗法. 华盛顿特区;美国心理学会.
Ellis, G. F. R., & Toronchuk, J. A. (2005). 神经发育:情感和免疫系统的影响。见 R. D. Ellis & N. Newton (编), 意识与情绪:能动性、有意识的选择和选择性感知 (第81–119页). 阿姆斯特丹:John Benjamins出版社.
Elsinger, P. J., Grattan, L. M., Damasio, H., & Damasio, A. R. (1992). 儿童期额叶损伤的发展后果。神经病学档案, 49, 764–769.
Epstein, M. (1993). 后共产主义的后现代主义。共同知识, 2(3), 110–111.
Evans, C. S. (1967). 短尾猕猴的饲养方法和社会互动。动物行为, 15, 263–266.
Everitt, B. J. (1990). 性动机:雄性大鼠食欲和交配反应机制的神经和行为分析。神经科学与生物行为评论, 14, 217–232.
Fagen, R. (1981). 动物游戏行为. 纽约,纽约州:牛津大学出版社.
Faimberg, H. (2007). 呼吁延后性(Nachträglichkeit)概念的扩展。精神分析季刊, 76, 1221–1240.
Fanselow, M. S., & Poulos, A. M. (2005). 哺乳动物联想学习的神经科学。心理学年度评论, 56, 207–234.
Faure, A., Richard, J. M., & Berridge, K. C. (2010). 来自伏隔核的欲望与恐惧:皮层谷氨酸和皮层下GABA在大鼠中差异性地产生动机和享乐影响。公共科学图书馆·综合, 5(6), e11223.
Faure, A., Reynolds, S. M., Richard, J. M., & Berridge K. C. (2008). 欲望和恐惧中的中脑边缘多巴胺:通过伏隔核中局部谷氨酸干扰使动机得以产生。神经科学杂志, 28, 7184–7192.
Fearon, R. P., Bakermans-Kranenburg, M. J., van Ijzendoorn, M. H., Lapsley, A. M., & Roisman, G. I. (2010). 不安全依恋和无序性在儿童外化行为发展中的意义:一项元分析研究。儿童发展, 81, 435–456.
Feifel, D., Macdonald, K., Nguyen, A., Cobb, P., Warlan, H., Galangue, B., Minassian, A., Becker, O., Cooper, J., Perry, W., Lefebvre, M., Gonzales, J., & Hadley, A. (2010). 辅助性鼻内催产素减轻精神分裂症患者的症状。生物精神病学, 68, 678–680.
Feldman, R., Gordon, I., Schneiderman, I., Weisman, O., & Zagoory-Sharon, O. (2010). 母性和父性照料的自然变异与亲子接触后催产素的系统性变化相关。心理神经内分泌学, 35, 1133–1141.
Ferris, C. F., Kulkarni, P., Sullivan, J. M. Jr., Harder, J. A., Messenger, T. L., & Febo, M. (2005). 幼崽吮吸比可卡因更具奖励性:来自功能性磁共振成像和三维计算分析的证据。神经科学杂志, 25, 149–156.
Fibiger, H. C., & Phillips, A. G. (1986). 奖励、动机、认知:中脑端脑多巴胺系统的心理生物学。见生理学手册, 第4卷, 神经系统,大脑的内在调节系统 (第647–675页). 贝塞斯达,马里兰州:美国生理学会.
Flagel, S. B., Clark, J. J., Robinson, T. E., Mayo, L., Czuj, A., Willuhn, I., Akers, C. A., Clinton, S. M., Phillips, P. E., & Akil, H. (2011a). 多巴胺在刺激-奖励学习中的选择性作用。自然, 469, 53–57.
Flagel, S. B., Cameron, C. M., Pickup, K. N., Watson, S. J., Akil, H., & Robinson, T. E. (2011b). 食物预测线索必须被赋予激励显著性才能诱导皮层-纹状体-丘脑脑区中c-fos mRNA的表达。神经科学, 196, 80–96.
Fleming, A. S., & Rosenblatt, J. S. (1974). 大鼠母性行为的嗅觉调节:I. 嗅球切除对有经验和无经验的哺乳期和发情周期雌性的影响。比较与生理心理学杂志, 86, 221–232.
Fleming, A. S., O’Day, D. H., & Kraemer, G. W. (1999). 母婴互动的神经生物学:跨发展阶段和代际的经验与中枢神经系统可塑性。Neuroscience and Biobehavioral Reviews, 23, 673–685.
Flynn, J. P. (1976). 威胁和攻击的神经基础。In R. G. Grenell & S. G. Abau (Eds.), 精神病学的生物学基础 (pp. 275–295). New York, NY: Raven Press.
Foa, E. B., & Kozak, M. J. (1998). 生物信息理论的临床应用:理解焦虑及其治疗。Behavior Therapy, 29, 675–690.
Fogelin, R. J. (2003). 走在理性的钢丝上:理性动物的不稳定生活. New York, NY: Oxford University Press.
Fosha, D. (2000). 情感的转化力量:加速变化的模型. New York, NY: Basic Books.
Fosha, D., Siegel, D., & Solomon, M. (Eds.). (2009a). 具身心智:临床实践中身体、大脑和心智的整合. New York, NY: W. W. Norton.
Fosha, D., Siegel, D. J., & Solomon, M. F. (Eds.). (2009b). 情感的治愈力量:情感神经科学、发展与临床实践. New York, NY: W. W. Norton.
Fox, M. D., and Raichle, M. E. (2007). 通过功能性磁共振成像观察到的大脑活动自发波动。Nature Reviews Neuroscience, 8, 700–711.
Fransson, P. (2006). 大脑功能的默认模式有多默认?来自内在BOLD信号波动的进一步证据。Neuropsychologia, 44, 2836–2845.
Freed, P. J., & Mann, J. J. (2007). 悲伤与失落:走向神经生物心理社会模型。American Journal of Psychiatry, 164, 28–34.
Freed, P. J., Yanagihara, T. K., Hirsch, J., & Mann, J. J. (2009). 悲伤调节的神经机制。Biological Psychiatry, 66, 33–40.
Freud, S. (1895/1968). 科学心理学方案。In 精神分析的起源. SE I. London: Hogarth Press. (原作发表于1895年)
Freud, S. (1905a/1968). 笑话及其与无意识的关系. 标准版 8. London: Hogarth Press. (原作发表于1905年)
Freud, S. (1905b/1968). 性学三论. 标准版 7. London: Hogarth Press, 1968. (原作发表于1905年)
Freud, S. (1937/1968). 分析中的建构. 标准版 23. London: Hogarth Press. (原作发表于1937年)
Galenson, E., & Roiphe, H. (1974). 生命第二年中生殖器意识的出现。In R. C. Friedman, R. M. Richard & R. L. van Wiele (Eds.), 行为中的性别差异 (pp. 223–231). New York, NY: Wiley.
Gallagher, S. (2000). 自我的哲学概念:对认知科学的启示。Trends in Cognitive Sciences, 4, 14–21.
Gallagher, H. L., & Frith, C. D. (2003). “心智理论”的功能成像。Trends in Cognitive Sciences, 7, 77–83.
Gallagher, S., & Shear, J. (Eds.). (1999). 自我模型. Exeter, UK: Imprint Academic.
Ganasen, K. A., Ipser, J. C., & Stein, D. J. (2010). 用药物疗法增强认知行为疗法。Psychiatric Clinics of North America, 33, 687–699.
Gardner, H. (1985). 心智的新科学:认知革命史. New York, NY: Basic Books.
Gavrilets, S., & Rice, W. R. (2006). 同性恋的遗传模型:生成可检验的预测。Proceedings of the Royal Society B, 273, 3031–3038.
Gedo, J. (1997). 怨愤与怀旧:精神分析中的生活和工作. New York, NY: Jason Aronson.
Georgiadis, J. R., Farrell, M. J., Boessen, R., Denton, D. A., Gavrilescu, M., Kortekaas, R., Renken, R. J., Hoogduin, J. M., & Egan, G. F. (2010). 具有生态效度的男性性活动期间的动态皮层下血流:一项灌注fMRI研究。Neuroimage, 50, 208–216.
Georgiadis, J. R., Kortekaas, R., Kuipers, R., Nieuwenburg, A., Pruim, J., Reinders, A. A., & Holstege, G. (2006). 健康女性阴蒂诱导性高潮相关的区域脑血流变化。European Journal of Neuroscience, 24, 3305–3316.
Georgiadis, J. R., Reinders, A. A., Paans, A. M., Renken, R., & Kortekaas, R. (2009). 男性与女性在性脑功能上的差异:触觉生殖器刺激期间的显著差异,但在性高潮期间没有差异。Human Brain Mapping, 30, 3089–3101.
Giegling, I. Rujescu, D., Mandelli, L., Schneider, B., Hartmann, A. M., Schnabel, A., Maurer, K., De Ronchi, D., Möller, H. J., & Serretti, A. (2007). 速激肽受体1变体与自杀行为中的攻击性相关。American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, 144B(6), 757–761.
Gleason, E. D., Fuxjager, M. J., Oyegbile, T. O., & Marler, C. A. (2009). 睾酮释放和社会背景:何时发生及其原因。Frontiers in Neuroendocrinology, 30, 460–469.
Goel, V., & Dolan, R. J. (2003). 热推理和冷推理期间外侧和腹内侧前额叶皮层内的相互神经反应。Neuroimage, 20, 2314–2321.
Goldman-Rakic, P. S. (1998). 皮层多巴胺系统:在记忆和认知中的作用。Advances in Pharmacology, 42, 707–711.
Goldstein, J. M. (2006). 精神分裂症中的性别、激素和情感唤起回路功能障碍。Hormones & Behavior, 50, 612–622.
Goleman, D. (2006). 社交智能:人际关系的新科学. New York, NY: Arrow.
Goodall, J. (1986). 冈贝的黑猩猩. Cambridge, MA: Harvard University Press.
Goodson, J. L., & Bass, A. H. (2001). 脊椎动物中加压素/抗利尿激素系统的社会行为功能和相关解剖特征。Brain Research Reviews, 35, 246–265.
Gooren, L. (2006). 人类心理性分化的生物学基础。Hormones and Behavior, 50, 589–601.
Gordon, N. S., Burke, S., Akil, H., Watson, S., & Panksepp, J. (2003). 社交诱导的大脑”施肥”:玩耍促进幼年大鼠杏仁核和背外侧额叶皮层脑源性神经营养因子的转录。神经科学通讯, 341, 17–20.
Gordon, N. S., Kollack-Walker, S., Akil, H., & Panksepp, J. (2002). 幼年大鼠在打闹玩耍过程中c-fos基因激活的表达。脑研究通报, 57, 651–659.
Gorski, R. A. (1988). 大脑的性别分化:神经科学的机制和影响。见 S. S. Easter Jr., K. F. Barald, 和 B. M. Carlson (主编), 从信息到心智:发育神经生物学方向 (pp. 256–271). Sunderland, MA: Sinauer.
Gottesman, I. I., & Gould, T. D. (2003). 精神病学中的内表型概念:词源和战略意图。美国精神病学杂志, 160, 636–645.
Grace, A. A. (1991). 相位性与强直性多巴胺释放以及多巴胺系统反应性的调节:精神分裂症病因学假说。神经科学, 41, 1–24.
Graeff, F. G. (2004). 血清素、导水管周围灰质和恐慌。神经科学与生物行为评论, 28, 239–259.
Grandin, T. (2005). 动物的翻译. New York, NY: Scribner.
Grandin, T. & Johnson, C. (2009). 动物让我们成为人类. Orlando, FL: Houghton Mifflin Harcourt Publishing Company.
Gray, J. (1990). 同时介导情绪和认知的大脑系统。认知与情绪, 4, 269–288.
Gray, J. (1992). 男人来自火星,女人来自金星. New York, NY: Harper Collins.
Gray, L., Miller, L. W., Philipp, B. L., & Blass, E. M. (2002). 母乳喂养对健康新生儿具有镇痛作用。儿科学, 109, 590–593.
Green, J. J., & Hollander, E. (2010). 自闭症和催产素:治疗转化方法的新进展。神经治疗学, 7, 250–257.
Greenberg, L. (2002). 情绪聚焦疗法:指导客户处理情感. Washington, DC: American Psychological Association.
Greenberg, L., & Watson, J. (2005). 抑郁症的情绪聚焦疗法. Washington, DC: American Psychological Association.
Greenspan, R. J., & Baars, B. J. (2005). 意识的黯淡:Jacques Loeb、Ivan P. Pavlov和1900年后还原论生物学的兴起。意识与认知, 14, 219–230.
Gregg, T., & Siegel, A. (2003). 中脑导水管周围灰质NK1受体对猫的防御性愤怒和捕食性攻击的差异影响。脑研究, 994, 55–66.
Griffin, D. R. (1984). 动物的思维. Cambridge, MA: Harvard University Press.
Griffin, D. R. (2001). 动物的心智:超越认知到意识. Chicago: University of Chicago Press.
Grimm, S., Beck, J., Schuepbach, D., Hell, D., Boesiger P., Bermpohl, F., Niehaus, L., Boeker, H., & Northoff G. (2008). 重度抑郁症中左右背外侧前额叶皮层失衡与负面情绪判断相关:严重重度抑郁障碍的fMRI研究。生物精神病学, 63, 369–376.
Grimm, S., Boesiger P., Beck, J., Schuepbach, D., Bermpohl, F., Walter, M, Ernst, J., Hell, D., & Northoff, G. (2009). 抑郁受试者在情绪处理过程中默认模式网络的负性BOLD反应改变。神经精神药理学, 34, 932–943.
Grippo, A. J, Trahanas, D. M., Zimmerman, R. R. 2nd, Porges, S. W., & Carter, C. S. (2009). 催产素防止长期社交隔离的负面行为和自主神经后果。精神神经内分泌学, 34, 1542–1553.
Groos, K. (1898). 动物的玩耍 (E.L. Baldwin, 译). New York, NY: Appleton.
Groos, K. (1901). 人类的玩耍 (E.L. Baldwin, 译). New York, NY: Appleton.
Grosenick, L., Greer, S., & Knutson, B. (2008). 可解释的fMRI分类器改善购买预测。IEEE神经系统与康复工程会刊, 16, 539–548.
Gross, J. J. (2008). 情绪调节。见 M. Lewis, J. M. Haviland-Jones, 和 L.F. Barrett (主编), 情绪手册 (第3版, pp. 497–512). New York, NY: Guilford.
Gross, J. J. (主编). (2009). 情绪调节手册. New York, NY: Guilford.
Gross-Isseroff, R., Biegon, A., voet, H., & Wezman, A. (1998). 自杀的大脑:死后受体/转运体结合研究综述。神经科学与生物行为评论, 22, 653–661.
Guastella, A. J., Howard, A. L., Dadds, M. R., Mitchell, P., & Carson, D. S. (2009). 鼻内催产素作为社交焦虑障碍暴露疗法辅助治疗的随机对照试验。精神神经内分泌学, 34, 917–923.
Guerra, D. J., Colonnello, V., & Panksepp, J. (2010). 愤怒和愤怒的神经生物学及其精神病学意义,重点关注抑郁症。见 F. Pahlavan (主编), 愤怒的多个面向:发怒还是恢复正义? (pp. 81–103). New York, NY: Nova Science.
Gur, R.C., Mozley, L.H., Mozley, P. D., Resnick, S. M., Karp, J. S., Alavi, A., Arnold, S. E., & Gur, R. E. (1995). 静息状态下区域脑葡萄糖代谢的性别差异。科学, 267, 528–531.
Gurian, M. (2004). 他在想什么?男人的大脑真正如何运作. New York, NY: St. Martin’s Griffin.
Haber, S. N., & Knutson, B. (2010). 奖赏回路:连接灵长类解剖学和人类成像。神经精神药理学评论, 35, 4–26.
Halasz, J., Toth, M., Mikics, E., Hrabovszky, E., Barsy, B., Barsvari, B., & Haller, J. (2008). 神经激肽1受体阻断对领地攻击和暴力攻击模型的影响。生物精神病学, 63, 271–278.
Hamburg, M. D. (1971). 下丘脑单元活动和进食行为。美国生理学杂志, 220, 980–985.
Hansen, S., & Kohler, C. (1984). 脚踏核周围核在大鼠性行为和射乳神经内分泌控制中的重要性。Neuroendocrinology, 39, 563–572.
Hao, S., Liu, S., Zheng, X., Zheng, W., Ouyang, H., Mata, M., & Fink, D. J. (2010). TNFα在纳洛酮催发的大鼠吗啡戒断期间导水管周围灰质中的作用。Neuropsychopharmacology, 36, 664–676.
Hardt, O., Einarsson. E. O., & Nader, K. (2010). 跨越困境之桥:再巩固作为认知与神经科学记忆研究传统之间的联系。Annual Review of Psychology, 61, 141–167.
Harlow, H. F. (1958). 爱的本质。American Psychology, 13, 673–685.
Harmon, K. M., Cromwell, H. C., Burgdorf, J., Moskal, J. R., Brudzynski, S. M., Kroes, R. A., & Panksepp, J. (2008). 选择性培育低水平50 kHz超声发声的大鼠在早期社交动机上表现出改变。Developmental Psychobiology, 50, 322–231.
Harmon-Jones, E., & Winkielman, P. (2007). 社会神经科学:整合社会行为的生物学和心理学解释。纽约:Guilford出版社。
Harro, J. (2010). 神经生物学的个体差异作为情感障碍的脆弱性因素:对精神药理学的意义。Pharmacological Therapeutics, 125, 402–422.
Harro, J., & Oreland, L. (2001). 抑郁症作为单胺能神经元的扩散性适应障碍:蓝斑核主要牵连的案例。Brain Research Reviews, 38, 79–128.
Harro, J., Kanarik, M., Matrov, D., & Panksepp, J. (2011). 用细胞色素氧化酶组织化学方法绘制抑郁症相关脑区模式:动物情感系统与人类障碍的相关性,重点关注对不良事件的韧性。Neuroscience & Biobehavioral Reviews, 35, 1876–1889.
Hart, S. L., & Legerstee, M. (主编). (2010). 嫉妒手册:理论、原则和多学科方法。纽约,NY:Wiley-Blackwell出版社。
Hartman, S. (2011). 再次向内探索(精神的生物学)。www.eonwriter.com。
Haubensak, W., Kunwar, P. S., Cai, H., Ciocchi, S., Wall, N. R., Ponnusamy, R., Biag, J., Dong, H. W., Deisseroth, K., Callaway, E. M., Fanselow, M. S., Lüthi, A., & Anderson. D. J. (2010). 调控条件性恐惧的杏仁核微环路的遗传解析。Nature, 468, 270–276.
Heath, R. (1996). 探索身心关系。巴吞鲁日,LA:Moran印刷。
Heath, R. G., Llewellyn, R. C., & Rouchell, A. (1980). 用于难治性行为障碍和癫痫的小脑起搏器:后续报告。Biological Psychiatry, 15, 254–256.
Hebb, D. O. (1949). 行为的组织。纽约,NY:John Wiley出版社。
Heidbreder, C., Gewiss, M., DeMot, B., Mertens, I., & De Witte, P. (1992). 在大鼠脑内注射胆囊收缩素和神经降压素后,伏隔核中谷氨酸和多巴胺的平衡调节自我刺激行为。Peptides, 13, 441–449.
Heim, C., & Nemeroff, C. B. (1999). 早期不良经历对焦虑和情感障碍病理生理学相关脑系统的影响。Biological Psychiatry, 4, 1509–1522.
Heim, C., Plotsky, P. M., & Nemeroff, C. B. (2004). 研究早期不良经历对抑郁症神经生物学发现的贡献的重要性。Neuropsychopharmacology, 29, 641–648.
Heim, C., Shugart, M., Craighead, W. E., & Nemeroff, C. B. (2010). 儿童虐待和忽视的神经生物学和精神病学后果。Developmental Psychobiology, 52, 671–690.
Hein, G., & Singer, T. (2008). 我感受到你的感受,但并非总是如此:共情脑及其调节。Current Opinions in Neurobiology, 18, 153–158.
Heinrichs, M., & Domes, G. (2008). 神经肽和社会行为:催产素和加压素对人类的影响。Progress in Brain Research, 170, 337–350.
Heinrichs, M., von Dawans, B., & Domes, G. (2009). 催产素、加压素和人类社会行为。Frontiers in Neuroendocrinology, 30, 548–557.
Heresco-Levy, U., Ermilov, M., Shimoni, J., Shapiro, B., Silipo, G. & Javitt, D. C. (2002). D-环丝氨酸添加到传统抗精神病药物、奥氮平和利培酮治疗精神分裂症的安慰剂对照试验。American Journal of Psychiatry, 159, 480–482.
Herman, B. H., & Panksepp, J. (1978). 吗啡和纳洛酮对分离痛苦和接近依恋的影响:阿片类物质介导社会情感的证据。Pharmacol Biochem Behav, 9(2), 213–220.
Herman, B. H., and Panksepp, J. (1981). 内啡肽对痛苦发声的上行抑制。Science, 211, 1060–1062.
Hermans, E. J., Ramsey, N. F., & van Honk, J. (2008) 外源性睾酮增强人类社会攻击神经回路对社会威胁的反应性。Biological Psychiatry, 63, 263–270.
Hess, W. R. (1954). 间脑:自主神经和锥体外系功能。纽约:Grune和Stratton出版社。
Hess, W. R. (1957). 间脑的功能组织。纽约,NY:Grune和Statton出版社。
Hess, W. R. (1964). 心智的生物学。芝加哥:芝加哥大学出版社。
Hitchcock, E., & Cairns, V. (1973). 杏仁核切除术。Postgraduate Medicine, 49, 894–904.
Hoffman, R. (主编). (2011). 崇高。纽约,NY:牛津大学出版社。
Hofmann, S. G., Pollack, M. H., & Otto, M. W. (2006). 用D-环丝氨酸增强焦虑障碍心理治疗的治疗。CNS Drug Reviews, 12, 208–217.
Hole, G. J., & Einon, D. F. (1984). 啮齿动物的游戏。在P. K. Smith (主编), 动物和人类的游戏 (第95–117页)。纽约,NY:Basil Blackwell出版社。
Holsboer, F., & Ising M. (2008). 抑郁和焦虑中的中枢CRH系统—来自CRH1受体拮抗剂临床研究的证据。欧洲药理学杂志,583,350–357。
Holstege, G., Georgiadis, J. R., Paans, A. M., Meiners, L. C., van der Graaf, F. H., & Reinders, A.A. (2003). 人类男性射精期间的大脑激活。神经科学杂志,23,9185–9193。
Holstege, G. R., & Saper, C. B. (2005). 特刊:灵魂的解剖学。比较神经学杂志,493,1–176。
Holt, R. R. (1989). 重新评价弗洛伊德。纽约:Guilford出版社。
Holtzheimer, P. E. III, & Nemeroff, C. B. (2006). 抑郁症治疗的进展。美国实验神经治疗学会杂志,3,42–56。
Hosobuchi, Y., Rossier, J., Bloom, F. E., & Guillemin, R. (1979). 刺激人类导水管周围灰质以缓解疼痛会增加脑室液中的免疫反应性β-内啡肽。科学,203,279–281。
Hoyenga, K. B., & Hoyenga, K. T. (1993). 性别相关差异:起源和结果。波士顿:Allyn & Bacon出版社。
Hrdy, S. (2009). 母亲与他人:相互理解的进化起源。剑桥:哈佛大学出版社。
Hubel, D., & Wiesel, T. (1979). 视觉的大脑机制。科学美国人241,150–162。
Huber, R., Panksepp, J. B., Nathaniel, T., Alcaro, A. and Panksepp, J. (2011). 螯虾中的药物敏感奖赏:用于研究寻求、奖赏、成瘾和戒断的无脊椎动物模型系统,神经科学与生物行为评论,35,1847–1853。
Huber, D., Veinante, P., & Stoop, R. (2005). 加压素(vasopressin)和催产素(oxytocin)兴奋中央杏仁核中不同的神经元群。科学,308,245–247。
Hughes, D. A. (2006). 建立依恋的纽带:唤醒深度困扰儿童的爱(第2版)。拉纳姆:Jason Aronson出版社。
Hughes, D. A. (2007). 依恋聚焦的家庭治疗。纽约:W. W. Norton出版社。
Hume, D. (1739). 人性论:将实验推理方法引入道德学科的尝试。纽约:企鹅经典。
Hume, D. (1748/1910). 人类理解研究。哈佛经典第37卷。纽约:P. F. Collier & Son出版社。
Huston, J. P., & Borbély, A. A. (1973). 通过使用奖赏性下丘脑刺激对前脑切除大鼠进行操作性条件反射。脑研究,50,467–472。
Huston, J. P., & Borbély, A. A. (1974). 丘脑大鼠:一般行为、使用奖赏性下丘脑刺激的操作性学习以及苯丙胺的影响。生理学与行为,12,433–448。
Hyman, S. E. (2007). 神经科学能否整合到DSM-V中?自然神经科学评论,8,725–732。
Iacoboni, M. (2009a). 模仿、共情和镜像神经元。心理学年度评论,60,19.1–19.18。
Iacoboni, M. (2009b). 镜像他人:我们如何与他人建立联系的新科学。纽约:Farrar, Straus and Giroux出版社。
Ikemoto, S. (2007). 多巴胺奖赏回路:从腹侧中脑到伏隔核–嗅结节复合体的两个投射系统。脑研究评论,56,27–78。
Ikemoto, S. (2010). 中脑边缘多巴胺系统之外的脑奖赏回路:一种神经生物学理论。神经科学与生物行为评论,35,129–150。doi: 10.1016/j.neubiorev.2010.02.001
Ikemoto, S., & Panksepp, J. (1992). 早期社会隔离对幼年大鼠社交游戏动机的影响。发育心理生物学,25,261–274。
Ikemoto, S., and Panksepp, J. (1994). 大鼠自我刺激与嗅探之间的关系:是否有共同的大脑系统介导这些行为?行为脑研究,61,143–162。
Ikemoto, S., and Panksepp, J. (1999). 伏隔核多巴胺在动机行为中的作用:特别参考奖赏寻求的统一解释。脑研究评论,31,6–41。
Insel, T. R. (1990). 催产素与母性行为。见N. A. Krasnegor & R. S. Bridges(编),哺乳动物的养育(第260–280页)。纽约:牛津大学出版社。
Insel, T. R. (2003). 社会依恋是一种成瘾性障碍吗?生理学与行为,79,351–357。
Insel, T. R. (2010). 社会神经科学中转化的挑战:催产素、加压素和亲和行为的综述。神经元,65,768–779。
Insel, T. R., & Harbaugh, C. R. (1989). 下丘脑室旁核损伤破坏母性行为的启动。生理学与行为,45,1033–1041。
Insel, T. R., & Shapiro, L. E. (1992). 催产素受体与母性行为。见C. A. Pedersen, J. D. Caldwell, & G. F. Jirikowski(编),催产素在母性、性和社会行为中的作用(第122–141页)。纽约:纽约科学院。
Insel, T. R., & Young, L. J. (2001). 依恋的神经生物学。自然神经科学评论,2,129–136。
Irle, E., Lange, C., Sachsse, U., & Weniger, G. (2009). 进一步证据表明创伤后应激障碍(post-traumatic stress disorder)而非分离性障碍(dissociative disorders)与创伤暴露个体的杏仁核和海马体积缩小有关。斯堪的纳维亚精神病学学报,119(4),330–331。
Ishak, W. W., Kahloon, M., & Fakhry, H. (2010). 催产素在增强幸福感中的作用:文献综述。情感障碍杂志,130,1–9。
Izard, C. E. (2007). 基本情绪、自然种类、情绪图式和新范式。心理科学展望,2,260–268。
Izdebski, K.(编)。(2008). 人声中的情绪,第1–3卷。圣迭戈:Plural出版社。
Jacob, F. (1977). 进化与修补。科学,196,1161–1166。
James, W. (1968). 什么是情绪?见M. Arnold(编),情绪的本质。巴尔的摩:企鹅出版社。(原著出版于1884年。)
James, W. (1892). 心理学:简明教程。纽约:Harper & Row出版社。
Janov, A. (2007). 原始治愈。Franklin Lakes, NJ: New Page Books.
Jeannerod, M. (2003). 人类自我识别的机制。行为脑研究,142,1–15。
Jirikowski, G. F., Caldwell, J. D., Stumpf, W. E., & Pederson, C. A. (1988). 雌二醇影响催产素免疫反应性脑系统。神经科学,25,237–248。
Johnson, S., Hollis, C., Kochhar, P., Hennessy, E., Wolke, D., & Marlow, N. (2010). 极早产儿童的自闭症谱系障碍。儿科学杂志,156,525–531.e2。
Jonason, K. R., Enlloe, L. J., Contrucci, J., & Meyer, P. M. (1973). 同时和连续隔膜及杏仁核损伤对大鼠社会行为的影响。比较与生理心理学杂志,83,54–61。
Jürgens, U. (2002). 发声控制的神经通路。神经科学与生物行为评论,26,235–258。
Kalin, N. H., Shelton, S. E., & Barksdale, C. M. (1988). 阿片类物质对非人灵长类动物分离诱导痛苦的调节。脑研究,440,285–292。
Kalin, N. H., Shelton, S. E., Davidson, R. J., & Kelley, A. E. (2001). 灵长类动物杏仁核介导急性恐惧,但不介导焦虑气质的行为和生理成分。神经科学杂志,21,2067–2074。
Kalogeras, K. T., Neiman, L. K., Friedman, T. C., Doppman, J. L., Cutler, G. B. Jr., Chrousos, G. P., Wilder, R. L., Gold, P. W. Q., & Yanovski, J. A. (1996). 健康人类受试者的岩下窦采样显示,单侧促肾上腺皮质激素释放激素诱导的精氨酸加压素释放与同侧促肾上腺皮质激素分泌相关。临床研究杂志,97,2045–2050。
Kanarik, M., Alttoa, A., Matrov, D., Koiv, K., Sharp, T., Panksepp, J., & Harro, J. (2010). 大脑对慢性社会失败压力的反应:区域氧化代谢的影响作为享乐特质的函数,以及易感和抗性大鼠的基因表达。欧洲神经精神药理学杂志,21,92–107。
Kandel, E. (2007). 追寻记忆:心智新科学的诞生。纽约,NY: W. W. Norton。
Kaplan-Solms, K., & Solms, M. (2000). 神经精神分析的临床研究:深度神经心理学导论。伦敦:Karnac。
Kassel, J. D. (2010). 物质滥用与情绪。华盛顿特区:美国心理学会。
Kehoe, P., & Blass, E. M. (1986). 10日龄大鼠分离痛苦的阿片类物质介导:母体刺激对压力的逆转。发展心理生物学,19,385–398。
Kelley, A. E. (1999). 伏隔核亚区域在学习和动机方面的神经整合活动。心理生物学,27,198–213。
Kelley, A. E. (2004). 记忆与成瘾:共享的神经回路和分子机制。神经元,44,161–179。
Keltner, D. (2009). 生而向善:有意义生活的科学。纽约,NY: W. W. Norton。
Kendrick, K. M. (2000). 催产素、母性与依恋。实验生理学,85,111S–124S。
Kendrick, K. M., Levy, F., and Keverne, E. B. (1992). 绵羊出生诱导的嗅觉信号感觉处理变化。科学,256,833–836。
Kennedy, S. E., Koeppe, R. A., Young, E. A., & Zubieta, J. K. (2006). 女性重度抑郁症患者内源性阿片类物质情绪调节回路的失调。普通精神病学档案,63,1199–1208。
Kerchner, G. A., & Nicoll, R. A. (2008). 沉默突触与长时程增强突触后机制的出现。自然神经科学评论,9,813–825。
Keverne, E. B., & Kendrick, K. M. (1994). 绵羊的母性行为及其神经内分泌调节。儿科学报增刊,379,47–56。
Keverne, E. B., Levy, F., Poindron, P., and Lindsay, D. R. (1983). “阴道刺激:绵羊母体依恋的重要决定因素。” 科学,219,81。
Keverne, E. B., Martensz, N., & Tuite, B. (1989). 猴子脑脊液中β-内啡肽浓度受理毛关系影响。心理神经内分泌学,14,155–161。
Keverne, E. B., Nevison, C. M., & Martel, F. L. (1997). 早期学习与社会纽带。纽约科学院年鉴,807,329–339。
Kimbrell, T. A., George, M. S., Parekh, P. I., Ketter, T. A., Podell, D. M., Danielson, A. L., Repella, J. D., Benson, B. E., Willis, M. W., Herscovitch, P., & Post, R. M. (1999). 健康成人短暂自我诱导焦虑和愤怒期间的区域脑活动。生物精神病学,46,454–465。
Kimchi, T., Xu, J., & Dulac, C. (2007). 雌性小鼠大脑中支持雄性性行为的功能回路。自然,448,1009–1014。
King, B. E., Packard, M. G., & Alexander, G. M. (1999). 雄性大鼠内侧视前区注射睾酮的情感特性。神经科学通讯,269,149–152。
King, H. E. (1961). 边缘系统兴奋的心理效应。In D. E. Sheer (Ed.),大脑电刺激 (pp. 477–486)。奥斯汀:德克萨斯大学出版社。
Kinsey, A. C., Pomeroy, W. B., & Martin, C. E. (1948). 人类男性的性行为。费城:W. B. Saunders。
Kinsley, C. H., & Lambert, K. G. (2006). 母性大脑。科学美国人,294,72–79。
Kinsley, C. H., & Lambert, K. G. (2008). 生殖诱导的神经可塑性:与后代生产和照料相关的自然行为和神经改变。神经内分泌学杂志,20,515–525。
Kinsley, C. H., Bardi, M., Karelina, K., Rima, B., Christon, L., Friedenberg, J., & Griffin, G. (2008). 母性在大鼠中诱导并维持整个生命周期的行为和神经可塑性。性行为档案,37,43–56。
Kisilevsky, B. S., & Davies, G. A. (2007). 生长受限胎儿的听觉处理缺陷影响后期语言发展。医学假说,68,620–628。
Klein, D. F., & Rabkin, J. (Eds.). (1981). 焦虑:新研究与概念变化. 纽约:Raven出版社。
Klüver, H., & Bucy, P. C. (1939). 猴子颞叶功能的初步分析。神经学与精神病学档案, 42, 979–1000。
Knack, J. M., Gomez, H. L., & Jensen-Campbell, L. A. (2011). 欺凌及其长期健康影响。见 G. MacDonald & L. A. Jensen-Campbell (编), 社交痛苦:丧失与排斥的神经心理学和健康影响 (pp. 215–236)。华盛顿特区:美国心理学会。
Knoll, A.T., & Carlezon, W. A. Jr. (2010). 强啡肽(Dynorphin)、压力与抑郁。脑研究, 1314, 56–73。
Knutson, B., Burgdorf, J., & Panksepp, J. (1998). 对玩耍的期待会引发幼年大鼠的发声。比较心理学杂志, 112, 65–73。
Knutson, B., Burgdorf, J., & Panksepp, J. (1999). 高频超声发声作为大鼠条件性药理奖赏的指标。生理学与行为, 66, 65–73。
Knutson, B., Burgdorf, J., & Panksepp, J. (2002). 超声发声作为大鼠情感状态的指标。心理学通报, 128(6), 961–977。
Knutson, B., & Cooper, J. C. (2005). 奖赏预测的功能性磁共振成像。神经生物学现代观点, 18, 411–417。
Knutson, B., & Greer, S. M. (2008). 预期性情感:神经相关性及其对选择的影响。伦敦皇家学会哲学汇刊B辑生物科学, 363, 3771–3786。
Knutson, B., Wolkowitz, O. M., Cole, S. W., Chan, T., Moore, E. A., Johnson, R. C., Terpstra, J., Turner, R. A., & Reus, V. I. (1998). 通过血清素干预选择性改变人格和社交行为。美国精神病学杂志, 155, 373–379。
Koenigsberg, H.W. (2010) 情感不稳定性:神经科学与心理学视角的整合。人格障碍杂志, 24, 60–82。
Kolb, B., & Tees, C. (编). (1990). 大鼠大脑皮层. 剑桥,马萨诸塞州:MIT出版社。
Kolmen, B. K., Feldman, H. M., Handen, B. L., & Janosky, J. E. (1997). 纳曲酮(Naltrexone)在幼年自闭症儿童中的应用:重复研究与学习测量。美国儿童与青少年精神病学会杂志, 36, 1570–1578。
Konner, M. (2010). 童年的进化. 剑桥,马萨诸塞州:哈佛大学出版社贝尔克纳普出版分社。
Koob, G. F., & Le Moal, M. (2001). 药物成瘾、奖赏失调与异态平衡(allostasis)。神经精神药理学, 24, 97–129。
Kovács, G. L., & Van Ree, M. V. (1985). 具有行为活性的催产素片段同时减弱大鼠的海洛因自我给药和耐受性。生命科学, 37, 1895–1900。
Kovács, G. L, Sarnyai, Z., & Szabó, G. (1998). 催产素与成瘾:一项综述。心理神经内分泌学, 23, 945–962。
Koziol, L. F., & Budding, D. E. (2009). 皮层下结构与认知:对神经心理评估的启示. 纽约,纽约州:施普林格出版社。
Kramer, P. D. (2005). 倾听百忧解. 纽约,纽约州:维京企鹅出版社。
Krueger, J. M., Rector, D. M., Roy, S., Van Dongen, H. P. A., Belenky, G., & Panksepp, J. (2008). 睡眠作为神经元集群的基本属性。自然神经科学评论, 9, 910–919。
Lahey, B. B., Van Hulle, C. A., Keenan, K., Rathouz, P. J., D’Onofrio, B. M., Rodgers, J. L., & Waldman, I. D. (2008). 生命第一年的气质与养育方式可预测未来儿童行为问题。异常儿童心理学杂志, 15, 548–558。
Lambert, M. J., & Barley, D. E. (2001). 治疗关系与心理治疗结果的研究综述。心理治疗:理论/研究/实践/培训, 38, 357–361。
Lambie, J. A., & Marcel, A. J. (2002). 意识与情绪体验的多样性:一个理论框架。心理学评论, 109, 219–259。
Land, B. B., Bruchas, M. R., Melief, E., Xu, M., Lemos, J., & Chavkin, C. (2008). 压力的烦躁成分由强啡肽-κ阿片系统的激活编码。神经科学杂志, 28, 407–414。
Lange, C. (1885). Om Sindsbevaegelser et Psyco.Studie. 哥本哈根:Kronar出版社。
Lanius, R. A., Williamson, P. C., Bluhm, R. L., Densmore, M., Boksman, K., Neufeld, R. W., Gati, J. S., & Menon, R. S. (2005). 创伤后应激障碍中分离反应的功能连接:功能性磁共振成像研究。生物精神病学, 57, 873–884。
Larsen, R. J., & Augustine, A. A. (2008). 与趋近和回避相关的基本人格倾向:外向性/神经质、行为激活系统(BAS)/行为抑制系统(BIS)与积极/消极情感。见 A. Elliot (编), 趋近与回避动机手册 (pp. 67–87)。马赫瓦,新泽西州:劳伦斯·厄尔鲍姆出版社。
Laureys, S., Pellas, F., Van Eeckhout, P., Ghorbel, S., Schnakers, C., Perrin, F., Berré, J., Faymonville, M. E., Pantke, K. H., Damas, F., Lamy, M., Moonen, G., & Goldman, S. (2005). 闭锁综合征:有意识但瘫痪且无法说话是什么感觉?脑研究进展, 150, 495–511。
Laviola, G., Adriani, W., Terranova, M. L., & Gerra, G. (1999). 人类青少年和动物模型中对精神兴奋剂易感性的心理生物学风险因素。神经科学与生物行为评论, 23, 993–1010。
LeDoux, J. (1996). 情绪脑:情绪生活的神秘基础. 纽约,纽约州:西蒙与舒斯特出版社。
LeDoux, J. E. (2000). 大脑中的情绪回路。神经科学年度评论, 23, 155–184。
LeDoux, J. (2002). 突触自我:我们的大脑如何成为我们. 纽约,纽约州:企鹅出版社。
LeDoux, J. (2007). 杏仁核。当代生物学, 17, R868–R874。
Léger, L., Sapin, E., Goutagny, R., Peyron, C., Salvert, D., Fort, P., & Luppi, P. H. (2010). 大鼠在清醒和异相睡眠期间表达Fos的多巴胺能神经元。化学神经解剖学杂志, 39, 262–271.
Le Gros Clark, W. E., Beattie, J., Riddoch, G., & Dott, N. M. (1938). 下丘脑的形态学、功能、临床和外科方面。爱丁堡:Oliver and Boyd出版社。
Léna, I., Parrot, S., Deschaux, O., Muffat-Joly, S., Sauvinet, V., Renaud, B., Suaud-Chagny, M.-F., & Gottesmann, C. (2005). 自由活动大鼠内侧前额叶皮层和伏隔核中多巴胺、去甲肾上腺素、谷氨酸和天冬氨酸细胞外水平在睡眠-觉醒周期中的变化。神经科学研究杂志, 81, 891–899.
Leskelä, U., Rytsälä, H., Komulainen, E., Melartin, T., Sokero, P., Lestelä-Mielonen, P., & Isometsä, E. (2006). 逆境和感知的社会支持对不同抑郁症状水平受试者重度抑郁障碍结局的影响。心理医学, 36, 779–788.
Lester, B. M., Masten, A., & McEwen, B. (2007). (编). 儿童的复原力(resilience)。见于纽约科学院年鉴, 第1094卷。纽约:Wiley-Blackwell出版社。
Levin, F. M., Trevarthen, C., Colibazzi, T., Ihanus, J., Talvitie, V., Carney, J. K., & Panksepp, J. (2008). 睡眠与梦,第1部分:梦是具有情绪意义的适应性学习引擎,通过延迟行动计划帮助我们识别和处理无意识(ucs)威胁;快速眼动睡眠巩固我们在梦中学习和表达的内容的记忆。见于F. Levin (编), 情绪与小脑的心理动力学 (第3–36页)。纽约:Other Press出版社。
Levine, M. I. (1951). 关于儿童手淫的儿科观察。儿童精神分析研究, 6, 117–124.
Levy, F., Guevara-Guzman, R., Hinton, M. R., Kendrick, K. M., and Kaverne, E. B. (1993). 分娩和母性经验对绵羊嗅球中去甲肾上腺素和乙酰胆碱释放的影响。行为神经科学, 107, 662–668.
Levy, F., Kendrick, K. M., Goode, J. A., Guevara-Guzman, R., and Kaverne, E. B. (1995). 分娩母羊嗅球中催产素和加压素的释放:随母性经验的变化以及对乙酰胆碱、γ-氨基丁酸、谷氨酸和去甲肾上腺素的影响。脑研究, 669, 197–206.
Lewis, D. O., & Pincus, J. H. (1989). 癫痫与暴力:神经精神攻击综合征的证据。神经精神病学与临床神经科学杂志, 1, 413–418.
Lewis, H. B. (1976). 男性和女性的心理战争。纽约:纽约大学出版社。
Lewis, M. D. (2005). 通过动态系统建模连接情绪理论和神经生物学。行为与脑科学, 28, 169–245.
Lewis, M., Haviland, J. M., & Barrett, L. F. (编). (2008). 情绪手册 (第3版)。纽约:Guilford出版社。
Liberzon, I., Trujillo, K. A., Akil, H., & Young, E. A. (1997). 催产素在条件性位置偏好范式中的动机特性。神经精神药理学, 17, 353–359.
Lieberman, M.D., Inagaki, T.K., Tabibnia, G., & Crockette, M.J. (2011). 在标记、重新评估和分散注意力期间对情绪刺激的主观反应。情绪, 11, 468–480.
Lieberman, P. (2001). 人类语言与我们的爬行动物脑:言语、句法和思维的皮层下基础。生物学与医学视角, 44, 32–51.
Lindsay, L. W. (1880). 低等动物的心智,在健康和疾病中。纽约:D. Appleton出版社。
Linke, R., De Lima, A. D., Schwegler, H., & Pape, H. C. (1999). 大鼠中来自上丘的轴突与已识别的丘脑-杏仁核投射神经元的直接突触连接:皮层下视觉通路到杏仁核的可能基质。比较神经学杂志, 403, 158–170.
Liotti, M., & Panksepp, J. (2004a). 对人类情绪和情感感受的成像:对生物精神病学的启示。见于J. Panksepp (编), 生物精神病学教科书 (第33–74页)。霍博肯:Wiley出版社。
Liotti, M., & Panksepp, J. (2004b). 关于人类情绪的神经本质及其对生物精神病学的启示。见于J. Panksepp (编), 生物精神病学教科书 (第33–74页)。霍博肯:Wiley出版社。
London, J. (1963). 白牙及其他故事。纽约:Dodd, Mead出版社。
Lonstein, J. S., & De Vries, G. J. (2000). 啮齿动物亲代行为的性别差异。神经科学与生物行为评论, 24, 669–686.
Lorberbaum, J. P., Newman, J. D., Horwitz, A. R., Dubno, J. R., Lydiard, R. B., Hamner, M. B., Bohning, D. E., & George, M. S. (2002). 丘脑-扣带回环路在人类母性行为中的潜在作用。生物精神病学, 51, 431–445.
Lorenz, K. (1935). 鸟类环境中的伙伴。鸟类学杂志, 83, 137–213.
Lumia, A. R., & McGinnis, M. Y. (2010). 合成代谢类固醇对青春期男性的影响。生理学与行为, 100, 199–204.
MacDonald, G., & Jensen-Campbell, L. A. (编). (2011). 社交疼痛:丧失和排斥的神经心理学和健康启示。华盛顿特区:美国心理学会。
MacDonald, K., & MacDonald, T. M. (2010). 结合的肽:催产素及其在人类中的亲社会效应的系统评价。哈佛精神病学评论, 18, 1–21.
Machado-Vieira, R., Salvadore, G., Diaz Granados, & Zarate, C. A. (2009). 氯胺酮(ketamine)和下一代快速起效抗抑郁药。药理治疗学, 123, 143–150.
MacLean, P. (1990). 进化中的三位一体脑:在古脑功能中的作用。纽约:Plenum出版社。
MacLean, P. D., & Ploog, D. W. (1962). 阴茎勃起的大脑表征。神经生理学杂志, 25, 29–55.
Malan, D. (1979). Individual psychotherapy and the science of psychodynamics. London: Butterworth-Heinemann.
Malan, D. (1999) Individual psychotherapy and the science of psychodynamics (2nd ed.). Oxford: Butterworth-Heinemann.
Malloch, S. & Trevarthen, C. (2009). Communicative musicality: Exploring the basis of human companionship. New York, NY: Oxford University Press.
Mancia, G., & Zanchetti, A. (1981). Hypothalamic control of autonomic functions. In P. J. Morgane and J. Panksepp (Eds.), Handbook of the hypothalamus: Vol. 3, part B, Behavioral studies of the hypothalamus (pp. 147–202). New York, NY: Marcel Dekker.
Manning, J. T. (2002). Digit ratio: A pointer to fertility, behavior and health. New Brunswick, NJ: Rutgers University Press.
Maren, S. (2005). Building and burying fear memories in the brain. Neuroscientist, 11, 89–99.
Maren, S., & Quirk, G. J. (2004). Neuronal signaling of fear memory. Nature Reviews Neuroscience, 5, 844–852.
Mark, V. H., Ervin, F. R., & Sweet, W. H. (1972). Deep temporal lobe stimulation in man. In E. B. Eleftheriou (Ed.), The neurobiology of the amygdala (pp. 485–507). New York, NY: Plenum.
Marks, L. S. (2004). 5α-还原酶:历史与临床重要性。Reviews in Urology, 6 (Supp. 9), S11–S21.
Martinson, F. M. (1994). The sexual life of children. Westport, CT: Bergin & Garvey.
Masserman, J. H. (1941). Is the hypothalamus a center for emotion? Psychosomatic Medicine, 3, 3–25.
Maté, G. (2008). In the realm of hungry ghosts: Close encounters with addiction. Toronto: Knopf.
Matthiesen, A. S., Ransjö-Arvidson, A. B., Nissen, E., & Uvnäs-Moberg, K. (2001). Postpartum maternal oxytocin release by newborns: Effects of infant hand massage and sucking. Birth, 28, 13–19.
Mayberg, H. S. (2009). Targeted electrode-based modulation of neural circuits for depression. Journal of Clinical Investigations, 119, 717–725.
Mayberg, H. S., Lozano, A. M., Voon, V., McNeely, H. E., Seminowicz, D., Hamani, C., Schwalb, J. M., & Kennedy, S. H. (2005). Deep brain stimulation for treatment-resistant depression. Neuron, 45, 651–660.
Mayer, D. J., Wolfle, T. L., Akil, H., Carder, B., & Liebeskind, J. C. (1971). Analgesia from electrical stimulation in the brainstem of the rat. Science, 174, 1351–1354.
McCarley, R. W. (2011). Neurobiology of REM sleep. Handbook of Clinical Neurology, 9, 151–171.
McCarthy, M. M. (1990). Oxytocin inhibits infanticide in wild female house mice (Mus domesticus). Hormones and Behavior, 24, 365–375.
McCarthy, M. M., Low, L-M., & Pfaff, D. W. (1992). Speculations concerning the physiological significance of central ocytocin in maternal behaviour. Annals of the New York Academy of Sciences, 652, 253–270.
McCleary, R. A., & Moore, R. Y. (1965). Subcortical mechanisms of behavior. New York, NY: Basic Books.
McGaugh, J. L., & Roozendaal, B. (2009). Drug enhancement of memory consolidation: Historical perspective and neurobiological implications. Psychopharmacology, 202, 3–14.
McGilchrist, I. (2009). The master and his emissary: The divided brain and the making of the western world. New Haven, CT: Yale University Press.
McGinnis, M. Y., & Vakulenko, M. (2003). Characterization of 50-kHz ultrasonic vocalizations in male and female rats. Physiology & Behavior, 80, 81–88.
McGowan, P. O., Sasaki, A., D’Alessio, A. C., Dymov, S., Labonté, B., Szyf, M., Turecki, G., & Meaney, M. J. (2009). Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nature Neuroscience, 12, 342–348.
McKeon, R. (Ed.). (1941) The basic works of Aristotle. New York, NY: Random House.
McMillan, F. (Ed.). (2005). Mental health and well-being in animals. Oxford: Blackwell.
Meaney, M. J. (2001). Maternal care, gene expression, and the transmission of individual differences in stress reactivity across generations. Annual Review of Neuroscience, 24, 1161–1192.
Meaney, M. J. (2010). Epigenetics and the biological definition of gene x environment interactions. Child Development, 81, 41–79.
Meck, W. H., Penney, T. B., & Pouthas, V. (2008). Cortico-striatal representation of time in animals and humans. Current Opinions in Neurobiology, 18, 145–152.
Melis, M. R., Argioolas, A., & Gessa, G. L. (1986). Oxytocin-induced penile erection and yawning: Site of action in the brain. Brain Research, 398, 259–265.
Mendl, M., Burman, O. H. P., & Paul, E. S. (2010). An integrative and functional framework for the study of animal emotions and mood. Proceedings of the Royal Society B, 277, 2895–2904.
Mennella, J., & Moltz, H. (1988). Infanticide in rats: Male strategy and female counter-strategy. Physiology & Behavior, 42, 19–28.
Merker, B. (2007). Consciousness without a cerebral cortex: A challenge for neuroscience and medicine. Behavioral and Brain Sciences, 30, 63–134.
Meston, C. M., & Frohlich, P. F. (2000). The neurobiology of sexual function. Archives of General Psychiatry, 57, 1012–1030.
Meyer-Lindenberg, A. (2008). Impact of prosocial neuropeptides on human brain function. Progress in Brain Research, 170, 463–470.
Miczek, K. A. (1987). The psychopharmacology of aggression. In L. L. Iversen, S. D. Iversen, & S. H. Snyder (Eds.), Handbook of psychopharmacology. Vol. 19, New directions in behavioral pharmacology (pp. 183–328). New York, NY: Plenum.
Miczek, K. A. (1991). Tolerance to the analgesic, but not discriminative stimulus effects of morphine after brief social defeat in rats. Psychopharmacology, 104(2), 181–186.
Middlebrook, D. W. (1998). “Suits me”: The double life of Billy Tipton. New York, NY: Mariner.
Miller, A. H., Maletic, V., & Raison, C. L. (2009). Inflammation and its discontents: The role of cytokines in the pathophysiology of major depression. Biological Psychiatry, 65, 732–741.
Mirsky, A. F., & Siegel, A. (1994). 暴力和攻击的神经生物学。载于 A. J. Reiss, K. A. Miczek & J. A. Roth (主编),理解和预防暴力,第2卷:生物行为影响 (第59-172页)。华盛顿特区:国家科学院出版社。
Mobbs, D., Marchant, J. L., Hassabis, D., Seymour, B., Tan, G., Gray, M., Petrovic, P., Dolan, R. J., & Frith, C. D. (2009). 从威胁到恐惧:人类防御性恐惧系统的神经组织。神经科学杂志,29,12236-12243。
Mobbs, D., Petrovic, P., Marchant, J. L., Hassabis, D., Weiskopf, N., Seymour, B., Dolan, R. J., & Frith, C. D. (2007). 当恐惧临近时:威胁迫近性引发人类前额叶-导水管周围灰质转换。科学,317,1079-1083。
Modney, B. K., & Hatton, G. I. (1990). 母性以功能上有意义的方式改变大细胞神经元相互关系。载于 N. A. Krasnegor & R. S. Bridges (主编),哺乳动物育幼 (第305-323页)。纽约:牛津大学出版社。
Möhler, H. (2011). 新型GABA药理学的兴起。神经药理学,60,1042-1049。
Money, J. (1995). 性别地图:社会建构主义、女性主义和性学历史。纽约:Continuum出版社。
Mook, D. G. (1989). 食欲和饱腹感中的口腔因素。纽约科学院年鉴,575,265-278。
Moore, B. E., & Fine, B. D. (1990). 精神分析术语和概念。纽黑文和伦敦:耶鲁大学出版社和美国精神分析协会。
Morris, P. H., Doe, C., & Godsell, E. (2008). 非灵长类物种中的次级情绪?动物主人的行为报告和主观声称。认知与情绪,22,3-20。
Moskal, J.R., Burgdorf, J., Kroes, R.A., Brudzynski, S.M., & Panksepp, J. (2011). 一种新型NMDA受体甘氨酸位点部分激动剂GLYX-13在治疗自闭症方面具有潜力。神经科学与生物行为评论,35,1982-1988。
Murray, T. J., Fowler, P. A., Abramovich, D. R., Haites, N., & Lea, R. G. (2000). 人类胎儿睾丸:妊娠中期的增殖和类固醇生成能力。临床内分泌与代谢杂志,85,4812-4817。
Myers, K. M., & Davis, M. (2007). 恐惧消退机制。分子精神病学,12,120-150。
Naber, F., van IJzendoorn, M. H., Deschamps, P., van Engeland, H., & Bakermans-Kranenburg, J. (2010). 鼻内催产素增加父亲在与孩子玩耍时观察到的反应性:一项双盲受试者内实验。心理神经内分泌学,35,1583-1586。
Nader, K., & Einarsson, E. O. (2010). 记忆再巩固:最新进展。纽约科学院年鉴,1191,27-47。
Nader, K., & Hardt, O. (2009). 记忆的单一标准:再巩固的论证。自然神经科学评论,10,224-234。
Naghavi, H. R., & Nyberg, L. (2005). 注意、记忆和意识中的共同额顶叶活动:对整合的共同需求?意识与认知,14,390-425。
Narvaez, D., Panksepp, J., Schore, A., & Gleason, T. (主编). (2012). 人性、早期经验和进化适应环境。纽约:牛津大学出版社。
Nashold, B. S., Wilson, W. P., & Slaughter, G. (1969). 刺激人类中脑引发的感觉。神经外科杂志,30,14-24。
Nathaniel, T. I., Panksepp, J., & Huber, R. (2009). 无脊椎动物系统中的药物寻求行为:小龙虾中吗啡诱导奖赏、消退和恢复的证据。行为脑研究,197,331-338。
Nathaniel, T. I., Panksepp, J., & Huber, R. (2010). 单次和重复吗啡治疗对小龙虾条件性和非条件性行为敏化的影响。行为脑研究,207,310-320。
Nelson, C. A., & Bloom, F. E. (1997). 儿童发育和神经科学。儿童发展,68,970-987。
Nelson, E., & Panksepp, J. (1996). 大鼠的催产素和婴儿-母亲结合。行为神经科学,110,583-592。
Nelson, E., & Panksepp, J. (1998). 婴儿-母亲依恋的脑基础:阿片类物质、催产素和去甲肾上腺素的贡献。神经科学与生物行为评论,22,437-452。
Nelson, R. J., Trainor, B. C., Chiavegatto, S., & Demas, G. E. (2006). 一氧化氮对攻击行为的多效性贡献。神经科学与生物行为评论,30,346-355。
Newman, J. D. (主编). (1988). 哺乳动物发声的生理控制。纽约:Plenum出版社。
Newton, J. R., Ellsworth, C., Miyakawa, T., Tonegawa, S., & Sur, M. (2004). 通过听觉通路加速视觉提示的条件性恐惧。自然神经科学,7,968-973。
Nishimori, K., Takayanagi, Y., Yoshida, M., Kasahara, Y., Young, L. J., & Kawamata, M. (2008). 敲除小鼠揭示的催产素受体功能新方面:社会性行为和能量平衡控制。脑研究进展,170,79-90。
Nocjar, C., & Panksepp, J. (2002). 慢性间歇性安非他明预处理增强未来对药物、食物和性奖赏的食欲行为:与环境变量的相互作用。行为脑研究,128,189-203。
Norberg, M. M., Krystal, J. H., & Tolin, D. F. (2008). D-环丝氨酸和恐惧消退及暴露疗法促进的荟萃分析。生物精神病学,63,1118-1126。
Normansell, L., & Panksepp, J. (1990). 吗啡和纳洛酮对幼年大鼠玩耍奖赏空间辨别的影响。发展心理生物学,23,75-83。
Normansell, L., & Panksepp, J. (2011). 谷氨酸能对分离痛苦的调节:兴奋性氨基酸在雏鸡中的深刻情绪效应。神经科学与生物行为评论,35,1890-1901。
Northoff, G. (2004). 脑的哲学:脑问题。阿姆斯特丹:John Benjamins出版社。
Northoff, G. (2007). 抑郁症中自我的精神病理学和病理生理学—神经精神病学假说。情感障碍杂志, 104, 1–14.
Northoff, G. (2011). 实践中的神经精神分析:大脑、自我和客体。纽约:牛津大学出版社。
Northoff, G., & Bermpohl, F. (2004). 皮层中线结构与自我。认知科学趋势, 8, 102–107.
Northoff, G., Heinzel, A., de Greck, M., Bermpohl, F., and Panksepp, J. (2006). 我们的大脑及其自我:皮层中线结构的核心作用,神经影像, 15, 440–457.
Northoff, G., & Panksepp, J. (2008). 跨物种的自我概念与皮层下-皮层中线系统。认知科学趋势, 12(7), 259–264.
Northoff, G., Qin, P., & Nakao, T. (2010). 大脑中的静息-刺激交互作用:综述。神经科学趋势, 33, 277–284.
Northoff, G., Schneider, F., Rotte, M., Matthiae, C. Tempelmann, C., Wiebking, C., Bermpohl, F., Heinzel, A., Danos, P., Heinze, H. J., Bogerts, B., Walter, M., & Panksepp, J. (2009). 不同脑区中自我相关性和情绪的差异参数调制。人脑图谱, 30, 369–382.
Northoff, G., Wiebking, C., Feinberg, T., & Panksepp, J. (2011). 重度抑郁症的静息态假说—系统障碍的转化性皮层下-皮层框架。神经科学与生物行为评论, 35, 1929–1945.
Novak, M. A. (1979). 生命第一年被隔离的猴子的社会恢复:II. 长期评估。发展心理学, 15, 50–61.
Numan, M. (1990). 母性行为的神经控制。见 N. A. Krasnegor & R. S. Bridges (编),哺乳动物的养育 (页 231–259)。纽约州,纽约:牛津大学出版社。
Numan, M., & Insel, T. R. (2003). 亲代行为的神经生物学。纽约州,纽约:施普林格出版社。
Oades, R. D. (1985). 去甲肾上腺素在调谐中的作用和多巴胺在中枢神经系统信号切换中的作用。神经科学与生物行为评论, 9, 261–282.
Ogden, P., Minton, K., & Pain, C. (2006). 创伤与身体:感觉运动心理治疗方法。纽约州,纽约:W. W. 诺顿出版社。
Oldfield, R. G., & Hofmann, H. A. (2011). 单配制慈鲷鱼社会行为的神经肽调节。生理学与行为, 102, 296–303.
Olds, J. (1977). 驱力与强化:下丘脑功能的行为研究。纽约州,纽约:雷文出版社。
Olds, J., Disterhoft, J., Segal, M., Kornblith, C., & Hirsch, R. (1972). 通过测量条件反应潜伏期绘制的大鼠大脑学习中枢图。神经生理学杂志, 35, 202–219.
Olds, J., & Milner, P. (1954). 隔膜区和大鼠大脑其他区域的电刺激产生的正强化。比较与生理心理学杂志, 47, 419–427.
Olmstead, M. C., & Franklin, K. B. (1997). 吗啡条件性位置偏好的发展:微注射到各种中枢神经系统部位的效果。行为神经科学, 111, 1324–1334.
O’Shea, K. (2009). EMDR友好的成人和儿童准备方法,见 R. Shapiro (编),EMDR解决方案II:针对抑郁症、饮食障碍、表现及更多 (页 289–312)。纽约州,纽约:W. W. 诺顿出版社。
Overton, D. A. (1991). 状态依赖性学习和药物辨别效应的历史背景。行为药理学, 2, 253–264.
Pahlavan, F. (编). (2010). 愤怒的多重面向:发怒还是恢复公正? 纽约州,纽约:诺瓦科学出版社。
Panksepp, J. (1971). 下丘脑电刺激在白化大鼠中引发的攻击行为。生理学与行为, 6, 311–316.
Panksepp, J. (1973). 大鼠进食模式的重新分析。比较与生理心理学杂志, 82, 78–94.
Panksepp, J. (1981a). 脑阿片类物质:麻醉品和社会依赖的神经化学基质。见 S. Cooper (编),精神药理学理论进展 (页 149–175)。伦敦:学术出版社。
Panksepp, J. (1981b). 行为的下丘脑整合:奖励、惩罚和相关的心理生物学过程。见 P. J. Morgane 和 J. Panksepp (编),下丘脑手册,第3卷,A部分。下丘脑的行为研究 (页 289–487)。纽约州,纽约:马塞尔·德克尔出版社。
Panksepp, J. (1981c). 大鼠游戏的个体发育。发展心理生物学, 14, 327–332.
Panksepp, J. (1982). 走向情绪的一般心理生物学理论。行为与脑科学, 5, 407–467.
Panksepp, J. (1985). 情绪变化。见 P. J. Vinken, G. W. Bruyn, & H. L. Klawans (编),临床神经病学手册(修订版)。第1卷 (45):临床神经心理学 (页 271–285)。阿姆斯特丹:爱思唯尔科学出版社。
Panksepp, J. (1988). 脑情绪回路与精神病理学。见 M. Clynes 和 J. Panksepp (编),情绪与精神病理学 (页 37–76)。纽约州,纽约:普列姆出版社。
Panksepp, J. (1990a). 情绪-认知界面的灰色地带:评论。认知与情绪, 4, 289–302.
Panksepp, J. (1990b). 恐惧的心理神经学:进化视角和动物模型在理解人类焦虑中的作用。见 R. Noyes (编),焦虑手册,焦虑的神经生物学 (页 3–58)。阿姆斯特丹:爱思唯尔/北荷兰生物医学出版社。
Panksepp, J. (1991). 情感神经科学:情绪神经生物学研究的概念框架。见 K. Strongman (编),国际情绪研究评论 (页 59–99)。奇切斯特,英国:威利出版社。
Panksepp, J. (1992). 催产素对情绪过程的影响:分离痛苦、社会联结及与精神疾病的关系。纽约科学院年鉴, 652, 243–252.
Panksepp, J. (1993). 粗野打闹游戏:一种基本的大脑过程。载于 K. B. MacDonald (主编),《父母与儿童的游戏》(pp. 147–184)。纽约州奥尔巴尼:纽约州立大学出版社。
Panksepp, J. (1998a). 《情感神经科学:人类和动物情绪的基础》。纽约:牛津大学出版社。
Panksepp, J. (1998b). 意识的边缘下基质:情感状态与自我的进化起源。《意识研究杂志》,5,566–582。
Panksepp, J. (1999). 从精神分析和神经科学视角看情绪:统合性练习及相关评论。《神经精神分析》,1,15–89。
Panksepp, J. (2000). 情绪的神经动力学:进化-神经发育观点。载于 M. D. Lewis & I. Granic (主编),《情绪、自组织与发展》(pp. 236–264)。纽约:剑桥大学出版社。
Panksepp, J. (2002). MacLean的遗产与情绪研究的一些现代趋势。载于 G. A. Cory, Jr. & R. Gardner, Jr. (主编),《Paul MacLean的进化神经行为学》(pp. ix–xxvii)。康涅狄格州韦斯特波特:Praeger出版社。
Panksepp, J. (2003a). 在情感、行为和认知神经科学的交界处:解码大脑的情感感受。《大脑与认知》,52,4–14。
Panksepp, J. (2003b). 对其他动物”分离哭声”的拟人化分析能否帮助我们了解人类社会丧失的情感本质?《心理学评论》,110,376–388。
Panksepp, J. (2003c). 感受社会丧失的痛苦。《科学》,302,237–239。
Panksepp, J. (主编) (2004). 《生物精神病学教科书》。新泽西州霍博肯:Wiley出版社。
Panksepp, J. (2005a). 情感意识:动物和人类的核心情感感受。《意识与认知》,14,19–69。
Panksepp, J. (2005b). 论核心情感的具身神经本质。《意识研究杂志》,12,158–184。
Panksepp, J. (2006a). 进化精神病学中的情感内表型。《神经精神药理学与生物精神病学进展》,30,774–784。
Panksepp, J. (2006b). 哺乳动物大脑的核心情感系统:人类情绪的基本基质。载于 J. Corrigall, H. Payne, & H. Wilkinson (主编),《关于身体:在心理治疗中使用具身心智》(pp. 14–32)。英国霍夫:Routledge出版社。
Panksepp, J. (2006c). 论社会痛苦、支持和共情的神经进化本质。载于《痛苦:关于其本质及其研究方法论的新论文》。M. Aydede (主编) pp. 367–387。马萨诸塞州剑桥:MIT出版社。
Panksepp J. (2007a). 情感意识。载于 M. Velmans 和 S. Schneider (主编),《布莱克威尔意识研究手册》(pp. 114–129)。马萨诸塞州莫尔登:Blackwell出版社。
Panksepp, J. (2007b). 游戏能否减轻ADHD并促进社会大脑的构建?《加拿大儿童与青少年精神病学会杂志》,16,57–66。
Panksepp, J. (2007c). 笑声和社会喜悦的神经进化来源:在实验室大鼠中模拟原始人类笑声。《行为脑研究》,182,231–244。
Panksepp, J., (2007d). 情感心理学的神经化:基于评估的建构主义和基本情绪理论如何共存。《心理科学视角》,2,281–296。
Panksepp, J. (2008a). 情感大脑与核心意识:神经活动如何产生情感感受?载于 M. Lewis, J. M. Haviland, & L. F. Barrett (主编),《情绪手册》(pp. 47–67)。纽约:Guilford出版社。
Panksepp, J. (2008b). 语言的力量可能存在于情感的力量中。《整合生理与行为科学》,42,47–55。
Panksepp, J. (2008c). 模拟哺乳动物大脑的原始情感心智:关于精神生活情感感受的赋格及对人工智能机器人的启示。载于 D. Dietrich, G. Fodor, G. Zucker, & D. Bruckner (主编),《模拟心智:技术性神经精神分析方法》(pp. 149–177)。纽约:Springer出版社。
Panksepp, J. (2009a). 大脑情感系统与精神生活的品质:从情感的动物模型到心理治疗的启示。载于 D. Fosha, D. J. Siegel, & M. F. Solomon (主编),《情绪的疗愈力量:情感神经科学、发展与临床实践》(pp. 1–26)。纽约:W. W. Norton出版社。
Panksepp, J. (2009b). 情感意识的非还原论物理主义解释。载于 S. J. Wood, N. B. Allen, & C. Pantelis (主编),《精神疾病的神经心理学》(pp. 399–407)。英国剑桥:剑桥大学出版社。
Panksepp, J. (2009c). 初级过程情感与大脑催产素。《生物精神病学》,65,725–727。
Panksepp, J. (2009/2010). 音乐和语言进化的情感先决条件。载于《音乐科学》“音乐与进化”特刊,pp. 229–259。
Panksepp, J. (2010a). 动物的情感意识:维度与初级过程情绪方法的视角。《英国皇家学会会刊,生物科学》,77,2905–2907。
Panksepp, J. (2010b). 情感大脑心智的情感神经科学:进化视角及对理解抑郁症的启示。《临床神经科学对话》,12,533–545。
Panksepp, J. (2010c). 嫉妒的进化来源:解决基本问题的跨物种方法。载于 S. L. Hart & M. Legerstee (主编),《嫉妒手册:理论、原理与多学科方法》(pp. 101–120)。纽约:Wiley-Blackwell出版社。
Panksepp, J. (2010d). 作为理解游戏之门的大脑科学:对Jaak Panksepp的访谈。《美国游戏杂志》,2,245–277。
Panksepp, J. (2011a). 动物社交丧失的神经生物学:解开人类精神痛苦之谜的一些关键。见 G. MacDonald & L. A. Jensen-Campbell (编), 社交痛苦:丧失和排斥的神经心理学和健康影响 (第 11–51 页)。华盛顿特区:美国心理学会。
Panksepp, J. (2011b). 跨物种情感神经科学对人类及相关动物原始情感体验的解码。PLoS ONE 6(8): e21236。
Panksepp, J. (2011c). 哺乳动物大脑的基本情绪回路:动物有情感生活吗?神经科学与生物行为评论, 35, 1791–1804。
Panksepp, J., Asma, S., Curran, G, Gabriel, R. (2012). 认知科学学会会议”情感神经科学的哲学意义”研讨会,意识研究杂志, 19, 6–48。
Panksepp, J., Bean, N. J., Bishop, P., Vilberg, T., and Sahley, T. L. (1980). 阿片阻断与雏鸟的社交安慰。药理学生物化学与行为, 13, 673–683。
Panksepp, J., & Beatty, W. W. (1980). 大鼠的社交剥夺与游戏。行为与神经生物学, 30, 197–206。
Panksepp, J., & Bekkedal, M. (1997). 神经肽与大脑焦虑的多样性。意大利精神病理学杂志, 1, 18–27。
Panksepp, J., & Bernatzky, G. (2002). 情感声音与大脑:音乐欣赏的神经情感基础。行为过程, 60, 133–155。
Panksepp, J., & Bishop, P. (1981). 大鼠大脑中(3H)双吗啡(diprenorphine)结合的放射自显影图谱:社交互动的影响。脑研究通报, 7, 405–410。
Panksepp, J., & Burgdorf, J. (1999). 大鼠会笑?嬉戏挠痒引发幼年啮齿动物的高频超声鸣叫。见 S. Hameroff, D. Chalmers, & A. Kazniak (编), 走向意识科学 III (第 231–244 页)。剑桥,马萨诸塞州:麻省理工学院出版社。
Panksepp, J., & Burgdorf, J. (2000). 大鼠对条件性和非条件性挠痒诱导奖赏的 50 千赫鸣叫(笑声?)反应:社交居住和遗传变量的影响。行为脑研究, 115, 25–38。
Panksepp, J., & Burgdorf, J. (2003). “笑”的大鼠与人类快乐的进化先例?生理学与行为, 79, 533–547。
Panksepp, J., Burgdorf, J., Gordon, N., & Turner, C. (2002). 使用哌醋甲酯(methylphenidate)治疗多动症可能使大脑的欲望基质敏感化:来自动物模型对药物滥用潜力变化的启示。意识与情感, 3, 7–19。
Panksepp, J., Burgdorf, J., Turner, C., & Gordon, N. (2003). 用大鼠单侧额叶皮层损伤模拟多动症型唤醒及游戏疗法的有益效果。大脑与认知, 52, 97–105。
Panksepp, J., & Crepeau, L. (1990). 双重嗅觉系统的选择性损伤与猫气味减弱的幼年大鼠游戏打斗。攻击行为, 16, 130–131。
Panksepp. J., Crepeau, L., & Clynes, M. (1987). 促肾上腺皮质激素释放因子(CRF)对幼年游戏中分离痛苦的影响。社会神经科学摘要, 13, 1320。
Panksepp, J., Fuchs, T., & Iacabucci, P. (2011). 哺乳动物情感体验的基础神经科学:皮层下恐惧回路的案例及其对临床焦虑的启示。应用动物行为科学, 129, 1–17。
Panksepp, J., & Gordon, N. (2003). 人类情感的本能基础:笑与哭的情感成像。意识与情感, 4, 197–206。
Panksepp, J., & Harro, J. (2004). 生物精神病学和情感精神药理学中神经肽的未来:目标与策略。见 J. Panksepp (编), 生物精神病学教科书 (第 627–660 页)。纽约,纽约州:威利出版社。
Panksepp, J., Herman, B., Conner, R., Bishop, P., & Scott, J. P. (1978). 社交依恋的生物学:阿片类物质缓解分离痛苦。生物精神病学, 13(5), 607–618。
Panksepp, J., Herman, B. H., Vilberg, T., Bishop, P., & DeEskinazi, F. G. (1980). 内源性阿片类物质与社交行为。神经科学与生物行为评论, 4, 473–487。
Panksepp, J. B., & Huber, R. (2004). 小龙虾行为的行为学分析:一个测量精神兴奋剂奖赏特性的新无脊椎动物系统。行为脑研究, 153, 171–180。
Panksepp, J., Jalowiec, J., DeEskinazi, F. G., & Bishop, P. (1985). 阿片类物质与幼年大鼠的游戏优势。行为神经科学, 99, 441–453。
Panksepp, J., Knutson, B., & Burgdorf, J. (2002). 情感大脑系统在成瘾中的作用:神经进化视角。成瘾, 97, 459–469。
Panksepp, J., Lensing, P., Leboyer, M., & Bouvard, M. P. (1991). 纳曲酮(Naltrexone)和其他自闭症潜在的新药理学治疗。脑功能障碍, 4, 281–300。
Panksepp, J., & Miller, A. (1996). 情感与衰老的大脑:遗憾与补救。见 C. Magai & S. H. McFadden (编), 衰老手册 (第 3–26 页)。纽约,纽约州:学术出版社。
Panksepp, J., & Moskal, J. (2008). 多巴胺与寻求(SEEKING):皮层下”奖赏”系统与食欲性冲动。见 A. Elliot (编), 接近与回避动机手册 (第 67–87 页) 纽约,纽约州:泰勒与弗朗西斯出版社。
Panksepp, J., Moskal, J., Panksepp, J. B., & Kroes, R. (2002). 进化心理学中的比较方法:分子神经科学遇见心智。神经内分泌学通讯, 23 (增刊 4): 105–115。
Panksepp, J., Nelson, E., & Bekkedal, M. (1997). 介导社交分离痛苦和社交奖赏的大脑系统。进化先例和神经肽中介。纽约科学院年鉴, 807, 78–100。
Panksepp, J., Newman, J. D., & Insel, T. R. (1992). 分析大脑分离痛苦系统中的关键概念问题。见 K. T. Strongman (编), 情感研究国际评论,第 2 卷 (第 51–72 页)。奇切斯特,英国:威利出版社。
Panksepp, J., Normansell, L., Cox, J. F., Crepeau, L. J., & Saks, D. S. (1987). 社会性玩耍的精神药理学。见 B. Oliver, J. Mos, & P. F. Brain (编), 动物和人类竞争行为的内药理学 (pp. 132–144). Dordrecht: Martinus Nijhoff.
Panksepp, J., Normansell, L. A., Cox, J. F., and Siviy, S. (1994). 新生儿去皮层对幼年大鼠社会性玩耍的影响。生理学与行为, 56, 429–443.
Panksepp, J., Normansell, L., Herman, B., Bishop, P., & Crepeau, L. (1988). 分离痛苦叫声的神经和神经化学控制。见 J. D. Newman (编), 哺乳动物发声的生理控制. New York, NY: Plenum.
Panksepp, J., & Northoff, G. (2009). 跨物种核心自我:通过皮层下-皮层中线网络内的自我相关处理产生的主动文化和神经生态主体。意识与认知, 18, 193–215.
Panksepp, J. & Panksepp, J. B. (2000). 进化心理学的七宗罪。进化与认知, 6, 108–131.
Panksepp, J., Sacks, D. S., Crepeau, L., & Abbott, B. B. (1991). 大脑恐惧系统的心理和神经生物学。见 M. R. Denny (编), 厌恶事件与行为 (pp. 7–59). New York, NY: Lawrence Erlbaum.
Panksepp, J. & Scott, E. (2012). 对打闹玩耍、社会发展和注意力缺陷多动障碍(ADHD)的反思。见 A. L. Meyer and T. P. Gullotta (编), 跨越生命周期的身体活动,儿童和家庭生活中的问题 (印刷中). New York: Springer.
Panksepp, J., Siviy, S., & Normansell, L. (1984). 玩耍的心理生物学:理论和方法论视角。神经科学和生物行为评论, 8, 465–492.
Panksepp, J., Siviy, S. M., & Normansell, L. A. (1985). 大脑阿片类物质和社会情绪。见 M. Reite and T. Fields (编), 依恋与分离的心理生物学 (pp. 3–49). New York, NY: Academic Press.
Panksepp, J., & Trevarthen, C. (2009). 音乐的心理生物学:音乐性表达中的动机冲动和情绪,以及对音乐的共情情绪反应。见 S. Malloch and C. Trevarthen (编), 交流性音乐性 (pp. 105–146). Cambridge, UK: Cambridge University Press.
Panksepp, J., & Trowill, J. A. (1967a). 口内自我注射:I. 强化延迟对消退抵抗的影响及其对自我刺激的意义。心理计量学科学, 9, 407–408.
Panksepp, J., and Trowill, J. A. (1967b). 口内自我注射:II. 用常规奖励模拟自我刺激现象。心理计量学科学, 9, 405–406.
Panksepp, J., Vilberg, T., Bean, N. J., Coy, D. H., & Kastin, A. J. (1978). 阿片样肽减少雏鸡的痛苦发声。脑研究通报, 3(6), 663–667.
Panksepp, J., & Watt, J. (2011). 抑郁为什么会痛?祖先原始过程分离痛苦(PANIC)和大脑奖励(SEEKING)过程减弱在抑郁情感发生中的作用。精神病学,人际和生物过程, 74, 5–14.
Panksepp, J., & Zellner, M. (2004). 走向基于神经生物学的攻击统一理论。国际社会心理学评论, 17, 37–61.
Panzica, G. C., Aste, N., Castagna, C., Viglietti-Panzica, C., & Balthazart, J. (2001). 类固醇诱导的鸟类大脑性二态性血管加压素能神经支配的可塑性:行为影响。脑研究评论, 37, 178–200.
Papes, F., Logan, D. W., & Stowers. L. (2010). 犁鼻器通过检测蛋白质信息素同源物介导物种间防御行为。细胞, 141, 692–703.
Parada, M., Chamas, L., Censi, S., Coria-Avila, G., & Pfaus, J. G. (2010). 阴蒂刺激在大鼠中诱导条件性位置偏好和Fos激活。激素与行为, 57, 112–118.
Passingham, R. (1993). 额叶和自主行动. New York, NY: Oxford University Press.
Paul, A. M. (2010). 起源:出生前九个月如何塑造我们余生. New York, NY: Simon and Schuster.
Paulsen, S. (2009). 透过创伤和分离的眼睛看:EMDR治疗师和来访者图解指南. Charleston, SC: Booksurge LLC.
Peciña, S., Smith, K., & Berridge, K. C. (2006). 大脑中的享乐热点。神经科学家, 12, 500–511.
Pedersen, C. A. (2004). 社会联结的生物学方面和人类暴力的根源。纽约科学院年鉴, 1036, 106–127.
Pedersen, C. A., Ascher, J. A., Monroe, Y. L., & Prange, A. J. (1982). 催产素在处女雌鼠中诱导母性行为。科学, 216, 648–649.
Pedersen, C. A., Caldwell, J. D., Peterson, G., Walker, C. H., & Mason, G. A. (1992). 催产素激活大鼠的母性行为。纽约科学院年鉴, 652, 58–69.
Pedersen, C. A., Vadlamudi, S. V., Boccia, M. L., & Amico, J. A. (2006). 未产催产素敲除小鼠的母性行为缺陷。基因、大脑与行为, 5, 274–281.
Pellis, S. M., & McKenna, M. (1995). 大鼠在打闹玩耍中觉得什么有奖励性?使用药物诱导的非玩耍伙伴进行的分析。行为脑研究, 68, 65–73.
Pellis, S. M., & Pellis V. C. (1987). 在实验室大鼠中,打闹玩耍与严肃打斗在攻击目标和战斗策略上都有所不同。攻击行为, 13, 227–242.
Pellis, S. M., & Pellis V. C. (2009). 爱玩耍的大脑:冒险探索神经科学的极限. Oxford, UK: Oneworld.
Perecman, E. (编). (1987). 重访额叶. New York, NY: IRBN Press.
Pessoa, L. (2008). 关于情绪和认知的关系。自然神经科学评论, 9, 148–158.
Pfaff, D. W. (1999). 驱力:性行为的神经生物学和分子机制. 剑桥,马萨诸塞州:麻省理工学院出版社。
Pfaff, D. (2006). 大脑唤醒与信息论. 剑桥,马萨诸塞州:哈佛大学出版社。
Pfaus, J. G., Kippin, T. E., & Coria-Avila, G. (2003). 动物模型能告诉我们关于人类性反应的什么信息?性研究年度评论, 14, 1–63.
Phillips, A. G., McDonald, A. C., & Wilkie, D. M. (1981). 神经阻断药物对脑刺激奖赏信号的自动塑形反应的干扰。药理学、生物化学与行为, 14, 543–548.
Phoenix, C., Goy, R., Gerall, A., & Young, W. (1959). 产前给予丙酸睾酮对雌性豚鼠交配行为相关组织的组织化作用。内分泌学, 65, 369–382.
Pincus, D., Kose, S., Arana, A., Johnson, K., Morgan, P. S., Borckardt, J., Herbsmand, T., Hardaway, F., George, M. S., Panksepp, J., & Nahas, Z. (2010). 催产素对抑郁和健康受试者归因他人心理活动的相反效应:一项双盲安慰剂对照功能磁共振成像研究。精神病学前沿, 1, 134. doi: 10.3389/fpsyt.2010.00134
Pincus, J. H. (2001). 基本本能:是什么让杀手杀人. 纽约,纽约州:W. W. 诺顿出版社。
Pincus, J. H. (1981). 暴力与癫痫。新英格兰医学杂志, 305(12), 696–698.
Pitman, R. K., van der Kolk, B. A., Orr, S. P., & Greenberg, M. S. (1990). 创伤后应激障碍中战斗相关刺激的纳洛酮可逆性镇痛反应。一项初步研究。普通精神病学档案, 47, 541–544.
柏拉图。(1941). 理想国 (B. Jowett编). 纽约,纽约州:现代文库。
Poeck, K. (1969). 与脑损伤相关的情绪障碍的病理生理学。载于P. J. Vinken & G. W. Bruyb (编), 临床神经学手册 (第3卷, 第343–367页). 阿姆斯特丹:北荷兰出版社。
Popik, P., Vetulani, J., & Van Ree, J. M. (1992). 低剂量催产素促进大鼠的社会识别。精神药理学, 106, 71–74.
Porges, S. W. (2009a). 多迷走神经理论:对自主神经系统适应性反应的新见解。克利夫兰诊所医学杂志, 76 (增刊2), S86–90.
Porges, S. (2009b). 身体与大脑在情感感知和表达中的相互影响。载于D. Fosha, D. J. Siegel, & M. F. Solomon (编), 情感的疗愈力量:情感神经科学、发展与临床实践 (第27–39页). 纽约,纽约州:W. W. 诺顿出版社。
Potegal, M., & Einon, D. (1989). 幼年期被剥夺玩耍打斗经验的成年大鼠的攻击行为。发展心理生物学, 22, 159–172.
Power, T. G. (2000). 儿童和动物的游戏与探索. 马瓦,新泽西州:劳伦斯·厄尔鲍姆出版社。
Powers, A., Ressler, K. J., & Bradley, R. G. (2009). 友谊对童年虐待和抑郁影响的保护作用。抑郁与焦虑, 26, 46–53.
Preter, M. & Klein, D. F. (2008). 恐慌、窒息错误警报、分离焦虑和内源性阿片类物质。神经精神药理学和生物精神病学进展, 32, 603–612.
Preter, M., Lee, S. H., Petkova, E., Vannucci, M., Kim, S., & Klein, D. F. (2011). 正常受试者纳洛酮先于乳酸输注的对照交叉研究;呼吸和主观反应:与内源性阿片系统、窒息错误警报理论和童年父母丧失的关系。心理医学, 41, 385–393.
Previc, F. H. (2009). 人类进化和历史中的多巴胺能心智. 剑桥,英国:剑桥大学出版社。
Propper, C., Moore, G. A., Mills-Koonce, W. R., Halpern, C. T., Hill-Soderlund, A. L., Calkins, S. D., Mary Anna Carbone, M. A., & Cox, M. (2008). 基因-环境对婴儿迷走神经反应性发展的贡献:多巴胺与母亲敏感性的相互作用。儿童发展, 79, 1377–1394.
Provine, R. R. (2000). 笑声:一项科学调查. 纽约,纽约州:维京出版社。
Pryor, K. (2005). 狗的响片训练。沃尔瑟姆,马萨诸塞州:阳光布鲁克斯出版公司。
Pulver, S. E. (1995). 精神分析过程与治疗改变机制。载于B. Moore & B. Fine (编), 精神分析,主要概念. 纽黑文:耶鲁大学出版社。
Ragland, J. D., Yoon, J., Minzenberg, M. J., & Carter, C. S. (2007). 精神分裂症认知障碍的神经影像学:寻找病理生理机制。国际精神病学评论, 19, 417–427.
Raichle, M. E., MacLeod, A. M., Snyder, A. Z., Powers, W. J., Gusnard, D. A., & Shulman, G. L. (2001). 大脑功能的默认模式。美国国家科学院院刊, 98, 676–682.
Redgrave, P., Prescott, T. J., & Gurney, K. (1999). 短潜伏期多巴胺反应是否太短而无法发出奖赏误差信号?神经科学趋势, 22, 146–151.
Reddy, V. (2008). 婴儿如何认识心智. 剑桥,马萨诸塞州:哈佛大学出版社。
Reuter, M., Panksepp, J., Schnabel, N., Kellerhoff, N., Kempel, P., & Hennig, J. (2005). 人格与创造力的生物学标记。欧洲人格杂志, 19, 83–95.
Richardson, D. E., & Akil, H. (1977). 通过电刺激大脑减轻人类疼痛,第1部分:导水管周围和脑室周围部位的急性给药。神经外科杂志, 47, 178–183.
Riches, D. (1974). 内特西利克爱斯基摩人:选择性女婴杀害的特殊案例。民族学, 13, 351–361.
Rifkin, J. (2009). 共情文明:在危机世界中奔向全球意识. 纽约,纽约州:杰里米·P·塔彻/企鹅出版社。
Rilling, J. K., Winslow, J. T., & Kilts, C. D. (2004). 优势雄性恒河猴配偶竞争的神经相关性。生物精神病学, 56, 364–375.
Rilling, J. K., Barks, S. K., Parr, L. A., Preuss, T. M., Faber, T. L., Pagnoni, G., Bremner, J. D., & Votaw, J. R. (2007). 人类和黑猩猩静息状态脑活动的比较。美国国家科学院院刊, 104, 17146–17151.
Riters, L. V. (2010). 阿片类物质参与鸟鸣动机的证据。化学神经解剖学杂志, 39, 141–150.
Riters, L. V. (2011). 寻求快乐与鸟鸣。神经科学与生物行为评论, 35, 1837–1845.
Riters, L. V., & Panksepp, J. (1997). 血管加压素对雄性日本鹌鹑攻击行为的影响。纽约科学院年鉴, 807, 478–480.
Rizzolatti, I., & Sinigaglia, C. (2008). 大脑中的镜像:我们的心智如何分享行动、情绪和体验。英国牛津:牛津大学出版社。
Roberts, V. J., & Cox, V. C. (1987). 在位置偏好范式中使用背侧中央灰质刺激进行主动回避条件反射。心理生物学, 15, 167–170.
Robinson, T. E., & Berridge, K. C. (1993). 药物渴求的神经基础:成瘾的激励敏化理论。脑研究评论, 19, 247–291.
Rogers, C. (1961). 成为一个人。波士顿:霍顿·米夫林出版社。
Rogers, C. (1980). 存在的方式。波士顿:霍顿·米夫林出版社。
Rolls, E. T. (1999). 大脑与情绪。英国牛津:牛津大学出版社。
Rolls, E. T. (2005). 情绪解析。英国牛津:牛津大学出版社。
Romanes, G. (1882). 动物智能。伦敦:劳特利奇与基根·保罗出版社。
Roosevelt, F. D. (1933年3月4日). 首次就职演说。[在华盛顿特区美国国会大厦发表的演讲]。
Rosenblatt, J. S. (1967). 大鼠母性行为的非激素基础。科学, 156(3781), 1512–1514.
Rosenblatt, J. S. (1990). 母性行为生理学研究的里程碑,特别关注大鼠。载于 N. A. Krasnegor 和 R. S. Bridges(编),哺乳动物的养育(第40–60页)。纽约:牛津大学出版社。
Ross, M. D., Owren, M. J., & Zimmermann, E. (2009). 重建类人猿和人类笑声的进化。当代生物学, 19, 1106–1111.
Ross, A. G., Shochet, I. M., & Bellair, R. (2010). 社交技能和学校联结在青春期前抑郁症状中的作用。临床儿童与青少年心理学杂志, 39, 269–275.
Ross, H. E., & Young, L. J. (2009). 催产素与调节社会认知和亲和行为的神经机制。神经内分泌学前沿, 30, 534–547.
Rossi, J., III, & Panksepp, J. (1992). 自我刺激嗅探与脑刺激嗅探之间关系的分析。生理学与行为, 51, 805–813.
Rossignol, D. A. (2009). 自闭症谱系障碍的新颖和新兴治疗方法:系统综述。临床精神病学年鉴, 21, 213–236.
Rosvold, H. E., Mirsky, A. F., and Pribram, K. H. (1954). 杏仁核切除术对猴子社会行为的影响。比较生理学与心理学杂志, 47, 173–178.
Rubin, L. H., Carter, C. S., Drogos, L., Pournajafi-Nazarloo, H., Sweeney, J. A., & Maki, P. M. (2010). 外周催产素与精神分裂症症状严重程度降低相关。精神分裂症研究, 124, 13–21.
Rumpel, S., LeDoux, J., Zador, A., & Malinow, R. (2005). 突触后受体运输是联想学习的基础。科学, 308, 83–88.
Russell, J. S. (1994). 面部表情的情绪识别是否具有普遍性?跨文化研究综述。心理学通报, 115, 102–141.
Russell, J. A. (2003). 核心情感与情绪的心理建构。心理学评论, 110, 145–172.
Russell, J. A. (2009). 情绪、核心情感与心理建构。认知与情绪, 23, 1259–1283.
Russell, J. A., Rosenberg, E. L., & Lewis, M. D.(编)(2011). 基本情绪理论,情绪评论, 3(4)。
Ryan, R.M. & Deci, E.L. (2006). 自我调节与人类自主性问题:心理学需要选择、自我决定和意志吗?人格杂志, 74, 1557–1585.
Sacks, O. (1973). 觉醒。纽约:达顿出版社。
Salamone, J. D. (1994). 伏隔核在食欲性和厌恶性动机中的参与。行为脑研究, 61, 117–133.
Salamone, J. D., Correa, M., Farrar, A. M., Nunes, E. J., & Pardo, M. (2009). 多巴胺、行为经济学与努力。行为神经科学前沿, 3, 13.
Savic, I., Heden-Blomqvist, E., & Berglund, H. (2009). 人类信息素信号转导:从嗅觉丧失中可以学到什么。人脑图谱, 30, 3057–3065.
Schachter, S., & Singer, J. (1962). 情绪状态的认知、社会和生理决定因素。心理学评论, 69, 379–399.
Schaefer, C. E., & Kaduson, H. G.(编)(2006). 当代游戏疗法:理论、研究与实践。纽约:吉尔福德出版社。
Schiff, N. D. (2007). 全局意识障碍。载于 M. Velmans 和 S. Schneider(编),布莱克韦尔意识指南(第589–604页)。英国牛津:布莱克韦尔出版社。
Schiller, D., Monfils, M. H, Raio, C. M., Johnson, D. C., Ledoux, J. E., & Phelps, E. A. (2010). 使用再巩固更新机制防止人类恐惧的回归。自然, 463, 49–53.
Schlaepfer, T. E., Cohen, M. X., Frick, C., Kosel, M., Brodesser, D., Axmacher, N., Joe, A. Y., Kreft, M., Lenartz, D., & Sturm, V. (2008). 对奖赏回路的深部脑刺激缓解难治性重度抑郁症的快感缺失。神经精神药理学, 33(2), 368–377.
Schnall, S., & Laird, J. D. (2003). 保持微笑:面部表情和姿势对情绪体验和记忆的持久影响。认知与情绪, 17, 787–797.
Schoene-Bake, J.-C., Parpaley, Y., Weber, B., Panksepp, J. Hurwitz, T. A., & Coenen, V. A. (2010). 抑郁症历史性病灶手术的纤维束成像分析。神经精神药理学, 35, 2553–2563。
Schore, A. (1994). 情感调节与自我的起源:情绪发展的神经生物学。新泽西州希尔斯代尔:劳伦斯·厄尔鲍姆出版社。
Schore, A. N. (2001). 安全依恋关系对右脑发育、情感调节和婴儿心理健康的影响。婴儿心理健康杂志, 22, 7–66。
Schore, A. (2003a). 情感失调与自我障碍。纽约州纽约市:W. W. 诺顿出版公司。
Schore, A. (2003b). 情感调节与自我修复。纽约州纽约市:W. W. 诺顿出版公司。
Schultz, W. (2006). 行为理论与奖赏的神经生理学。心理学年度评论, 57, 87–115。
Schultz, W. (2010). 奖赏价值和风险的多巴胺信号:基础与最新数据。行为脑功能, 6, 24。
Schultz, W., Apicella, P., & Ljunberg, T. (1993). 猴子多巴胺神经元在学习延迟反应任务连续步骤中对奖赏和条件刺激的反应。神经科学杂志, 13, 900–913。
Schultz, W., & Dickinson, A. (2000). 预测误差的神经元编码。神经科学年度评论, 23, 473–500。
Schultz, W., & Romo, R. (1990). 猴子中脑的多巴胺神经元:对引发即时行为反应的刺激的条件性反应。神经生理学杂志, 63, 607–617。
Scott, E. (2001). 面向有益于儿童课堂注意力的游戏项目(博士论文)。俄亥俄州鲍灵格林:鲍灵格林州立大学。
Scott, E., & Panksepp, J. (2003). 人类儿童的粗暴打闹游戏。攻击行为, 29, 539–551。
Scott, J. P. (1974). 精神药物对狗分离痛苦的影响。国际医学文摘大会系列, 359, 735–745。
Scott, J. P., & Fuller J. L. (1998). 狗的遗传学与社会行为。芝加哥:芝加哥大学出版社。
Sewards, T. V. (2004). 味觉的感觉和享乐方面的双重独立通路。大脑研究通报, 62, 271–283。
Shair, H. N. (2007). 社会介导分离反应的获得和表达。行为与大脑研究, 182, 180–192。
Shamay-Tsoory, S. G., Fischer, M., Dvash. J., Harari, H., Perach-Bloom, N., & Levkovitz, Y. (2009). 鼻内给予催产素增加嫉妒和幸灾乐祸。生物精神病学, 66, 864–870。
Shapiro, F. (2001). 眼动脱敏与再加工:基本原理、方案和程序(第2版)。纽约州纽约市:吉尔福德出版社。
Shapiro, F. (2002). EMDR引入12年后:过去和未来的研究。临床心理学杂志, 58, 1–22。
Shedler, J. (2010). 心理动力学心理治疗的疗效。美国心理学家, 65, 98–109。
Sheehan, T., & Numan, M. (2002). 雌激素、孕激素和妊娠终止改变大鼠控制母性行为的大脑区域的神经活动。神经内分泌学, 75, 12–23。
Sheldon, K., Kashdan, T., & Steger, M. (编). (2011). 设计积极心理学的未来:盘点现状与继续前进。纽约州纽约市:牛津大学出版社。
Shema, R., Sacktor, T. C., & Dufai, Y. (2007). PKM zeta抑制剂快速消除皮层中的长期记忆关联。科学, 317, 951–953。
Shewmon, D. A., Holmes, G. L., & Byrne, P. A. (1999). 先天性去皮层儿童的意识:作为自我实现预言的发育性植物状态。发育医学与儿童神经学, 41, 364–374。
Shi, C., & Davis, M. (2001). 用恐惧增强惊吓测量的恐惧条件反射中涉及的视觉通路:行为和解剖学研究。神经科学杂志, 21, 9844–9855。
Siegel, A. (2005). 攻击和愤怒的神经生物学。佛罗里达州博卡拉顿:CRC出版社。
Siegel, D. J. (2007). 正念的大脑:培养幸福感的反思与调谐。纽约州纽约市:W. W. 诺顿出版公司。
Siegel, D. J. (2010). 正念。纽约州纽约市:班坦出版社。
Sierra-Mercado, D., Padilla-Coreano, N., & Quirk, G. J. (2011). 前边缘皮层和下边缘皮层、腹侧海马和基底外侧杏仁核在条件性恐惧的表达和消退中的可分离作用。神经精神药理学, 36, 529–538。
Singewald, N. (2007). 通过挑战范式和功能映射揭示高焦虑啮齿动物改变的大脑活动处理。神经科学与生物行为评论, 31, 18–40。
Singh, D., & Randall, P. K. (2007). 美在整形外科医生的眼中:腰臀比(WHR)与女性吸引力。个性与个体差异, 43, 329–340。
Siviy, S. M. (2010). 游戏与逆境:爱玩耍的哺乳动物大脑如何抵御威胁和焦虑。美国游戏杂志, 2, 297–314。
Siviy, S. M., Harrison, K. A., & McGregor, I. S. (2006). 幼年大鼠的恐惧、风险评估和玩耍性。行为神经科学, 120, 49–59。
Siviy, S. M., & Panksepp, J. (1985). 背内侧间脑参与大鼠的幼年游戏。行为神经科学, 99, 1103–1113。
Siviy, S. M., & Panksepp, J. (1987a). 大鼠幼年游戏的感觉调节。发育心理生物学, 20, 39–55。
Siviy, S.M., & Panksepp, J. (1987b). 大鼠的幼年游戏:丘脑和脑干的参与。生理学与行为, 41, 103–114。
Skinner, B. F. (1938). 有机体的行为。纽约州纽约市:阿普尔顿-世纪-克罗夫茨出版社。
Skinner, B. F. (1948). 鸽子的”迷信”。实验心理学杂志, 38, 168–172。
Skinner, B. F. (1953). 科学与人类行为。纽约州纽约市:西蒙与舒斯特出版社。
Skolnick, P. (2009). 基于谷氨酸的抗抑郁药:20年回顾。药理学趋势, 30, 563–569.
Skrundz, M., Bolten, M., Nast, I., Hellhammer, D. H., & Meinlschmidt, G. (2011). 怀孕期间血浆催产素(oxytocin)浓度与产后抑郁发展的关联。神经精神药理学, 1–8, 在线提前发表, 2011年5月11日, doi: 10.1038/npp.2011.74.
Slade, A. & Wolf, D. P. (主编). (1994). 游戏中的儿童:意义和表征的临床与发展方法. 纽约州纽约市: 牛津大学出版社.
Slotnick, B. M. (1975). 大鼠母性行为的神经和激素基础。见 B. E. Eletheriou & R. L. Sprott (主编), 行为的激素相关性. 第2卷 (第 585–656页). 纽约州纽约市: Plenum出版社.
Smith, W. D. (1979). 希波克拉底传统. 纽约州伊萨卡: 康奈尔大学出版社.
Smotherman, W. P. & Robinson, S. R. (1992). κ阿片类物质对胎儿对乳汁反应的中介作用。行为神经科学, 106, 396–407.
Snyder, S. H. (1972). 脑内儿茶酚胺(catecholamines)作为苯丙胺精神病的介质。普通精神病学档案, 27, 169–179.
Solanto, M. V. (2000). 注意力缺陷多动障碍(AD/HD)的临床精神药理学:对动物模型的启示。神经科学与生物行为评论, 24, 27–30.
Solms, M. (2000). 做梦和快速眼动睡眠由不同的大脑机制控制。行为与脑科学, 23, 843–850.
Solms, M. (2002). 做梦:胆碱能(cholinergic)和多巴胺能假说。见 E. Perry, H. Ashton, & A. Young (主编), 意识的神经化学:心智中的神经递质. 意识研究进展 (第36卷, 第 123–131页). 阿姆斯特丹, 荷兰: John Bensamis出版社.
Solms, M., & Panksepp, J. (2010). 为什么抑郁让人感觉糟糕。见 E. K. Perry, D. Collerton, F. E. N. LeBeau, & H. Ashton (主编), 意识神经科学的新视野, (第 169–178页). 阿姆斯特丹: John Benjamins出版社.
Solms, M., & Panksepp, J. (2012). “本我”比”自我”承认的知道得更多:神经精神分析(neuropsychoanalytic)和原始意识视角看情感与认知神经科学的接口。脑科学, 2(2), 147–175.
Solms, M., & Turnbull, O. (2002). 大脑与内在世界:主观体验神经科学导论. 纽约州纽约市: Other Press出版社.
Solomon, A. (2001). 正午的恶魔:抑郁地图集. 纽约州纽约市: Scribner出版社.
Soltysik, S., & Jelen, P. (2005). 在大鼠中,叹气与解脱相关。生理学与行为, 85, 598–602.
Sparks, D. L. (1988). 神经制图学(neural cartography):上丘(superior colliculus)中的感觉和运动地图。脑行为与进化, 31, 49–56.
Spinka, M., Newberry, R. C., & Bekoff, M. (2001). 哺乳动物的游戏:为意外做训练。生物学季刊评论, 76, 141–168.
Spitz, R. A., and Wolf, K. M. (1946). 依恋性抑郁(anaclitic depression):早期儿童精神病理状况发生的探究。儿童精神分析研究, 2, 313–342.
Stein, D. J. (2009). 边缘型人格障碍:走向整合。中枢神经系统谱, 14, 352–356.
Steiner, J. E., Glaser, D., Hawilo, M. E., & Berridge, K. C. (2001). 享乐性影响的比较表达:人类婴儿和其他灵长类动物对味觉的情感反应。神经科学与生物行为评论, 25, 53–74.
Stengel, A., & Taché, Y. (2010). 促肾上腺皮质激素释放因子(corticotropin-releasing factor)信号传导和对压力的内脏反应。实验生物学与医学, 235, 1168–1178.
Stepniewska, I., Fang, P. C., & Kaas, J. H. (2005). 微刺激(microstimulation)揭示原猴婴猴(prosimian galagos)后顶叶皮层中针对不同复杂运动的专门化亚区域,美国国家科学院院刊, 102, 4878–4883.
Stepper, S., & Strack, F. (1993). 情感和非情感感觉的本体感受(proprioceptive)决定因素。人格与社会心理学, 64, 211–220.
Stern, D. N. (2004). 心理治疗和日常生活中的当下时刻(present moment). 纽约州纽约市: W. W. Norton出版社.
Sterns, F. R. (1972). 笑:生理学、病理生理学、心理学、精神病理学和发展. 伊利诺伊州斯普林菲尔德: Charles C. Thomas出版社.
Stone, K. C., LaGasse, L. L., Lester, B. M., Shankaran, S., Bada, H. S., Bauer, C. R., & Hammond, J. A. (2010). 产前物质暴露儿童的睡眠问题:母亲生活方式研究。儿科与青少年医学档案, 164, 452–456.
Strassman, R. J., Appenzeller, O., Lewy, A. J., Qualls, C. R., & Peake, G. T. (1989). 28.5英里山地赛跑后血浆褪黑素(melatonin)、β-内啡肽(beta-endorphin)和皮质醇(cortisol)的增加:与表现的关系及纳洛酮(naltrexone)无效应。临床内分泌学与代谢杂志, 69, 540–545.
Strehler, B. L. (1991). 自我在哪里?意识的神经解剖学理论。突触, 7, 44–91.
Strüber, D., Lück, M., & Roth, G. (2008). 性、攻击性和冲动控制:一个整合性解释。神经病例, 14, 93–121.
Stutz, R. M., Rossi, R. R., Hastings, L., & Brunner, R. L. (1974). 颅内刺激的可辨别性:解剖学连接性的作用。生理学与行为, 12, 69–73.
Suddendorf, T., & Corballis, M. (1997). 心理时间旅行(mental time travel)与人类心智的进化。遗传学、社会与普通心理学专著, 123, 133–167.
Sukhotinsky, I., Zalkind, V., Lu, J., Hopkins, D. A., Saper, C. B., & Devor, M. (2007). 与脑内微注射GABAA活性麻醉剂引起的意识丧失相关的神经通路。欧洲神经科学杂志, 25, 1417–1436.
Sunderland, M. (2006). 养育的科学. 纽约州纽约市: Dorling Kindersley出版社.
Suomi, S. J. (2006). 恒河猴中的风险、复原力和基因×环境交互作用。纽约科学院年鉴, 1094, 52–62.
Sur, M., & Rubinstein, J. L. (2005). 大脑皮层的模式化(patterning)和可塑性。科学, 310, 805–810.
Swain, J. E, Lorberbaum, J. P, Korse, S., & Strathearn, L. (2007). 早期亲子互动的脑基础:心理学、生理学和体内功能神经影像学研究。儿童与青少年精神病学杂志, 48, 262–287.
Szalavitz, M. & Perry, B. D. (2010). 为爱而生:为什么共情至关重要且濒临危险. 纽约,纽约州:HarperCollins出版社。
Szyf, M., McGowan, P., & Meaney, M. J. (2008). 社会环境与表观基因组。环境与分子诱变, 49, 46–60.
Takahashi, H., Matsuura, M., Yahata, N., Koeda, M., Suhara, T., & Okubo Y. (2006). 男性和女性在性背叛和情感背叛想象中表现出不同的脑激活。神经影像, 32, 1299–1307.
Talmi, D., Hurlemann, R., Patin, A., & Dolan, R. J. (2010). 双侧杏仁核损伤后的框架效应。神经心理学, 48, 1823–1827.
Tansey, K. E., Brookes, K. J., Hill, M. J., Cochrane, L. E., Gill, M., Skuse, D., Correia, C., Vicente, A., Kent, L., Gallagher, L., & Anney, R. J. (2010). 催产素受体(OXTR)在自闭症病因学中未起主要作用:遗传和分子研究。神经科学通讯, 474, 163–167.
Taylor, S. E., Klein, L. C., Lewis, B. P., Gruenewald, T. L., Gurung, R. A., & Updegraff, J. A. (2000). 女性对压力的生物行为反应:照料和结盟(tend-and-befriend),而非战或逃(fight-or-flight)。心理学评论, 107, 411–429.
Taylor, S. E., Saphire-Bernstein, S., & Seeman, T. E. (2010). 女性血浆催产素和男性血浆加压素是痛苦配偶关系的生物标记物吗?心理科学, 21, 3–7.
Teasdale, J. D., Taylor, R., & Fogarty, S. J. (1980). 诱导性兴奋-抑郁对快乐和不快乐经历记忆可及性的影响。行为研究与治疗, 18, 339–346.
Tenore, P. L. (2008). 阿片激动剂治疗的心理治疗益处。成瘾障碍杂志, 27, 49–65.
Thandeka. (2005). 施莱尔马赫的情感神学。实践神学国际杂志, 9, 197–216.
Thandeka. (2009). 美国自由神学的未来设计。美国神学与哲学杂志, 30, 72–100.
Thor, D. H., & Holloway, W. R. Jr. (1984a). 幼年大鼠社交游戏行为的发育分析。心理学会公报, 22, 587–590.
Thor, D. H. & Holloway, W. R. Jr. (1984b). 幼年大鼠的社交游戏:十年的方法学和实验研究。神经科学生物行为评论, 455–464.
Thorndike, E. L. (1911). 动物智能:实验研究. 纽约,纽约州:Macmillan。
Tinbergen, N. (1951). 本能研究. 纽约,纽约州:牛津大学出版社。
Tomasello, M. (2009). 我们为何合作. 剑桥,马萨诸塞州:麻省理工学院出版社。
Tomkins, S. S. (1962). 情感、想象、意识:第1卷. 积极情感. 纽约,纽约州:Springer。
Tomkins, S. S. (1963). 情感、想象、意识:第2卷. 消极情感. 纽约,纽约州:Springer。
Trezza, V., & Vanderschuren, L. J. (2008). 大麻素和阿片对青春期大鼠社交游戏行为的调节:不同的行为机制。欧洲神经精神药理学, 18, 519–530.
Trezza, V., & Vanderschuren, L. J. (2009). 具有不同靶选择性的花生四烯酸乙醇胺(anandamide)转运体抑制剂对青春期大鼠社交游戏行为的不同影响。药理学与实验治疗学杂志, 328, 343–350.
Tronick, E. (2007). 婴幼儿的神经行为和社会情感发展. 纽约,纽约州:W. W. Norton。
Tronson, N. C., & Tayor, J. R. (2007). 记忆再巩固的分子机制。自然神经科学评论, 8, 262–275.
Tucker, D., & Williamson, P. A. (1984). 人类自我调节中的不对称神经控制系统。心理学评论, 91, 185–215.
Tuiten, A., van Honk, J., Koppeschaar, H., Bernaards, C., Thijssen, J., & Verbaten, R. (2000). 睾酮给药对女性性唤起影响的时间进程。普通精神病学档案, 57, 149–153.
Tulving, E. (2001). 情景记忆(episodic memory)与常识:相距多远?英国皇家学会伦敦哲学汇刊B生物科学, 356, 1505–1515.
Tulving, E. (2002). 情景记忆:从心智到大脑。心理学年度评论, 53, 1–25.
Tulving, E. (2005). 情景记忆与自我意识(autonoesis):人类独有?载于H. S. Terrace & J. Metcalfe (编), 认知中缺失的环节:自我反思意识的起源 (第3–56页). 纽约,纽约州:牛津大学出版社。
Tzschentke, T. M. (2007). 用条件性位置偏爱(CPP)范式测量奖赏:过去十年的更新。成瘾生物学, 12, 227–462.
Udry, J. R. (2000). 性别建构的生物学限制。美国社会学评论, 65, 443–457.
Uhl-Bronner, S., Waltisperger, E., Martínez-Lorenzana, G., Condes Lara, M., & Freund-Mercier, M. J. (2005). 大鼠前脑和脊髓中催产素结合位点的性别二态性表达。神经科学, 135, 147–154.
Ungerstedt, U. (1971). 6-羟基多巴胺诱导黑质-纹状体多巴胺系统退化后的无饮症和无食症。斯堪的纳维亚生理学学报 (增刊367), 95–122.
Ungless, M. A. (2004). 多巴胺:显著性问题。神经科学趋势, 27, 702–706.
Uvnäs-Moberg, K. (1998). 催产素可能介导积极社会互动和情感的益处。心理神经内分泌学, 23, 819–835.
Valenstein, E. S. (1966). 强化的解剖学位点。载于E. Stellar & J. M. Sprague (编), 第1卷 生理心理学进展 (第149–190页). 纽约,纽约州:学术出版社。
Valenstein, E. S., Cox, V. C., & Kakolewski, V. C. (1970). 重新审视下丘脑在动机中的作用。心理学评论, 77, 16–31.
Vaillant, G. E. (2008). 积极情绪、灵性与精神病学实践。Mental Health, Spirituality, Mind, 6, 48–62.
Vandekerckhove, M., & Panksepp, J. (2009). 从无意识到自知意识和自传体意识的流动:对过去事物记忆和想象未来中无知的无意识和知晓的自知意识的展望。Consciousness and Cognition, 18(4), 1018–1028.
van der Kolk, B. A. (2006). 神经科学研究在PTSD中的临床意义。Annals of the New York Academy of Sciences, 1071, 277–293.
Vanderschuren, L. J. M. J. (2010). 大脑如何让玩耍变得有趣。American Journal of Play, 2, 315–337.
Vanderschuren, L. J. M. J., Niesink, R. J. M., Spruijt, B. M., & Van Ree, J. M. (1995). 吗啡对幼年大鼠社交游戏不同方面的影响。Psychopharmacology, 117, 225–231.
van Gulick, R. (2004). 意识。Stanford Encyclopedia of Philosophy. http://plato.stanford.edu/entries/consciousness/
van Honk, J., Harmon-Jones, E., Morgan, B. E., & Schutter, D. J. (2010). 社交爆发性思维:反应性攻击的三重失衡假说。Journal of Personality, 78, 67–94.
van Honk, J., & Pruessner, J. C. (2010). 精神神经内分泌成像。在Psychoneuroendocrinology, 35特刊中, 1–4.
van Honk, J., Schutter, D. J., Hermans, E. J., Putman, P., Tuiten, A., & Koppeschaar, H. (2004). 睾酮改变健康年轻女性对惩罚和奖励敏感性之间的平衡。Psychoneuroendocrinology, 29, 937–943.
van Leengoed, E., Kerker, E., & Swanson, H. H. (1987). 通过向脑室注入催产素拮抗剂抑制大鼠产后母性行为。Journal of Endocrinology, 112, 275–282.
Varela, F. J. (1999). 当下时刻意识。Journal of Consciousness Studies, 6, 111–140.
Veenema, A. H., & Neumann, I. D. (2008). 中枢血管加压素和催产素释放:复杂社会行为的调节。Progress in Brain Research, 170, 261–276.
Velmans, M. (2009). 理解意识(第2版)。纽约:Routledge出版社。
Vertes, R. P. & Kocsis, B. (1997). 控制海马θ节律的脑干-间脑-隔-海马系统。Neuroscience, 81, 893–926.
Viviani, D., & Stoop, R. (2008). 催产素和血管加压素对恐惧反应情绪表达的相反作用。收录于J. D. Neumann & R. Landgraf(编),Progress in brain research, 170, 207–220.
Viviani, D., Terrettaz, T., Magara, F., & Stoop, R. (2010). 催产素增强地西泮在大鼠中央内侧杏仁核的抑制作用。Neuropharmacology, 58, 62–68.
Voracek, M., Manning, J. T., & Dressler, S. G. (2007). 专家进行的手指比率(2D:4D)测量的可重复性和观察者间误差。American Journal of Human Biology, 19, 142–146.
Vul, E., Harris, C., Winkelman, P., & Pashler, H. (2009). fMRI情绪、人格和社会认知研究中令人困惑的高相关性。Perspectives on Psychological Science, 3, 274–290.
Vyas, A. Kim, S. K., Giacomini, N., Boothroyd, J. C., & Sapolsky, R. M. (2007). 弓形虫感染引起的啮齿动物行为变化高度特异于对猫气味的厌恶。Proceedings of National Academy of Sciences, 104, 6442–6447.
Vytal, K., & Hamann, S. (2010). 基本情绪的离散神经相关性的神经影像学支持:基于体素的元分析。Journal of Cognitive Neuroscience, 22, 2864–2885.
Wallace, B. A. (2000). 主观性的禁忌。纽约:牛津大学出版社。
Walsh, R. J., Slaby, F. J., & Posner, B. I. (1987). 催乳素从血液到脑脊液转运的受体介导机制。Endocrinology, 120, 1846–1850.
Waraczynski, M. A. (2006). 中央扩展杏仁核网络作为奖励评估的潜在回路。Neuroscience and Biobehavioral Reviews, 30, 472–496.
Ward, I. L. (1992). 性行为:围产期激素和青春期前社会因素的产物。收录于A. A. Gerall, H. Moltz, & I. L. Ward(编),性别分化,第11卷,行为神经生物学手册(第157–178页)。纽约:Plenum出版社。
Watson, D., Clark, L. A., & Tellegen, A. (1988). 积极和消极情感简短测量的开发和验证:PANAS量表。Journal of Personality and Social Psychology, 54, 1063–1070.
Watson, J. B. (1929). 行为主义。纽约:W. W. Norton出版社。
Watt, D. F. (2000). 中枢脑和丘脑皮层整合:被忽视的导水管周围灰质的贡献,Emotion and Consciousness, 1, 93–116.
Watt, D. F. (2007). 迈向共情的神经科学(附评论)。Neuro-Psychoanalysis, 9, 119–172.
Watt, D. F., & Panksepp, J. (2009). 抑郁症:一种终止分离痛苦的进化保守机制?对氨基能、肽能和神经网络视角的综述。Neuropsychoanalysis, 11, 5–104.
Watt, D. F., & Pincus, D. I. (2004). 意识的神经基础:对临床精神病学的启示。收录于J. Panksepp(编),生物精神病学教科书。霍博肯:Wiley出版社。
Wayner, M. J., Barone, F. C., & Loulis, C. C. (1981). 外侧下丘脑与附属行为。收录于P. J. Morgane and J. Panksepp(编),下丘脑手册,第3卷,B部分。下丘脑的行为研究(第107–146页)。纽约:Marcel Dekker出版社。
Weisler, A., & McCall, R. R. (1976). 探索和游戏:总结与重新定向。American Psychologist, 31, 492–508.
Welker, W. I. (1971). 游戏和探索行为的个体发生:问题的界定和新概念解决方案的探索。收录于P. P. G. Bateson & H. Moltz(编),脊椎动物行为的个体发生(第171–228页)。纽约:Academic Press出版社。
Wiegmann, D. D., Wiegmann, D. A., & Waldron, F. A. (2003). 奖励下降对大黄蜂觅食者消费行为和花朵选择的影响。生理学与行为, 79, 561–566。
Wiest, G., Lehner-Baumgartner, E., & Baumgartner, C. (2006). 双侧杏仁核选择性病变个体的恐慌发作。神经病学档案, 63, 1798–1801。
Wild, B., Rodden, F. A., Grodd, W., & Ruch, W. (2003). 笑和幽默的神经相关性。大脑, 126, 2121–2138。
Williams, G. C. (1992). 自然选择:领域、层次与挑战。纽约,纽约州:牛津大学出版社。
Wise, R. A. (1971). 下丘脑刺激位点效应的个体差异。生理学与行为, 6, 569–572。
Wise, R. A. (1982). 神经阻滞剂与操作性行为:快感缺失假说。行为与脑科学, 5, 39–87。
Wise, R. A., & Rompre, P. P. (1989). 脑多巴胺与奖励。心理学年度评论, 40, 191–225。
Wittgenstein, L. (1967). 哲学研究(G. E. M. Anscombe译)。牛津:布莱克威尔出版社。(原著出版于1953年。)
Wittgenstein, L. (1981). 逻辑哲学论。伦敦:劳特利奇出版社。(原著出版于1922年。)
Wöhr, M., Kehl, M., Borta, A., Schänzer, A., Schwarting, R. K., & Höglinger, G. U. (2009). 神经发生与情感关系的新见解:挠痒诱导发出食欲性50-kHz超声波发声的大鼠海马细胞增殖。神经科学, 163, 1024–1030。
Worthman, C. M., Plotsky, P. M., Schechter, D. S., & Cummings, C. A. (2010). 形成性体验。纽约,纽约州:剑桥大学出版社。
Wright, J., & Panksepp, J. (2011). 迈向基于情感回路的抑郁症临床前模型:背侧导水管周围灰质唤醒致敏导致大鼠正性情感持续抑制。神经科学与生物行为评论, 35, 1902–1915。
Yamamuro, T., Senzaki, K, Iwamoto, S., Nakagawa, Y., Hayashi, T., Hori, M., Sakamoto, S., Murakami, K, Shiga, T., & Urayamaa, O. (2010). 带有积极情绪的挠痒刺激增强大鼠海马齿状回的神经发生。神经科学研究, 68, 285–289。
Yamasue, H., Kuwabara, H., Kawakubo, Y., & Kasai, K. (2009). 催产素、社会脑的性别二态特征与自闭症。精神病学与临床神经科学, 63, 129–40。
Yarbus, A. L. (1967). 眼动与视觉。纽约,纽约州:普莱纳姆出版社。
Yates, A. (1978). 无羞性教育:鼓励儿童的健康性发展。纽约,纽约州:威廉·莫罗出版社。
Yeomans, J. S., Mathur, A., & Tampakeras, M. (1993). 奖励性脑刺激:激活多巴胺神经元的被盖胆碱能神经元的作用。行为神经科学, 107, 1077–1087。
Young, L. J., & Wang, Z. (2004). 配对结合的神经生物学。自然神经科学, 7, 1048–1054。
Zachar, P., & Ellis, R.(编)。(2012). 雅克·潘克塞普(Jaak Panksepp)与吉姆·罗素(Jim Russell)的情感理论。阿姆斯特丹:约翰·本杰明出版社。
Zaharna, M., Dimitriu, A., & Guilleminault, C. (2010). 发作性睡病药物治疗的专家意见。药物治疗专家意见, 11, 1633–1645。
Zellner, M., Watt, D. F., Solms, M., & Panksepp, J. (2011). 解决两个棘手精神问题的情感神经科学和神经精神分析方法:为什么抑郁症感觉如此糟糕以及成瘾者真正想要什么。神经科学与生物行为评论, 35, 2000–2008。
Zhang, D., & Raichle, M. E. (2010). 疾病与大脑的暗能量。自然神经病学评论, 6, 15–28。
Zhang, T. Y., & Meaney, M. J. (2010). 表观遗传学与基因组及其功能的环境调控。心理学年度评论, 61, 439–466。
Zhou, J.-N., Hofman, M. A., Gooren, L. J. G., & Swaab, D. F. (1995). 人类大脑的性别差异及其与易性症的关系。自然, 378, 68–70。
Zinken, J., Knoll, M., & Panksepp, J. (2008). 情感发声的普遍性与多样性。见K. Izdebski(编),人类声音中的情感,第1卷:基础(第185–202页)。圣地亚哥,加利福尼亚州:普鲁拉尔出版社。
Zubieta, J. K., Ketter, T. A., Bueller, J. A., Xu, Y., Kilbourn, M. R., Young, E. A., & Koeppe, R. A. (2003). 前扣带回和边缘系统μ-阿片神经传递对人类情感反应的调节。普通精神病学档案, 60, 1145–1153。
Zuckerman, M., & Kuhlman, D. M. (2000). 人格与冒险:共同的生物社会因素。人格杂志, 68, 999–1029。
本书印刷版中的索引已从电子书中删除。请使用您的电子阅读设备上的搜索功能查找感兴趣的术语。供您参考,印刷版索引中出现的术语列于下方
乙酰胆碱
附属行为
促肾上腺皮质激素
情感平衡疗法
情感意识(Affective Consciousness)
临床概念化
核心自我机制
定义
语言与
神经解剖学
神经生理学
研究挑战
情感功能
动物的
行为主义研究与理论
身体反应
临床意义
认知功能与
动物间的共同特征
意识与
当前理解
维度模型
情绪功能与
进化发展
体验
反馈理论
功能解剖学
语言习得中的
学习中的
控制层次
测量
记忆过程中的
神经生理学
精神分析理论
研究方法
感觉性情感
社会建构主义观点
心智理论中的
统一理论
动物研究的价值
意志控制
另见情感意识;情绪系统(总体);初级过程情绪(Primary-process Emotions);七个基本情感系统
情感神经科学(Affective Neuroscience)
脑心(BrainMind)概念化
意识研究
基本要素
演化模型
未来展望
目标
对发展心理学的影响
对心理治疗的影响
将研究发现整合到临床实践中
情绪系统模型
精神病理学模型
神经化学
整体-部分混淆
精神分析理论与
RAGE系统研究的理论依据
读出理论
研究挑战
理论发展
三角验证方法
《情感神经科学:人类和动物情绪的基础》(Panksepp)
情感神经科学人格量表
攻击行为
三级过程层面攻击冲动的相互作用
笑声与
神经生理学
游戏行为与
捕食性
RAGE系统与
性行为与
社会支配与
睾酮与
类型
向无生命物体发泄
Aichhorn, A.
5-α还原酶
AMPA
苯丙胺
杏仁核
情感处理功能
在恐惧条件反射中
在FEAR系统中
在情绪系统功能解剖中
在LUST系统中
在PLAY系统中
在RAGE系统中
无脑畸形
愤怒
在动物中
SEEKING系统的唤起
脑成像
临床概念化
个体差异
生理效应
愤怒带来的快感
情感神经科学研究的前景
RAGE与
另见 攻击行为;RAGE系统
快感缺失
动物
情感神经科学对动物的理解
动物的愤怒
动物认知过程研究
物种间共同的神经生理特征
条件性位置偏好/厌恶
区分积极和消极情绪
早期情感研究
早期关于动物心智和大脑的哲学概念化
动物的情绪体验
动物的情绪系统
情绪相关发声
情感体验的证据
情感的进化
实验性神经化学操控
群体攻击
动物的高阶情绪过程
稳态情感
无脊椎动物
动物间的母性养育
母婴结合
催产素研究
对动物的PANIC/GRIEF系统研究
阿片类输注的位置偏好
捕食行为
对动物的RAGE系统研究
繁殖策略
自我意识
自我刺激研究
感觉情感
动物的时间感
动物研究在情感研究中的价值
另见 大鼠和小鼠研究
预期
抗精神病药物
焦虑
情感来源
临床概念化
恐惧与焦虑
FEAR与GRIEF
先天的焦虑能力
催产素效应
药物治疗
血清素与
治疗
另见 FEAR系统
失语症
趋近动机系统
阿瑞匹坦
亚里士多德
芳香化酶
上行网状激活系统
评估
依恋过程
在人类中
依恋的损害或失败
不良依恋的代际传递
阿片假说
产后结合窗口期
理论发展
依恋的多样性
注意力缺陷多动障碍
圣奥古斯丁
Austin, J.
自闭症
自传体记忆
自知意识
自主神经系统
自动塑形
Averill, J.
基底神经节
基底外侧杏仁核复合体
终纹床核
行为激活系统
行为抑制系统
行为主义
行为矫正疗法
边沁
苯二氮卓类药物
柏林生物物理学俱乐部
Berridge, K.
β受体阻滞剂
β-内啡肽
生物胺
鸟类
威廉·布莱克
血压
边缘型人格障碍
Bowlby, J.
脑源性神经营养因子
脑心智(BrainMind)
情感神经科学概念化
行为主义概念化
早期神经科学研究
早期哲学概念化
内表型
表观遗传塑造
恐惧化学
嵌套层级模型
初级加工
研究挑战
性别发展
结构进化
动物研究在脑心智研究中的价值
Buck, R.
丁丙诺啡
Burghardt, G.
罗伯特·彭斯
丁螺环酮
癌症创伤治疗
大麻素
Cannon, W.
CARE系统
对后代的益处
跨物种表现
当前理解
发展目标
进化
功能
性别差异
对心理治疗的影响
与其他初级过程系统的相互作用
在母子互动中的表现
母婴结合
妊娠的神经化学
神经进化
神经生理学
促进母性感受
精神分析理论
心理学意义
SEEKING系统与
敏化
作为精神不适的来源
治疗性操控
另见 初级过程情绪;七种基本情感系统
儿茶酚胺
宣泄
猫
小脑皮层
变化
情感调节作为目标
氯氮卓
胆囊收缩素
乔姆斯基
染色质
扣带皮层
昼夜节律
Claparede, E.
Clarici, A.
经典条件反射
认知行为疗法
认知功能
情感功能与
在动物中
概念思维作为情感体验的要求
条件反射与
意识与
与皮层中线结构相关的缺陷
新皮层过度刺激中的妄想思维
在寻找因果关系中的妄想思维
进化
情感的反馈理论
意识层次
神经生理学
研究障碍
初级过程情绪与
在SEEKING系统中
自我相关信息处理
工作记忆
另见 三级过程
认知神经科学
认知理论和疗法
共病
心智计算理论
条件反射
作为自动脑反应
自动塑形与
经典模型
内源性阿片系统
消退
恐惧
通往条件反射的高级和低级通路
学习模型
神经生理学
条件反射中的疼痛
位置偏好/厌恶
对情境刺激的敏感性
另见 强化;非条件反应
信心
意识
情感功能与
在动物中,早期哲学概念化
基于运动功能系统的基础
认知和
核心
核心-自我的整合作用
作为无意识过程的学习
层次
神经进化
神经心理学模型
情绪唤醒的处理
初级过程情绪在研究中的相关性
研究目标
情境性恐惧条件反射
控制
情绪的
脑心的嵌套层级
愤怒系统调节
核心-自我
情感基础
解剖学
在动物中
临床概念化
当前理解
双面一元论模型
在知性意识的出现中
证据
进化
功能
自我相关处理中的下脑运动功能
情感意识的机制
感觉处理在
跨物种的变异
胼胝体
皮层中线结构
皮层-丘脑通路
促肾上腺皮质激素释放因子
情感反应
对玩耍系统的影响
在恐惧系统中
在悲伤系统中
长期暴露于
寻求系统唤醒
在分离痛苦中
皮层下回路
皮质醇
Cousins, N.
Cozolino, L.
好奇心
Damasio, A.
Darwin, Charles
Davanloo, H.
D-环丝氨酸(d-cycloserine)
陈述性记忆
深部脑刺激
抑郁症
适应功能
情感神经科学模型
脑刺激治疗
原因
皮质醇分泌和
强啡肽(dynorphin)和
悲伤系统在
神经生理学
神经科学研究
阿片假说
阿片刺激在治疗中
安慰剂效应在治疗中
产后
临床前建模
寻求系统在
压力和
治疗
Descarte, R.
《人类的由来》(Darwin)
个体发展
情感神经生理学
情感神经科学模型
压力暴露导致的脑损伤
条件反射效应
核心情感情绪
早期悲伤体验
治疗早期童年病理的有效性
情绪功能
表观遗传塑造
进化层次
经验决定因素
恐惧系统
婴儿对养育的需求
学习经验和
恐慌/悲伤系统的成熟
新皮质
神经模式
父母的理解
初级过程情绪
亲社会结构的
保护性情绪体验
精神分析理论
性
情绪体验的意义
玩耍的意义
社会功能
社会玩耍行为
在子宫内
《精神疾病诊断与统计手册》
诊断分类
地西泮(diazepam)
地西泮结合抑制剂
己烯雌酚(diethylstilbestrol)
弥散张量成像
双氢睾酮(dihydrotestosterone)
情绪生活的维度观
Dissanayake, E.
多巴胺能系统
预期和消费效应
在认知功能中
药物使用和
功能
在学习中
神经通路
神经受体
药物替代
在玩耍系统中
心理意义
奖励预测误差假说
在寻求系统中
自尊和
在情绪系统中的意义
在睡眠中
时间感和
背外侧额叶区
做梦
驱力理论
药物滥用和成瘾
相关的初级过程情绪
动物中的条件性位置偏好/厌恶
神经生理学
神经科学研究
过程
精神兴奋剂使用倾向于
寻求系统在
社会依恋和
二元论
强啡肽
Einstein, Albert
Ekman, P.
艾托拉嗪(eltoprazine)
情绪记忆
早期童年
恐惧学习和
内隐学习和
神经生理学
学习的初级过程控制和
心理意义
再巩固
治疗性重塑
无意识唤醒
情绪系统,一般而言
行动系统和
情感功能和
情感神经科学模型
在动物中
行为神经科学模型
发展
在独特自我的出现中
心理学理论的进化
反馈理论
更高认知处理
个体差异
作为习得反应
神经生理学
神经心理学观点
解读理论
另见 情感功能;情绪记忆
内表型
内啡肽
嫉妒
脑心的表观遗传塑造
癫痫
情景记忆
Epstein, M.
雌激素
动物行为学
进化
抑郁症的适应功能
情感功能
祖先记忆
基本情感系统
脑心结构
照料系统
意识
核心-自我发展
情绪系统
外适应(exaptation)
恐惧性
神经发育
恐慌/悲伤系统
玩耍行为
痛苦发声的作用
自我意识
感觉系统
性满足
睡眠和觉醒结构
核心情感的生存功能
外显记忆
《人类与动物的情绪表达》(Darwin)
条件反射的消退
眼动脱敏与再加工
Eysenck, H.
面部表情
家庭功能
顶核
恐惧
情感神经科学研究
条件反射
进化作用
体验
无畏
性别差异
天生能力
学习
神经化学
疼痛和
寻求感觉行为
主动暴露于恐惧情境
另见 恐惧系统
恐惧系统
适应功能
经典条件反射
认知神经科学模型
情境性条件反射
电刺激研究
在情绪学习和记忆中
情绪记忆在
恐惧条件反射和
功能
悲伤系统和
天生的神经能力
内在本质
学习和
在儿童中的表现
误解
神经化学研究
神经生理学
无对象的恐惧在
过度激活
疼痛和
恐慌/悲伤系统和
病理
基于药物治疗
精神分析理论和
愤怒系统和
研究需求
寻求系统和
焦虑的来源在
治疗意义
无条件反应在
另见 初级过程情绪;七个基本情感系统
情感反馈理论
Feld4蛋白家族
战斗/逃跑/僵住
闪光灯记忆
Fleming, A.
足部电击
宽恕
自由意志
Freud, S.
功能性磁共振成像
Gall, F. J.
γ-氨基丁酸
Gedo, J.
性别差异
攻击行为
愤怒反应
造成社会伤害的能力
研究中的概念问题
恐惧性
杀婴行为
PANIC/GRIEF系统的成熟
嫉妒的神经生理学
养育后代
催产素效应
游戏行为
初级过程情绪
SEEKING系统激活
性发育
性神经生理学
社会支配行为
遗传学
谷氨酸
在FEAR系统中
在GRIEF系统中
在记忆过程中
在RAGE系统中
受体
在SEEKING系统中
促性腺激素释放激素
Grandin, Temple
Gray, J.
Greenberg, L.
悲伤。另见 PANIC/GRIEF系统
Griffin, D.
内疚
习惯化
Harlow, H.
仇恨
听觉
Heath, R.
Hebb, D. O.
Hess, W.
海马体
皮质醇损伤
过度暴露于皮质醇的影响
在恐惧条件反射中
情绪系统的功能解剖
功能
记忆功能
神经可塑性
压力反应
θ节律
Hippocrates
Holt, R.
稳态情感
LUST作为情绪情感与
RAGE系统唤醒
稳态情感中的SEEKING系统敏化
同性恋
蜜蜂
Hubel, D.
Hume, David
体液理论
HUNGER(饥饿)
鬣狗
Hyman, S.
下丘脑
在CARE系统中
在皮质醇调节中
在FEAR系统中
情绪系统的功能解剖
在LUST系统中
在RAGE系统中
感觉处理功能
个性化自我
成像技术
丙咪嗪
内隐记忆
冲动控制障碍
杀婴
下丘
岛叶
胰岛素样生长因子1
情绪强度
内感受
中间核
精神分析释义
智商
Izard, C.
Jacob, F.
James, W.
James-Lange模型
Janov, A.
嫉妒
Kandel, E.
Kant, Immanuel
Kendrick, K.
Klein, D.
Kluver-Bucy综合征
Knutson, B.
Lange, C. 另见 James-Lange模型
语言
习得过程
情感意识与
情绪功能的行为神经科学观点
研究情绪的能力
情感描述
动物的情绪相关发声
意识进化与
语言的治愈力量
局限性
意义与
作为情感体验的必要条件
言语游戏
外侧膝状体核
外侧下丘脑
笑声
学习
情感神经科学模型
SEEKING系统中的食欲性学习
作为自动过程
行为主义理论
临床医生对学习的误解
条件反射
情境恐惧条件反射
情绪记忆与
情绪作为习得反应
恐惧性
游戏的功能
作为习得行为的母性奉献
在新皮层中
神经生理学
阿片假说
初级过程对情绪记忆的控制与
程序性学习
重复行为与
学习中的奖励和惩罚
奖励预测误差
奖励/强化理论
SEEKING系统与
工作记忆在学习中的作用
LeDoux, J.
Lichter, A.
边缘系统
Loeb, J.
London, Jack
长时程增强
弓背姿势
Lorenz, K.
爱
Ludwig, C.
LUST系统
CARE系统与
作为情绪-情感过程
作为物种保存的进化冲动
女性回路
功能
神经进化
催产素在其中的作用
心理意义
研究问题
SEEKING系统与
另见 初级过程情绪;七种基本情感系统;性与性行为
MacLean, P.
Malan, D.
Maren, S.
Marvell, Andrew
Mayberg, H.
McHugh, P.
Meaney, M.
意义性
内侧前脑束
动物自我刺激
连接性
在多巴胺能系统中
功能
一般行为功能
在SEEKING系统中
内侧膝状体核
褪黑素
记忆
情感成分
祖先记忆
作为自动过程
压力暴露造成的脑损伤
陈述性记忆
情绪学习与
眼动疗法
在自我意识生成中
心理健康与
关于记忆的误解
情绪体验期间的新皮层激活与
神经生理学
程序性学习
保持
SEEKING系统与
社会记忆
治疗目标
时间感与
记忆类型
情绪的无意识唤醒
工作记忆
另见 情绪记忆
中脑运动区
Miller, Anesa
Milner, P.
镜像神经元
一元论
单胺氧化酶抑制剂
运动功能
音乐
纳洛酮
纳曲酮
自恋
新皮层
去皮层研究
过度刺激中的妄想思维
发展
情绪系统的进化
发展的经验决定因素
在恐惧学习中
功能
在自我意识的生成中
大脑半球分工
在学习和记忆中
PLAY系统对新皮层的重塑
RAGE系统调节
在SEEKING系统中
工作记忆的基质
嵌套层次
神经经济学
神经肽。另见具体肽类
神经生理学
情感意识的
情感功能的
CARE系统
妊娠期的变化
认知系统与情绪系统
核心-自我解剖
抑郁症的
婴儿期养育的发展效应
早期研究
持续释放皮质醇的影响
情绪系统的
网络的表观遗传塑造
进化层次
情感意识的进化
实验性神经化学操作
FEAR系统
情绪强度
雄性间攻击
学习的
母性行为的
记忆过程的
母婴结合窗口期
依恋失败的结果
PANIC/GRIEF系统
阿片类药物注射的位置偏好
PLAY系统
RAGE系统
自我相关信息处理
感觉情感的
感觉处理的
性与性行为
社会丧失的
社会记忆的
压力反应
皮层下网络发展
情感研究的三角测量方法
工作记忆的
另见具体脑结构或系统
神经精神分析
神经科学,一般而言
认知神经科学
概念和技术演变
当前对情感功能的理解
另见 情感神经科学;成像技术
神经递质和受体
一氧化氮合酶
NMDA
去甲肾上腺素
伏隔核
尾侧脑桥网状核
Numan, M.
Olds, J.
阿片类药物
成瘾
愉悦效应
RAGE系统调节
动物皮层下研究
内源性阿片类物质
自闭症与
在CARE系统中
在抑郁症病因学中
通过刺激治疗抑郁症
外源性刺激和抑制
功能
在GRIEF系统中
在学习过程中
催产素与
在PLAY系统中
受体
SEEKING系统唤醒
触觉与
在社会功能中
治疗潜力
对立过程
眶额皮层
食欲素
O’Shea, K.
催产素
动物研究
自闭症与
在CARE系统中
当前研究
对PLAY系统的影响
内源性阿片类物质系统与
进化
功能
缝隙连接同步化
性别差异
GRIEF系统调节
在LUST系统中
音乐效应
外周效应
在妊娠期
主要效应
RAGE系统抑制
受体密度
在社会功能中
治疗潜力
疼痛、恐惧与
惊恐发作
动物研究
在边缘型人格障碍中
概念基础
在抑郁症中
发展目标
药物使用与
早期暴露
进化
FEAR系统与
功能
与其他初级过程系统的相互作用
表现形式
成熟
神经生理学
病理学
精神分析理论与
心理意义
SEEKING系统与
触摸的安抚作用
焦虑来源
治疗考虑
另见 初级过程情绪;七大基本情感系统
Panksepp, J.
副交感神经系统
室旁核
在CARE系统中
在皮质醇调节中
应激反应
Pasternak, Boris
Paulsen, S.
Peciña, S.
儿童心理治疗
导水管周围灰质
情感处理功能
在核心自我神经解剖学中
在FEAR系统中
情绪系统的功能解剖
在PANIC/GRIEF系统中
在RAGE系统中
在情绪系统中的意义
人格障碍
药物治疗
抗抑郁药物的作用
攻击行为治疗
焦虑治疗策略
抑郁症治疗
恐惧研究
针对FEAR系统相关障碍
一般性去抑制效应
FEAR系统的意义
操纵GRIEF系统
神经化学
催产素的潜在用途
长期使用精神兴奋剂
情感神经科学研究的前景
心理治疗与
RAGE系统抑制剂
RAGE系统研究的理论依据
在情绪记忆重塑中
基于血清素的
另见特定药物;特定类型药物
实践智慧
鸽子
垂体
安慰剂效应
柏拉图
游戏行为
攻击行为与
相关发声
临床益处
好奇心与
定义特征
学习
对冲动的调节因素
捕食练习
粗暴打闹形式
SEEKING系统与
性行为与
作为社交活动
产生的社会支配关系
邀请信号
另见 PLAY系统
动物间的共同特征
情境决定因素
发展意义
幼年动物的发展
做梦与
社会隔离的影响
负面情绪的出现
享受
在新皮层的表观遗传重塑中
进化起源
功能
多动症与
冲动控制障碍与
与其他初级过程系统的相互作用
误解
神经生理学
心理意义
SEEKING系统与
治疗考虑
图雷特综合征与
言语的
另见 游戏行为;七大基本情感系统
Plutarch
Plutchick, R.
正性情感和负性情感量表
实证主义
正电子发射断层扫描
后现代主义
创伤后应激障碍
增强
祈祷
捕食性攻击
妊娠
视前区
Press, O.
原始疗法
初级过程情绪
大脑的行动网络与
适应性功能
作为无意识的
评估
研究障碍
核心自我的基础
循环因果关系
当前理解
定义
在抑郁症中
在双面一元论模型中
体验
功能
性别心智
高级心智互动
语言与
LUST作为
神经振荡
情绪系统的神经进化
神经生理学
作为前语言的
心理意义
情绪系统的结构
皮层下基础
在治疗关系中
治疗意义
动物研究的价值
另见 七大基本情感系统
灵长类梳理行为
过程体验疗法
孕酮
催乳素
普萘洛尔
原始自我
精神分析理论
精神病理学
情感神经科学模型
诊断概念化
依恋失败导致的
FEAR系统的
GRIEF系统的
情绪系统间的相互作用
代际传递
LUST系统的
临床前建模
情感神经科学方法的前景
RAGE系统的
SEEKING系统的
动物研究的价值
另见 心理治疗;特定诊断
精神病
心身疾病
精神兴奋剂使用
心理治疗
情感平衡疗法
情感调节作为目标
儿童治疗
处理童年早期经历的有效性
对情绪过程的新兴兴趣
整合情感神经科学发现
动物研究的局限性
GRIEF系统考虑
PLAY系统考虑
情感神经科学方法的前景
精神药物治疗与
情绪记忆的再巩固
FEAR系统的意义
初级过程情绪的意义
对RAGE系统的理解
积极社会情绪的价值
情绪记忆的工作
情感体验
动物研究
唤醒
脑成像研究
临床概念化
FEAR系统与
性别差异
抑制剂
神经进化
神经生理学
对象
研究障碍
相关病理
生理效应
捕食性攻击行为与
情感神经科学研究的理论基础
调节
SEEKING系统与
假怒与
社会支配行为与
三级过程表现
治疗考虑
战争与
另见 初级过程情绪;七种基本情感系统
Raimondi, Marantonio
大鼠和小鼠研究
厌恶性条件反射
情绪相关的发声
杀婴行为
先天恐惧
LUST系统研究
母性行为
神经解剖学
游戏行为
捕食性攻击
SEEKING行为
分离痛苦
性发育
情绪的读出理论
神经递质受体
抗抑郁药物的作用
激动剂
拮抗剂
阻断剂
化学操纵
婴儿期养育的发育效应
在记忆过程中
阿片受体
催产素受体
性激素
强化
宗教与灵性
快速眼动睡眠
重复行为
网状结构
复仇
奖励预测误差
奖励/惩罚
动物自我刺激研究
愉悦的预期
行为主义理论
心理学理论的概念演变
动物的条件性位置偏好/厌恶
恐惧条件反射
稳态模型
学习理论
神经生理学
神经科学研究
强化程序
感觉奖励
仪式化行为
Rogers, C.
Rolls, E.
Romanes, G.
Roosevelt, F. D.
Russell, J.
Sacks, O.
悲伤。见 PANIC/GRIEF
施虐受虐狂
Saunders, James
精神分裂症
Schlaepfer, T.
Schore, A.
Schultz, W.
Scott, J. P.
次级过程
攻击行为与
自塑与
唤醒的行为表现
CARE系统与
概念演变
持续运作
过度刺激中的妄想思维
药物使用与
眼动疗法与
信仰与
功能连接
功能
性别差异
稳态失衡效应
对抑郁症治疗的意义
杀婴行为与
固有唤醒刺激
与其他情感系统的互动
学习功能
LUST系统与
测量
记忆与
误解
神经进化
神经生理学
过度消费与
病理
游戏行为与
PLAY系统与
预期的愉悦
捕食行为与
精神分析理论与
心理意义
RAGE系统与
重复附加行为与
自我相关处理
感觉寻求行为
动物间的相似性
策略性和创造性思维
时间感与
活动不足
另见 初级过程情绪;七种基本情感系统
选择性5-羟色胺再摄取抑制剂
自我概念
作为嵌入式
作为具身
哲学和心理学演变
另见 核心自我
《自我意识的脑》 (Damasio)
自尊
感觉寻求
感觉系统
情感特征
情感功能的行为神经科学观点
意识与
核心自我过程
发育
恐惧预测线索处理
LUST作为情绪情感与感觉情感
运动系统联系
神经进化
神经生理学
在PLAY冲动中
在性行为中
在社会联结中
皮层下处理
分离痛苦
GRIEF系统
神经解剖学
神经化学
催产素效应
发声
大脑隔区
5-羟色胺
特征
当前理解
发育
在神经网络的表观遗传塑造中
进化功能
性别差异
对认知的影响
先天神经能力
相互作用
学习与
调节
对心理治疗的意义
动物间的相似性
在动物研究中的价值
另见特定系统
性与性行为
攻击与
与动物发声相关
交配权竞争
研究中的概念问题
文化和政治背景
当前理解
发育
手指长度生物标志物
一般社会行为与
同性恋和跨性别者
杀婴行为与
性高潮
游戏行为与
捕食性攻击
精神分析理论
第二性征
升华理论
另见 LUST系统
Shapiro, F.
兄弟姐妹竞争
Siegel, A.
Siegel, D.
Skinner, B. F.
慢波睡眠
嗅觉
社会建构主义
社会关系
成瘾特征
基本情感系统
临床意义
哀悼中的文化差异
当前神经科学理解
PLAY系统的发育意义
支配行为
心理学理论的演变
母婴联结
神经生理学
阿片假说
游戏行为
心理意义
RAGE系统唤醒
拒绝
SEEKING系统功能
性行为与
技能发展
社会记忆
交流冲动
在工作场所
Solomon, A.
躯体标记假说
灵魂
Spitz, R.
Spurzheim, J. G.
惊吓反应
《哭泣骆驼的故事》
皮层下脑
情感功能
临床意义
在认知功能中
动物间的共同特征
发育
早期研究
进化
情绪系统的功能解剖
在产生自我意识中
作为原始过程情绪的位置
LUST系统
快速条件反射
感觉处理
皮层下中线系统
升华
P物质
上丘
视交叉上核
交感神经系统
味觉
教师
气质
时间感
在动物中
记忆与
SEEKING系统与
三级过程
作为愤怒
在动物中
临床意义
意识
当前情绪过程理论
定义
攻击冲动的互动
睾酮
《生物精神病学教科书》
丘脑
心智理论
治疗关系
θ节律
THIRST(口渴系统)
桑代克, E.
挠痒
廷伯根, N.
蒂普顿, 比利
汤姆金斯, S.
触觉
妥瑞症
移情
跨性别者
创伤治疗
图尔文, E.
酪氨酸
非条件反应
昂格施泰特, U.
功利主义
阴道-宫颈刺激
瓦伦斯坦, E.
加压素
血管加压素
维尔曼斯, 马克斯
腹侧被盖区
腹内侧下丘脑
视觉系统
活力论
田鼠
冯·布吕克, E.
战争
华生, J. B.
维塞尔, T.
威廉斯, G. W.
威利斯, T.
怀斯, R.
维特根斯坦, L.
工作记忆
工作场所
ζ抑制肽
版权所有 © 2012 by Jaak Panksepp and Lucy Biven
保留所有权利
第一版
如需复制本书选段的许可信息,请致信:Permissions, W. W. Norton & Company, Inc., 500 Fifth Avenue, New York, NY 10110
如需大量采购特别折扣信息,请联系 W. W. Norton Special Sales:specialsales@wwnorton.com 或 800-233-4830
书籍设计:MidAtlantic Publishing Services
制作经理:Leeann Graham
美国国会图书馆出版物编目数据
Panksepp, Jaak, 1943-
The archaeology of mind : neuroevolutionary origins of human emotions / Jaak Panksepp, Lucy Biven. – 1st ed.
cm. -- (A Norton professional book) (The Norton series on interpersonal neurobiology)包含参考文献和索引。
ISBN 978-0-393-70531-7 (精装)
BF531.P36 2012
152.4–dc23
2012010323s
ISBN: 978-0-393-70531-7
EISBN: 978-0-393-70731-1
W. W. Norton & Company, Inc., 500 Fifth Avenue, New York, N.Y. 10110
www.wwnorton.com
W. W. Norton & Company Ltd., Castle House, 75/76 Wells Street, London W1T 3QT
1 2 3 4 5 6 7 8 9 0
[1]我们必须记住,情绪区域的皮层下神经元的放电速度通常比丘脑和新皮层的高级感知区域慢得多。在许多系统中,神经放电的聚集模式比其更高的模式化速率更重要。通常,某些情感-内脏神经化学物质(如神经肽)的力量和持续作用在情绪感受和反应的模式中比神经放电速率的升高更重要,而PET和fMRI等技术监测的是由此产生的血流增加。因此,这些技术对低级情感脑功能的敏感度不如对高级认知功能的敏感度,这在使用这些技术研究情绪时产生了不良偏差:它们对情绪的伴随认知-思维相关性比对情感基质更敏感。
[2]当前的脑成像时代,尽管基于更准确的脑活动测量(血流和代谢变化),仍然产生的脑功能图像并不能准确反映产生心理事件的潜在脑活动。它们只是让我们更准确地估计哪些脑区可能最重要,需要研究以了解心智-大脑的组织方式。正如大多数从业者所知,这种新的”颅相学”仍然面临许多挑战,在试图将脑活动转化为心理过程时存在麻烦的缺陷。
[1]目前人们对那些对预测奖励的刺激特别感兴趣和互动的动物越来越关注,即那些”信号追踪者”和那些似乎对即将到来的食物最感兴趣的”目标追踪者”。前者在对预期刺激的反应中表现出比奖励本身更多的脑多巴胺唤起,而后者对预测性刺激和目标刺激都表现出更适度的唤起。这似乎反映了潜在SEEKING系统的气质特征。信号追踪者比目标追踪者更容易对可卡因等药物成瘾(Flagel等人,2011)。
[2]“燃素”(Phlogiston)是早期物理学家给一种假想的(理论上假设的)物质起的名字,可燃材料含有这种物质使它们能够燃烧。当它们燃烧成灰烬后,这些物质被认为是”去燃素化”的。当然,这被证明是一个实际上并不存在的实体的名称;它被用来产生一种理解的感觉,在有任何真正理解之前。
[1]对抗过程(opponent process)是一种倾向于直接抑制在它之前并触发它的过程的效果。例如,某些成瘾药物的积极情感在大脑内部被负面情感感受的积累所抵消,这导致戒断的痛苦。
[1]苯二氮卓类药物也称为小镇静剂或抗焦虑药,最早的包括氯氮卓和地西泮(品牌名称为利眠宁和安定)。现在有许多其他种类的小镇静剂,其中一些也作为安眠药或肌肉松弛剂出售。
[1]http://www.emory.edu/INTELNET/fi.hasid.html
[1]例如,我们长期以来一直困惑于这样一个事实:对下丘脑的恐惧系统进行极低水平的电刺激可以促使动物僵住,而增加电流则可以引发逃跑行为(Panksepp, Sacks et al., 1991)。如此截然不同的恐惧反应怎么可能共享一个共同的神经基础?解决这一难题的一种理论方法是假设,在较高电流水平下,我们的局部脑刺激正在扩散到寻求系统,这使动物从静止的僵住状态中解脱出来,进入逃跑的显著步伐,因为它们在”寻求安全”——显然这对动物来说应该是一种更愉快的状态。它应该充满希望!这提出了一个引人入胜的理论可能性,即恐惧的惊恐如何可能爆发为乐观的渴望,有时几乎是逃跑的嬉戏性。不幸的是,这一想法仍有待严格的神经科学评估,因此不应将其视为结论,而只是作为进一步研究的工作假设。
[1]本章和下一章完全由Jaak Panksepp撰写,旨在分享一个愿景(以及一些个人回忆),关于哺乳动物情绪的知识如何能够帮助推进生物精神病学科学的发展,以及发展可能颇具争议的新心理治疗方法。
[2]应该指出的是,关于哺乳动物无条件恐惧系统细节的大量精彩工作已经从巴西的几个实验室中出现,最突出的是与Frederico Graeff(例如,Del-Ben & Graeff, 2009)和Marcus Brandão(Brandão et al., 2005)合作的研究人员。
[1]计算机革命推动了这样一种观念,即深层生物性的心智可以在硅平台上进行计算——这一愿景在新的认知科学中占主导地位,并且似乎在学术界的许多其他角落仍然活跃(参见Panksepp, 2008c)。